 2024, Vol. 44
2024, Vol. 44文章信息
- 孙宇璇, 曾涛, 陈双丹, 张珮
- SUN Yuxuan, ZENG Tao, CHEN Shuangdan, ZHANG Pei
- 彭州市不同土地利用方式土壤小型分解者群落结构和功能
- The community structure and function of soil micro-decomposers under different land use types in Pengzhou City
- 生态学报. 2024, 44(13): 5567-5582
- Acta Ecologica Sinica. 2024, 44(13): 5567-5582
- http://dx.doi.org/10.20103/j.stxb.202307271611
-
文章历史
- 收稿日期: 2023-07-27
- 网络出版日期: 2024-04-25





2. 四川大学生命科学学院 四川省濒危野生动物保护生物学重点实验室, 成都 610065
2. Sichuan Provincial Key Laboratory of Conservation Biology of Endangered Wild Animals, College of Life Sciences, Sichuan University, Chengdu 610065, China
分解作用是农业生态系统关键的养分循环过程之一, 也是土壤肥力的重要来源, 强烈影响着农作物生长与农业生态系统的生产力[1—2]。土壤微生物、线虫等土壤小型分解者作为分解作用的重要参与者, 它们的生命活动和相互作用不仅在分解生物残体、改变土壤理化性质以及促进物质循环和能量转化过程中起着重要作用, 还驱动着地上植物的多样性和生产力[3]。大量研究评估并探讨了不同自然生态系统中分解作用的速率和影响因素[4], 但在受到强烈人为干扰和土地利用方式频繁发生变化的农业生态系统中分解作用却鲜少受到关注[5—7]。如何在维持生产力的同时, 尽量降低成本, 合理配置土地资源, 维持并提高土壤肥力和生产力, 一直是农业可持续发展的关键议题[8]。因此, 评估农业生态系统中不同生境分解作用的效率和影响因素, 优化土地利用方式和人为管理模式, 对于农业生态系统结构和功能的维持至关重要。
分解作用的效率主要取决于三个方面, 分别是气候条件、凋落物质量和数量以及分解者群落特征[9]。在较大地理尺度下, 气候条件是影响凋落物的主要因素, 但在较小尺度上, 凋落物质量和数量以及分解者群落特征对分解作用的影响更大[9]。总体而言, 单位面积凋落物质量越低、难分解成分越多, 分解越难进行[10]。不同生境下的土壤温度、湿度等性质直接影响了凋落物分解的微环境, 同时这些土壤物理环境还影响了土壤动物的生命活动, 如水分限制会对土壤动物取食造成不利影响, 进而减慢分解速率[8, 11]。土壤动物是影响凋落物分解的重要生物因子, 通过破碎、掘穴、取食和刺激微生物活动等直接或者间接影响凋落物分解[12]。土壤生物群落组成和数量的变化势必也会导致其生态功能的改变, 进而影响分解速率[7]。土壤线虫取食细菌和真菌, 通过改变微生物群落结构来影响分解过程[7, 12]。线虫和微生物对环境变化敏感, 是土壤质量的良好指示生物[13]。同时, 二者均在土壤养分循环中发挥着重要作用。如细菌是土壤中分布最广、数量最多的微生物类群, 在调节土壤C、N循环等生物化学过程中起着至关重要的作用[14];真菌可以通过与植物共生影响植物生长于生产力, 也具备改善土壤肥力的功能[15]。农田生态系统的土地利用方式多样, 不同生境下的土壤物理和化学特性具有显著差异, 土壤生物数量和群落结构特征也不同[2]。这些不同生境下分解者群落的差异如何影响了农田生态系统的分解作用仍不清楚。本实验拟在气候条件相似的农田生态系统内, 运用两种标准化茶包模拟凋落物, 进而揭示不同土地利用方式下土壤分解者群落特征和分解作用之间的关系。
自1999年四川省退耕还林工程实施以来, 成都作为工程实施重点市之一, 有计划地停耕并改造了一些作物产量低的耕地, 因地制宜地造林种草, 恢复植被, 使成都市的土地利用方式发生了很大转变[4]。彭州市位于成都市东北方向, 区内农业用地面积占比大, 耕地面积达到75.7%, 是四川省内具有代表性的斑块型农业区域。彭州市的农业用地方式多样, 在面积较小区域范围内同时存在农耕地、灌草丛、湿地、人工林等多种生境类型, 因此可以作为开展不同生境土壤生物群落和分解作用效率研究的良好试验区域。已有研究报道过该区在城市绿地、农田、撂荒地、湿地和林地下土壤性质及土壤动物群落的差异[4], 研究结果发现不同土地利用方式下土壤动物群落组成及结构差异较大, 但总体而言地表植被种类及数量的增加和土壤扰动的减少有助于土壤动物生存。但是, 从土壤分解者群落的生态功能角度出发综合评价该区在不同生境下分解速率的研究尚未有报道。本研究拟通过野外调查分析并比较彭州市几种主要生境土壤分解者群落特征及分解速率, 探讨农业生态系统中分解者群落和分解速率对土地利用方式变化的响应。该研究旨在为彭州市农业生态系统的肥力维持、土地规划利用和农业管理措施提供理论依据。
1 材料与方法 1.1 研究区域概况彭州市隶属四川成都市, 地理坐标为东经103°40′—104°10′,北纬30°54′—31°26′, 该区域为亚热带湿润季风气候, 年平均气温为15.6 ℃, 年平均降水量为867 mm。研究区域主要为丘陵地形, 样地最高海拔659 m, 最低547 m。
1.2 实验样方设置及样品采集样方设置:2022年5月在彭州市布设试验样地, 选择6处同时存在农田、经济林、次生杂木林和撂荒地四种生境类型的样地, 样地大小为100 m× 100 m(图 1)。每个样地里, 相邻样地之间至少间隔5 km;同一样地内, 每种生境类型的面积至少为200 m2, 相邻不同生境采样点至少间隔10 m。

|
| 图 1 研究区域示意图 Fig. 1 Schematic map of the study area |
当地农田为玉米(Zea mays L.)和油菜(Brassica napus L.)轮作。春季4月种植玉米, 种植规格为大行1.3 m, 小行0.7 m, 株距0.2 m。种植前将磷肥或钾肥与优质农家肥混合均匀作为底肥, 施入耕作层土壤。秋季9月前后收获玉米, 10月种植冬油菜, 于次年4月收获。
经济林为种植桂花树(Osmanthus fragrans)或银杏(Ginkgo biloba)的纯林, 两种经济林均种植5 a以上, 实行常规林地管理, 林下主要为空心莲子草(Alternanthera philoxeroides), 地面有较厚的凋落物层。
次生杂木林以银杏、枇杷(Eriobotrya japonica)和核桃树(Juglans regia L.)为主, 混生有皂角树(Gleditsia sinensis)、喜树(Camptotheca acuminata)、构树(Broussonetia papyrifera)和马尾松(Pinus massoniana)等, 栽种约8—10 a, 期间无人为管理。地面草本层为空心莲子草、酢浆草(Oxalis corniculata L.)等, 基本无凋落物,
撂荒地为休耕2—5 a后形成的以草本和低矮灌木为主的灌草丛, 地面凋落物极少。主要植物种类有接骨草(Sambucus chinensis)、香椿(Toona sinensis)和猪秧秧(Galium spurium L.)等。
土壤样品采集:本研究选择土壤线虫、土壤细菌和真菌作为小型分解者群落的代表。于2022年8月11日进行土壤取样。在每种生境类型下, 以棋盘取样法分别采集15个有代表性的原位土样, 用直径5 cm的裂式土钻收集地表下0—10 cm原状土样, 并将同一样地内相同生境下的土样混合均匀, 手动分拣肉眼可见的石块和动植物残体。混合均匀的新鲜土样分为四部分, 其中一部分新鲜土样用于分离与鉴定土壤线虫, 一部分存放于-20 ℃冰箱中用于提取土壤微生物DNA, 一部分存放于4 ℃冰箱中用于测定土壤铵态氮和硝态氮含量, 其余土样自然风干后用于土壤基本理化性质的测定。
分解作用测定:本研究引入茶包指数(Tea Bag Index, TBI)作为标准化定量描述不同生境分解作用效率的指标[10]。由于自然条件下不同生境中凋落物的种类和质量差异较大, 为了更直观地比较不同生境下分解速率的快慢及其与各自生境中分解者群落和环境因子的关系, 本研究使用两种不同类型的茶包(立顿绿茶和红茶茶包, 联合利华公司, 净含量2 g)作为标准化凋落物, 分别代表快速(绿茶)和慢速(红茶)分解基质。将两种茶包在不同生境类型的土壤中掩埋一段时间后分别测定其质量损失, 以评价分解者群落和环境因子对有机质分解速率造成的影响。在红茶和绿茶茶包标签上标记茶叶类型, 于2022年8月11日成对掩埋在各样地中, 每种生境类型掩埋3对(4种生境类型× 6个重复样地× 3对茶包/样地=共72对)。掩埋时, 将茶包放入地表下8 cm深, 保持标签在土壤上方可见, 并标记掩埋地点。掩埋90天后将茶包挖出取回, 称取剩余茶叶重量, 并计算一定时间内被分解的茶叶重量。
1.3 研究方法 1.3.1 土壤理化性质的测定土壤含水量和容重采用烘干法测定;土壤pH的测定采用电位法并用酸度计(赛多利斯PB-10 pH)测量[16]。土壤总氮和总碳使用元素分析仪(FlashSMART, 美国)测定;土壤铵态氮和硝态氮先用1 mol/L的氯化浸提后, 采用流动分析仪(SEAL Auto Analyzer3, 德国)测定[16]。土壤速效钾的测定采取火焰光度法, NH4OAc(1 mol/L)浸提后用火焰光度计(FP6410)测定;有效磷采取双酸浸提-钼锑抗比色法测定。
1.3.2 土壤线虫分离、计数和鉴定称取100 g新鲜土壤样品, 加入500 ml浓度为4%的甲醛溶液, 杀死并固定土壤线虫。采用LUDOX TM原液悬浮法分离土壤线虫[17]。分离后的土壤线虫在解剖镜(OLYMPUS, SZX7)下计数, 并换算为每100 g干土中的线虫数量。每个样品随机挑取120条左右线虫制备成永久玻片, 在2000X显微镜下(OLYMPUS, BX51)鉴定到属水平。鉴定资料参考《中国土壤动物检索图鉴》。根据Yeates[18]将鉴定到的土壤线虫分为食细菌线虫(Bacterivores, 简称Ba)、食真菌线虫(Fungivores, 简称Fu)、植食性线虫(Herbivores, 简称H)、捕食和杂食性线虫(Omnivores-predators, 简称O/P)以及食藻类线虫(Algivores, 简称A)。并将所有的线虫样品赋予对应的c-p值, 以计算相关的生态学指数[19]。
参考Bonger[20]的方法, 本研究计算了如下四类线虫生态学指数:
(1) 多样性指数
计算了衡量线虫属丰富度的Margalef丰富度指数(Margalef richness Index, SR), 显示线虫优势属的存在与优势程度的优势度指数(Dominance index, λ), 以及衡量群落中土壤线虫在属水平上均匀程度的均匀度指数(Evenness index, J′)。
(2) 成熟度指数
植食线虫成熟度指数(Plant parasite index, PPI)反应了植食性线虫对植物的危害程度[21]。其计算公式为:
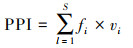
|
自由生活线虫的成熟度指数(Maturity index, MI)能够反映线虫群落演替阶段, 进而评价土壤受干扰的程度。低MI表明线虫群落演替处于早期阶段, 土壤受干扰程度高; 高MI说明线虫群落演替处于后期, 受到的干扰较少[21]。其计算公式为
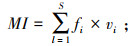
|
式中, vi为i属线虫的c-p值;fi为样本中i属出现的频率。
PPI与MI这两个指数的比值能够反映土壤养分富集水平和受扰动程度。PPI/MI值越高, 说明土壤养分富集水平越高, 生境受干扰水平也越高[21]。
(3) 线虫通道指数(Nematode channel ratio, NCR)用来反映土壤有机质的分解途径, 其值在1和0之间, 值越大说明分解过程更倾向由细菌完成, 反之则说明分解更依赖真菌[18]。其计算公式为
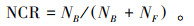
|
Wlaszka指数(Wasilewska index, WI)用于反映土壤线虫群落组成和土壤的健康程度, WI值越大, 表明土壤矿化途径主要由食细菌线虫和食真菌线虫参与, 土壤健康程度越高;其值越低, 表明矿化途径主要由植食性线虫参与, 土壤健康程度越低[22]。计算公式为
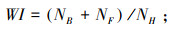
|
式中, NB为样本中食细菌线虫的数量;NF为样本中食真菌线虫的数量;NH为样本中植食性线虫数量。
(4) 富集指数(Enrichment index, EI)反映了外界营养物质输入的水平, 较高的EI代表更多的外部营养输入, 土壤食物网有更多可利用的外部资源[15]。其计算公式为
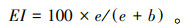
|
结构指数(Structure index, SI)表示土壤食物网的连通性和食物链的长度。SI越高, 食物网相对连通性越高, 食物链越长[15]。其计算公式为
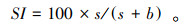
|
式中, b(Basal)是食物网基础组分, 代表c-p值为2的食细菌和食真菌线虫类群;e(Enrichment)是食物网富集组分, 代表c-p值为1食细菌线虫和c-p值为2的食真菌线虫;s(Structure)是食物网结构组分, 代表c-p值为3—5的食细菌、食真菌、捕食/杂食线虫类群。
1.3.3 土壤微生物分析本实验采用DNeasy PowerSoil DNA试剂盒(Takara公司)提取土壤总DNA。称取0.25 g新鲜土样, 严格按照试剂盒说明书提取土壤总DNA, 并用1%琼脂糖凝胶电泳进行检测, 核酸分析仪(NanoDrop 2000)测定其浓度。
将DNA样品统一稀释到20 μg/μL后, 对细菌16S rRNA V3—V4区和真菌ITS rRNA 1区进行PCR扩增[23]。扩增产物使用1%琼脂糖凝胶电泳鉴定和分离条带, DNA胶纯化试剂盒(Invitrogen 75510-019)对PCR产物进行纯化。纯化后的产物等量混合后在Illumina Miseq平台进行高通量测序, 原始数据以FASTQ格式保存。
将原始数据进行聚类处理, 采用以OTUs聚类为基础的Vsearch方法分析土壤细菌群落的原始数据。选用Silva数据库(Release132, http://www.arb-silva.de)对细菌16S rRNA基因进行物种分类学注释[24], UNITE数据库(Release 8.0, https://unite.ut.ee/)对真菌ITS序列进行物种分类学注释[25]。用QIIME2(2019.4)软件中的classify-sklearn算法[26]对每个细菌和真菌OTU代表序列使用预先训练好的Naive Bayes分类器进行物种注释。
利用QIIME2(2019.4)软件中的qiime feature-table rarefy功能对OTU表做95%深度样本序列量的抽平处理, 以获取每个样本内微生物在不同生境下各分类水平的具体组成表。未经抽平的细菌OTU表使用FAPROTAX(Functional Annotation of Prokaryotic Taxa)工具进行功能注释[27];真菌则使用FUNGuild工具依据ITS序列预测其潜在功能[28]。
为了测定土壤细菌和真菌样品的绝对丰度, 本研究通过SYBR Green Ⅰ染料法对细菌16S rRNA和真菌ITS1基因进行实时荧光绝对定量检测(Quantitative Real-time PCR, qPCR)。将标准品按10倍浓度梯度稀释, 然后以其为模板进行PCR反应, 标准样品和供试样品均设置3次重复。以标准品拷贝数的对数为横坐标, 以所得的Ct值(指实时监测扩增过程的荧光信号达到指数扩增时的循环周期数)为纵坐标, 绘制标准曲线。对样品在相同条件下进行qPCR反应获取样品的Ct值, 并根据标准曲线计算出拷贝数[29]。所得Eub338基因、ITS1基因拷贝分别表征土壤微生物群落中活细菌和真菌的总多度。
1.3.4 数据分析茶包指数(TBI)由分解速率(Decomposition rate index, k)和凋落物稳定因子(Litter stabilization factor, S)共同组成, 其中k表征茶包在一段时间内分解速率的快慢, S表示凋落物中不易水解的部分在生境中的稳定性, S与k的计算公式如下[10]:
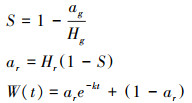
|
式中, ag为绿茶实际被分解重量的比例;Hg为绿茶茶包可水解部分重量的比例, 为常数84%[10];ar为红茶实际被分解重量的比例;Hr为红茶茶包可水解部分重量的比例, 为55%[10];W为剩余红茶茶叶重量;t为茶包掩埋至回收的天数。
本研究采用单因素方差分析(ANOVA)和最小显著差异法(Fisher′s LSD test)比较土壤理化性质、植物群落特征、线虫生态指数、线虫营养类群组成、土壤微生物群落组成、分类群和功能类群丰度在不同生境之间的差异, 方差分析在SPSS 24.0进行。采用主坐标分析(Principal coordinates analysis, PCoA)比较各生境间细菌和真菌群落组成的相似性。使用Mantel test[11]分析土壤细菌、真菌和线虫与土壤理化因子、植物群落特征和分解作用之间的相关性, 各变量之间的相关性用Pearson相关性系数表示, 上述分析在R 4.2.3运用Qiime2包进行。
2 研究结果 2.1 不同生境类型下的环境因子不同生境下土壤理化性质和植被盖度如表 1所示。土壤硝态氮含量在农田中最高, 杂木林其次, 撂荒地和经济林中最低。杂木林的土壤总碳、总氮、速效钾含量和乔木盖度都显著高于其它三种生境(P<0.05), 速效钾在杂木林中的含量达到其它生境的1.6倍以上。四种生境下的土壤都呈弱酸性, 撂荒地和杂木林的土壤pH值显著高于农田和经济林(P=0.04)。土壤容重在撂荒地中最高, 农田中最低, 农田中的土壤容重比撂荒地低了32%。杂木林和经济林中乔木盖度显著高于撂荒地(P=0.04), 农田中则没有乔木存在。农田中的草本盖度最高, 显著高于其它生境(P=0.02)。杂木林植被种类最多, 撂荒地次之, 农田和人工林植物丰富度最低。四种生境间土壤含水率、铵态氮和有效磷含量无显著差异。
| 环境因子 Environmental factor |
农田 Farmland |
撂荒地 Fallow land |
经济林 Economic woodland |
杂木林 Mixed woodland |
| 土壤含水率Soil moisture content/% | 23.39±1.67a | 22.42±2.35a | 24.23±2.81a | 25.32±1.52a |
| 土壤容重Soil bulk density/(g/cm3) | 1.19±0.03c | 1.77±0.01a | 1.54±0.03b | 1.47±0.02b |
| 土壤pH Soil pH | 5.68±0.08c | 6.75±0.09a | 5.90±0.13b | 6.40±0.07a |
| 土壤总氮Soil total nitrogen/(g/kg) | 2.29±0.03b | 2.04±0.13c | 1.99±0.06c | 3.57±0.08a |
| 土壤总碳Soil total carbon/(g/kg) | 27.33±0.66b | 26.01±2.08b | 23.94±1.03b | 34.42±1.44a |
| 土壤硝态氮Soil nitrate nitrogen/(mg/kg) | 8.94±0.42a | 2.89±0.56c | 3.15±0.48c | 5.17±0.89b |
| 土壤铵态氮Soil ammonium nitrogen/(mg/kg) | 32.67±4.42a | 32.78±4.63a | 38.76±9.99a | 40.67±7.80a |
| 土壤有效磷Soil available phosphorus/(mg/kg) | 147.44±15.90a | 122.44±15.41a | 159.75±13.14a | 138.50±30.89a |
| 土壤速效钾Soil available potassium/(mg/kg) | 75.00±7.98bc | 80.67±5.02b | 55.17±6.81c | 135.50±7.94a |
| 乔木平均盖度Average arbor coverage/% | 0.00c | 1.83±0.91c | 67.50±3.81b | 83.33±2.79a |
| 灌木平均盖度Average shrub cover/% | 0.00b | 22.00±7.03a | 0.00b | 11.67±3.61ab |
| 草本平均盖度Average herb coverage/% | 64.17±5.39a | 34.17±5.69b | 26.67±4.41b | 30.83±5.23b |
| 植物丰富度Plant richness | 4.33±0.61c | 6.50±0.42b | 3.33±0.83c | 9.33±0.81a |
| 不同小写字母表示不同生境下的环境因子在P<0.05水平上差异显著 | ||||
微生物拷贝数表示绝对丰度, 代表了不同生境间微生物不同分类群数量的差异。单因素方差分析结果显示, 土壤真菌丰度在不同生境间存在显著差异, 经济林和杂木林真菌丰度显著高于撂荒地和农田(P =0.01)(图 2)。不同生境下的细菌丰度则无显著差异(图 2)。

|
| 图 2 不同生境类型下土壤细菌和真菌的绝对丰度 Fig. 2 Total absolute abundances of soil soil bacteria and fungi in different habitat types 不同小写字母表示不同生境下的土壤微生物基因拷贝数在P<0.05水平上差异显著 |
相对丰度表征微生物群落组成在不同生境类型间的组成差异。所有生境类型中, 细菌的优势类群(门水平)均是厚壁菌门(Firmicutes)和变形菌门(Proteobacteria;图 3)。在各生境类型中厚壁菌门占比分别为:农田21%, 经济林12%, 次生杂木林11%, 撂荒地10%;且农田中厚壁菌门占比显著高于其它生境(P=0.03)。担子菌门(Basidiomycota)在除农田外的其它三类生境中都是优势类群, 占比均超过17%, 但在农田中的占比仅5%(图 3)。

|
| 图 3 不同生境类型下土壤细菌和真菌主要类群的相对丰度 Fig. 3 Relative abundances of the main taxa in soil bacterial and fungal community in different habitat types |
四种生境间变形菌门相对丰度的差异不显著, 子囊菌门(Ascomycota)在各生境真菌群落中的占比都超过60%(图 3), 但其相对丰度在各生境间无显著差异。相比细菌群落, 真菌群落组成在生境间差异更大(图 3)。
2.2.2 土壤微生物主要分类群和功能群丰度不同生境中土壤细菌群落主要分类群的绝对丰度较为相似, 仅厚壁菌门表现出显著差异(图 4)。农田和经济林中厚壁菌门绝对丰度均显著高于撂荒地(P=0.03), 尤其是农田中厚壁菌门的丰度是撂荒地中的2倍。除壶菌门外, 其余土壤真菌主要类群的绝对丰度在四种生境间的差异则较大(图 5)。其中, 子囊菌门、担子菌门和被孢菌门的绝对丰度均在经济林中最高(图 5), 杂木林中罗兹菌门和球囊菌门的丰度则显著最高(图 5)。

|
| 图 4 不同生境类型下土壤细菌主要类群的绝对丰度 Fig. 4 Abundances of the main taxa in the bacterial community in different habitat types 不同小写字母表示不同生境下的土壤细菌基因拷贝数在P<0.05水平上差异显著 |

|
| 图 5 不同生境类型下土壤真菌主要类群的绝对丰度 Fig. 5 Abundances of the main taxa in the fungal community in different habitat types 不同小写字母表示不同生境下的土壤真菌基因拷贝数在P<0.05水平上差异显著 |
主坐标分析(PCoA)的结果显示, 撂荒地和杂木林中的土壤细菌群落结构相似性较高(图 6)。土壤真菌群落的结构在各生境间差异极显著, 且比细菌群落呈现出更高的生境差异性(图 6)。农田中的土壤细菌和真菌群落结构均与其它三种生境有较大差异(图 6)

|
| 图 6 不同生境下土壤细菌和真菌群落的PCoA分析 Fig. 6 The principal coordinates analysis (PCoA) of soil soil bacterial and fungal community in different habitat types |
根据FAPROTAX功能预测的结果, 本研究特别关注了与C循环相关、N循环相关和病原细菌三类细菌。研究结果显示, 农田中病原细菌丰度最低, 且显著低于撂荒地和杂木林(P=0.02)(图 7)。各功能群中参与C循环的细菌类群丰度最高, 在各生境间差异并不显著(图 7)。FUNGuild功能预测则将土壤真菌划分为病原真菌、共生真菌和腐生真菌三类, 且以腐生真菌占比最高(图 7)。并且, 杂木林和撂荒地中腐生真菌的丰度显著高于农田和经济林(P=0.01), 病原真菌的丰度也表现出和腐生真菌相同的趋势(图 7)。共生真菌在杂木林和撂荒地中的丰度也显著高于农田(P= 0.01)(图 7)。

|
| 图 7 不同生境下土壤细菌和真菌功能类群的绝对丰度 Fig. 7 Absolute abundances of soil bacterial and fungal functional taxa in different habitat types 不同小写字母表示不同生境下的土壤微生物功能类群基因拷贝数在P<0.05水平上差异显著 |
本次研究调查到的土壤线虫分属6目16科25属(附表 3)。其中经济林和杂木林的线虫丰度显著高于农田和撂荒地中的线虫丰度(表 2)。不同线虫营养类群的比较结果显示, 撂荒地中的食细菌线虫丰度相对最低, 每100 g干土中较其它三类生境少60条以上;杂木林中的食真菌和捕食/杂食类线虫数量最多, 显著高于其它三类生境(P=0.01);捕食/杂食类线虫在农田和撂荒地中数量极少, 每100 g干土中数量小于7条, 经济林和杂木林中这两类线虫数量较高, 每100 g干土中超过30条(表 2)。食藻类线虫仅在杂木林土壤中发现且丰度过低(附表 3), 因此未在表 2中列出。
| 农田 Farmland |
撂荒地 Fallow land |
经济林 Economic woodland |
杂木林 Mixed woodland |
|
| 线虫丰度Soil nematode abundance/(individual/100 g dry soil) | ||||
| 总丰度Total abundance | 275±31.36b | 259±44.67c | 455±84.81a | 513±73.71a |
| 植食性线虫Herbivores | 90.41±4.17a | 110.65±8.34a | 184.19±18.09a | 157.52±15.63a |
| 食细菌线虫Bacterivores | 164.55±9.15ab | 104.03±8.81b | 196.38±11.27a | 171.83±11.42ab |
| 食真菌线虫Fungivores | 15.56±1.36b | 37.59±3.71b | 43.72±6.00b | 110.82±8.72a |
| 捕食/杂食线虫Omnivores/predators | 3.80±0.74c | 6.77±1.40c | 30.28±2.73b | 47.21±3.06a |
| 生态学指数Ecological index | ||||
| 丰富度指数Margalef richness Index, SR | 2.13±0.01b | 2.33±0.01b | 2.50±0.01b | 3.51±0.01a |
| 优势度指数Dominance index, λ | 0.18±0.01a | 0.12±0.01bc | 0.13±0.01b | 0.09±0.01c |
| 均匀度指数Evenness index, J′ | 0.81±0.02a | 0.75±0.03a | 0.84±0.02a | 0.80±0.04a |
| 植食性线虫与自由生活线虫成熟度指数比Plant parasitism index/Maturity index | ||||
| PPI/MI | 1.88±0.30a | 1.09±0.13b | 1.86±0.24a | 1.39±0.20ab |
| 线虫通道指数Nematode channel index, NCR | 0.90±0.02a | 0.72±0.06bc | 0.83±0.04ab | 0.63±0.07c |
| Wlaszka指数Wlaszka index, WI | 2.02±0.16a | 1.44±0.25a | 1.51±0.25a | 2.11±0.33a |
| 富集指数Enrichment index, EI | 80.69±4.12a | 53.95±0.81c | 62.65±6.01b | 53.86±1.26c |
| 结构指数Structure index, SI | 23.80±7.26b | 18.62±4.08b | 23.09±6.35b | 35.75±1.52a |
| 不同小写字母表示不同生境下的线虫丰度和生态指数在P<0.05水平上差异显著 | ||||
由表 2可知, Margalef丰富度指数、优势度指数、PPI/MI指数、富集指数和结构指数在不同生境间差异显著(P<0.05)。次生杂木林中土壤线虫群落的Margalef丰富度指数和结构指数SI在四种生境中最高。农田的优势度指数最高, 是杂木林中的2倍(P=0.03)。PPI/MI指数(P=0.02)和富集指数EI (P=0.04)均在农田中最高, 经济林次之, 且均显著高于撂荒地。四种生境下的线虫通道比NCR都大于0.5, Wlaszka指数WI均大于1, 不同生境间差异不显著。
2.4 不同生境类型下的分解作用对比不同生境类型下的TBI指数发现, 杂木林和撂荒地中分解速率指数k显著高于经济林和农田(图 8), 杂木林的分解速率指数(k=8.41×10-3)几乎是农田(k=4.59 ×10-3)的2倍。凋落物稳定因子S也在各生境间存在显著差异, 从高到低依次为农田>经济林>撂荒地>杂木林(图 8)。

|
| 图 8 不同生境类型下的茶包指数(TBI指数) Fig. 8 Tea Bag Indices in different habitat types 不同小写字母表示不同生境下TBI指数在P<0.05水平上差异显著 |
Mantel test结果表明, 土壤线虫和微生物群落都与土壤理化性质和植被盖度存在一定显著相关关系。土壤总碳含量与三类分解者群落均显著相关(P<0.05), 但土壤线虫与全碳含量的相关系数(r=0.18)低于土壤细菌群落(r=0.23)和土壤真菌群落(r=0.26)。细菌群落还与土壤容重(r=0.15, P=0.02)、速效钾含量(r=0.17, P=0.02)和有效磷含量(r=0.21, P=0.01)显著相关。线虫群落则与土壤全氮(r=0.25, P=0.01)和速效钾(r=0.18, P=0.02)显著正相关, 和乔木盖度间也存在极显著相关关系(r=0.29, P<0.001)。
分解作用与多个环境因子密切相关。表征分解作用的两项指标S和k为极显著负相关。S与除铵态氮、硝态氮和有效磷外的所有土壤理化因子都存在显著或极显著的负相关关系, 且与乔木盖度极显著负相关, 草本盖度极显著正相关(P<0.001), 但与分解者群落相关性较弱。k与容重、pH和灌木盖度极显著正相关, 与硝态氮含量极显著负相关(P<0.001), 同时与土壤细菌群落存在显著相关性(r=0.21, P=0.03)。
3 讨论本研究发现彭州农田生态系统四类典型生境(农田、撂荒地、经济林和杂木林)的土壤细菌、真菌和土壤线虫的群落组成、结构和多样性均有显著差异。这些差异可能与不同土地利用方式中的植被群落特征、生境复杂度和人为管理方式相关[30—31]。不同生境下分解效率的差异与分解者群落的差异相适应。杂木林和撂荒地中分解效率最高, 经济林次之, 农田最低。
研究结果表明, 林业生态系统中土壤微生物的丰度更高, 经济林和杂木林中的细菌和真菌丰度均高于农田和撂荒地(图 2)。植物是土壤小型分解者的重要资源来源, 其凋落物经过分解, 能够作为土壤微生物的稳定碳源[9], 相比于灌木和草本植物, 高大的木本植物能够产生更多可供分解的凋落物, 因此林地中土壤微生物的丰度更高, 分解作用速率也相对更快[32](图 2)。同时, 相比农田作物, 林木具有更加发达的植物根系, 有利于和植物共生的球囊菌门的聚集(图 5)。此外, 研究结果还显示与土壤养分富集和物质循环密切相关的子囊菌门、担子菌门和被孢菌门在林地中的绝对丰度都比较高(图 5), 说明经济林和杂木林更适合这类微生物的生存和聚集, 进而提高了林地中养分循环的效率[33—34]。厚壁菌门能通过促进土壤中的有机物降解, 增加土壤中的有益微生物数量, 改善土壤结构, 提高土壤肥力[35]。结果显示在杂木林中, 厚壁菌门丰度显著高于撂荒地(图 4), 说明林木刺激了厚壁菌门的繁殖和生存, 进而对土壤肥力增长做出一定贡献。土壤线虫群落也与林地生态系统密切相关(图 9)。相比农田和撂荒地, 经济林与杂木林中植物群落的丰富度更高, 有利于更多土壤微生物和土壤动物的聚集, 为线虫提供了更多食物资源, 因此总线虫丰度、植食性线虫丰度和捕食/杂食线虫丰度在林地中有所提高(表 2)。同时, 比较两种林地生境, 木本植物盖度更高, 种类更丰富的次生杂木林中, 食真菌线虫和捕食/杂食线虫丰度也显著更高[33](表 2)。各生境中土壤线虫富集指数的对比结果也进一步证明了林地能够为土壤提供更多的资源输入。尽管杂木林和撂荒地均不存在显著的人为干扰, 植物丰富度均较高, 但是次生杂木林比撂荒地拥有更多的可利用资源, 因而能够支持一个更加庞大的地下食物网[35]。同时, 杂木林中地下分解者群落之间的联系也更紧密, 因此土壤线虫的结构指数也高于农田和撂荒地, 食物网成熟度更高(表 2)。

|
| 图 9 环境因子、土壤小型分解者和分解作用之间的相互关系 Fig. 9 The correlations between environmental factors, soil micro-decomposers and decomposition SMC:土壤含水率Soil moisture content; BD:容重Soil bulk density; pH:土壤pH Soil pH; TN:土壤总氮Soil total nitrogen; TC:土壤总碳Soil total carbon; NO3:土壤硝态氮Soil nitrate nitrogen; NH4:土壤铵态氮Soil ammonium nitrogen; AP:土壤有效磷Soil available phosphorus; AK:土壤速效钾Soil available potassium; AC:乔木盖度Arbor coverage; SC:灌木盖度Shrub coverage; HC:草本盖度Herb coverage; PR:植物群落丰富度Plant richness; S:凋落物稳定指数Litter stability index; k:分解速率Decomposition rate |
此外, 地上植被丰富度也会影响分解者群落的多样性和碎屑食物网[1, 9—10]。Wang等[36]在比较单一作物农田与植物种类更丰富的自然土壤中发现, 自然土壤中杂食性和捕食性线虫比例更高, 群落间连通性更强。这表明更复杂的地上环境能够提升地下食物网的复杂度, 食物网自上而下的调节能力也更强[6]。本研究中, 杂木林的地上植物丰富度最高(表 1), 生境中不仅存在能够增加垂直异质性的多种木本植物, 林间还生长有灌木和草本。撂荒地中以灌木和多年生草本为主, 植被丰富度仅次于次生林, 且显著高于种植单一经济树种的人工林或种植单一作物的农田(表 1)。因此认为杂木林植被类型最丰富, 撂荒地次之, 经济林和农田则相对简单。杂木林中线虫群落的多个生态指数都优于其它生境, 如杂木林中土壤线虫群落的多样性最高, 反映土壤小型食物网复杂程度的结构指数也更高, 说明杂木林能够支持一个更加复杂且多样化的土壤小型食物网[27]。同时, 线虫通道比NCR在杂木林中最低, 表明参与分解的食真菌线虫比例较其他生境更多, 可以分解更多难分解的底物, 分解作用能够更彻底[8, 32]。而生境复杂程度相对最低的农田中土壤线虫群落丰富度最低, 优势度和NCR最高, 线虫群落结构趋于单一化, 多以体型小、c-p值低的线虫参与分解。生境内植被类型复杂程度还会同时影响分解作用速率k和凋落物稳定因子S(图 8)。本研究发现随着地上植物丰富度的增加, k值增大而S值降低。这与Becker等[37]对比10种不同复杂程度生境间TBI的结果一致:随着生境异质性的增强, 分解速率增加, 同时环境对不稳定组分降解的抑制作用降低。对比发现杂木林和撂荒地中k值都显著较高, 且与分解速率显著相关的土壤细菌群落结构也存在很高相似性(图 6)。推测这两类相对更复杂的生境中可能存在大量参与分解作用的细菌, 如硝化细菌属、反硝化细菌属、芽孢杆菌属和假单胞菌属等[36]。
农业管理措施等人为干扰是农田生态系统的重要特征, 不同土地利用方式下人为干扰强度的差异可能也是影响农田生态系统分解者群落组成、多样性和功能的重要因素[37]。在本研究选取的四类生境中, 农田和经济林均受到不同强度的人为管理, 如中耕除草、施肥追肥、灌溉排水和防治病虫等, 而杂木林和撂荒地则几乎不存在人为干扰。因而, 土壤线虫PPI/MI的结果也反映出相同趋势, 农田受干扰程度最高, 经济林次之, 杂木林和撂荒地都较少受到外在干扰(表 2)。同时, 受干扰程度较高的农田和经济林中, 共生真菌和腐生真菌的丰度低于杂木林和撂荒地(图 7)。共生真菌具有改善植物营养、调节植物代谢和增强植物抗病能力等功能, 腐生真菌更是直接参与分解作用, 这两类微生物的数量对于农田生态系统土壤肥力维持和植物养分吸收具有重要意义[35]。而农田中强烈的人为干扰可能在一定程度上抑制了这些土壤中有益微生物的聚集[5—7, 38], 阻碍了共生菌和腐生菌的活动。
值得注意的是, 人为干扰对于土壤分解者群落组成、多样性和功能的影响并不完全是不利的。人为管理控制了土壤中病原菌的丰度, 病原细菌和病原真菌的丰度在农田和经济林中都低于撂荒地和杂木林(图 7), 这可能与人工施用农药相关。合理、适量地施用农药可以控制有害微生物的数量, 维持土壤健康, 进而提高经济作物成活率, 对保障生产力, 提高农林业经济效应有重要意义。对比杂木林, 存在少量人为干扰的经济林中子囊菌门、担子菌门和被孢菌门的丰度优势更加明显(图 5)。这三类微生物与土壤养分循环关系密切, 说明适度人为干扰能够促进土壤的营养元素循环。本研究也发现存在人为管理措施的经济林中植食性线虫和食细菌线虫的数量更高(表 2), 这与Li等[39]对6825个全球土壤样品数据中线虫营养类群对土地管理强度响应的研究结果相同。这两类线虫在食物网中处于较低营养级, 食性简单, 获取食物资源更容易, 且由于c-p值小, 生存周期短, 生长发育速度快, 因此能够更好地适应环境变化[39]。
除了植物群落特征外, 土壤pH值、土壤容重和土壤总碳含量是影响本研究中土壤分解者群落结构的重要因素, 且pH值和容重还与分解作用速率显著相关(图 9)。这些土壤理化性质在不同生境中的差异可能与人为施肥和松土密切相关。农田中施加的氮肥会经由硝化作用转化成硝酸盐, 雨水和灌溉会使部分硝酸盐流失至植物根系活动层以下, 这个过程会带走土壤中大量的钙、镁离子。且硝化作用产生大量氢离子, 因此施肥会导致pH值下降, 土壤酸化[40];而松土则会降低土壤容重。土壤pH值则与土壤微生物的生物量及群落结构相关[40], pH值可以通过改变土壤微生物的组成与分布模式进而对线虫群落产生间接影响[40]。施肥造成的土壤酸度增加和翻犁土层造成的容重降低, 改变了地下分解者群落的生活环境, 使分解者的生命活动和数量都受到抑制[4], 并最终显著影响了分解速率(图 9)。因此, 存在人为管理的农田和经济林中, 分解速率显著低于杂木林和撂荒地这两种半自然生境。此外, 有机肥料的施入、轮作和深翻等有利于土壤含碳量的提高[41]。本研究也发现土壤含碳量最高的杂木林中土壤线虫丰度最高, 捕食/杂食类线虫丰度也显著最高。Pan等[42]在东北黑土区开展的研究同样发现, 线虫总丰度和捕食/杂食类线虫丰度在土壤含碳量更高的区域更高, 这是因为含碳量高的土壤提供了适合微生物生存繁殖的环境, 增加微生物生物量, 给土壤线虫提供了更多的食物资源[43—44]。
4 结论综上所述, 本研究通过比较研究彭州市四种不同生境内环境因子、土壤分解者群落特征以及分解作用的差异, 发现提高生境中的乔木盖度, 能够有效增加小型分解者数量、改善土壤肥力以及增强土壤食物网的稳定性。生境异质性的提高促进了分解作用, 分解效率的提升又增加了单位质量土壤中养分元素的含量, 进而可能提高植物养分利用的效率。人为干扰改变了土壤酸度和紧实程度, 不利于土壤微生物参与分解, 降低了分解速率, 但对减少病原微生物, 维持土壤健康和促进元素循环也有一定正向效果。因此, 推行农林间作模式的混合型农业, 在增加乔木盖度、提高垂直生境异质性和维持土壤食物网连通性的同时, 适度进行施肥和病虫害管理, 以维持养分含量和健康状况良好的土壤环境, 促进分解作用更快速、彻底地进行, 这对提高农田生态系统的土壤肥力、维持产量具有重要意义。
致谢: 廖静和耿雪薇对样地选择和野外采样方法提供指导, 谢映虹对数据分析和图表制作提供帮助, 特此致谢!| [1] |
Handa I T, Aerts R, Berendse F, Berg M P, Bruder A, Butenschoen O, Chauvet E, Gessner M O, Jabiol J, Makkonen M, McKie B G, Malmqvist B, Peeters E T H M, Scheu S, Schmid B, van Ruijven J, Vos V C A, Hättenschwiler S. Consequences of biodiversity loss for litter decomposition across biomes. Nature, 2014, 509: 218-221. DOI:10.1038/nature13247 |
| [2] |
Bradford M A, Wieder W R, Bonan G B, Fierer N, Raymond P A, Crowther T W. Managing uncertainty in soil carbon feedbacks to climate change. Nature Climate Change, 2016, 6: 751-758. DOI:10.1038/nclimate3071 |
| [3] |
Wang Y, Wei W, Yang X Z, Chen L D, Yang L. Interrelationships between soil fauna and soil environmental factors in China: research advance. Ying Yong Sheng Tai Xue Bao=the Journal of Applied Ecology, 2010, 21(9): 2441-2448. |
| [4] |
杨丽红, 石红艳, 游章强, 唐中海, 刘昊, 贾小东. 不同土地利用方式对大型土壤动物群落结构的影响. 四川农业大学学报, 2015, 33(2): 208-214. |
| [5] |
Xue R, Wang C, Zhao L, Sun B R, Wang B L. Agricultural intensification weakens the soil health index and stability of microbial networks. Agriculture, Ecosystems & Environment, 2022, 339: 108118. |
| [6] |
Sánchez-Moreno S, Ferris H, Young-Mathews A, Culman S W, Jackson L E. Abundance, diversity and connectance of soil food web channels along environmental gradients in an agricultural landscape. Soil Biology and Biochemistry, 2011, 43(12): 2374-2383. DOI:10.1016/j.soilbio.2011.07.016 |
| [7] |
Beare M H, Reddy M V, Tian G, Srivastava S C. Agricultural intensification, soil biodiversity and agroecosystem function in the tropics: the role of decomposer biota. Applied Soil Ecology, 1997, 6(1): 87-108. DOI:10.1016/S0929-1393(96)00150-3 |
| [8] |
Guo T F, Zhang Q, Ai C, Liang G Q, He P, Zhou W. Nitrogen enrichment regulates straw decomposition and its associated microbial community in a double-rice cropping system. Scientific Reports, 2018, 8: 1847. DOI:10.1038/s41598-018-20293-5 |
| [9] |
García-Palacios P, Maestre F T, Kattge J, Wall D H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecology Letters, 2013, 16(8): 1045-1053. DOI:10.1111/ele.12137 |
| [10] |
Keuskamp J A, Dingemans B J J, Lehtinen T, Sarneel J M, Hefting M M. Tea Bag Index: a novel approach to collect uniform decomposition data across ecosystems. Methods in Ecology and Evolution, 2013, 4(11): 1070-1075. DOI:10.1111/2041-210X.12097 |
| [11] |
Duan X Z, Sun J T, Wang L T, Shu X H, Guo Y, Keiichiro M, Zhu Y X, Bing X L, Hoffmann A A, Hong X Y. Recent infection by Wolbachia alters microbial communities in wild Laodelphax striatellus populations. Microbiome, 2020, 8(1): 104. DOI:10.1186/s40168-020-00878-x |
| [12] |
Griffiths B S. Microbial-feeding nematodes and protozoa in soil: their effectson microbial activity and nitrogen mineralization in decomposition hotspots and the rhizosphere. Plant and Soil, 1994, 164(1): 25-33. DOI:10.1007/BF00010107 |
| [13] |
López-Fando C, Bello A. Variability in soil nematode populations due to tillage and crop rotation in semi-arid Mediterranean agrosystems. Soil and Tillage Research, 1995, 36(1/2): 59-72. |
| [14] |
Yang F, Wu J J, Zhang D D, Chen Q, Zhang Q, Cheng X L. Soil bacterial community composition and diversity in relation to edaphic properties and plant traits in grasslands of Southern China. Applied Soil Ecology, 2018, 128: 43-53. DOI:10.1016/j.apsoil.2018.04.001 |
| [15] |
Ferris H, Bongers T, de Goede R G M. A framework for soil food web diagnostics: extension of the nematode faunal analysis concept. Applied Soil Ecology, 2001, 18(1): 13-29. DOI:10.1016/S0929-1393(01)00152-4 |
| [16] |
于向华, 张明. 氯化钾浸提法测定土壤中铵态氮含量条件研究. 农业科技与装备, 2013(3): 11-12. |
| [17] |
Palmer L, Neilson R, Griffiths B S, Boag B. The use of colloidal silica to extract nematodes from small samples of soil or sediment. Nematologica, 1990, 36(1/2/3/4): 465-473. |
| [18] |
Yeates G W. Nematodes as soil indicators: functional and biodiversity aspects. Biology and Fertility of Soils, 2003, 37(4): 199-210. DOI:10.1007/s00374-003-0586-5 |
| [19] |
Bongers T. The maturity index: an ecological measure of environmental disturbance based on nematode species composition. Oecologia, 1990, 83(1): 14-19. DOI:10.1007/BF00324627 |
| [20] |
Bongers T, Bongers M. Functional diversity of nematodes. Applied Soil Ecology, 1998, 10(3): 239-251. DOI:10.1016/S0929-1393(98)00123-1 |
| [21] |
陈云峰, 韩雪梅, 李钰飞, 胡诚. 线虫区系分析指示土壤食物网结构和功能研究进展. 生态学报, 2014, 34(5): 1072-1084. DOI:10.5846/stxb201307021821 |
| [22] |
Liu T, Ye C L, Chen X Y, Ran W, Shen Q R, Hu F, Li H X. Effects of different organic manure sources and their combinations with chemical fertilization on soil nematode community structure in a paddy field of East China. Ying Yong Sheng Tai Xue Bao=the Journal of Applied Ecology, 2013, 24(12): 3508-3516. |
| [23] |
柳习月, 朱琪, 杨帆, 张娟, 张巧玲, 李江华, 王莉. 多组学解析酱香型大曲风味物质的形成. 食品与发酵工业, 2021, 47(22): 35-41. |
| [24] |
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner F O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research, 2013, 41(D1): D590-D596. |
| [25] |
Kõljalg U, Nilsson R H, Abarenkov K, Tedersoo L, Taylor A F S, Bahram M, Bates S T, Bruns T D, Bengtsson-Palme J, Callaghan T M, Douglas B, Drenkhan T, Eberhardt U, Dueñas M, Grebenc T, Griffith G W, Hartmann M, Kirk P M, Kohout P, Larsson E, Lindahl B D, Lücking R, Martín M P, Matheny P B, Nguyen N H, Niskanen T, Oja J, Peay K G, Peintner U, Peterson M, Põldmaa K, Saag L, Saar I, Schüßler A, Scott J A, Senés C, Smith M E, Suija A, Taylor D L, Telleria M T, Weiss M, Larsson K H. Towards a unified paradigm for sequence-based identification of fungi. Molecular Ecology, 2013, 22(21): 5271-5277. DOI:10.1111/mec.12481 |
| [26] |
Bokulich N A, Subramanian S, Faith J J, Gevers D, Gordon J I, Knight R, Mills D A, Caporaso J G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nature Methods, 2013, 10: 57-59. DOI:10.1038/nmeth.2276 |
| [27] |
Louca S, Parfrey L W, Doebeli M. Decoupling function and taxonomy in the global ocean microbiome. Science, 2016, 353(6305): 1272-1277. DOI:10.1126/science.aaf4507 |
| [28] |
Nguyen N H, Song Z W, Bates S T, Branco S, Tedersoo L, Menke J, Schilling J S, Kennedy P G. FUNGuild: an open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecology, 2016, 20: 241-248. DOI:10.1016/j.funeco.2015.06.006 |
| [29] |
高宇琼. 金花茶体胚发生过程中PPO基因的克隆及其表达[D]. 福州: 福建农林大学, 2009.
|
| [30] |
袁访, 邓承佳, 唐静, 宋理洪. 不同土地利用方式下土壤动物对凋落物的分解作用及影响因素. 土壤学报, 2023, 60(2): 568-576. |
| [31] |
张曼夏, 季猛, 李伟, 刘华存, 王彦杰, 张林, 潘开文. 土地利用方式对土壤团聚体稳定性及其结合有机碳的影响. 应用与环境生物学报, 2013, 19(4): 598-604. |
| [32] |
Li Y N, Qian Z Y, Li D J. Effects of tree diversity on soil microbial community in a subtropical forest in Southwest China. European Journal of Soil Biology, 2023, 116: 103490. DOI:10.1016/j.ejsobi.2023.103490 |
| [33] |
Römbke J, Höfer H, Garcia M V B, Martius C. Feeding activities of soil organisms at four different forest sites in Central Amazonia using the bait lamina method. Journal of Tropical Ecology, 2006, 22(3): 313-320. DOI:10.1017/S0266467406003166 |
| [34] |
Zhu L Y, Chen Y, Sun R B, Zhang J B, Hale L, Dumack K, Geisen S, Deng Y, Duan Y H, Zhu B, Li Y, Liu W Z, Wang X Y, Griffiths B S, Bonkowski M, Zhou J Z, Sun B. Resource-dependent biodiversity and potential multi-trophic interactions determine belowground functional trait stability. Microbiome, 2023, 11(1): 95. DOI:10.1186/s40168-023-01539-5 |
| [35] |
张雅茜, 方晰, 冼应男, 王振鹏, 项文化. 亚热带区4种林地土壤微生物生物量碳氮磷及酶活性特征. 生态学报, 2019, 39(14): 5326-5338. DOI:10.5846/stxb201809081925 |
| [36] |
Wang K H, Kokalis-Burelle N, McSorley R, Gallaher R. Cover crops and organic mulches for nematode, weed and plant health management. Nematology, 2008, 10(2): 231-242. DOI:10.1163/156854108783476412 |
| [37] |
Becker J N, Kuzyakov Y. Teatime on Mount Kilimanjaro: assessing climate and land-use effects on litter decomposition and stabilization using the Tea Bag Index. Land Degradation & Development, 2018, 29(8): 2321-2329. |
| [38] |
王移, 卫伟, 杨兴中, 陈利顶, 杨磊. 我国土壤动物与土壤环境要素相互关系研究进展. 应用生态学报, 2010, 21(9): 2441-2448. |
| [39] |
Li X P, Liu T, Li H X, Geisen S, Hu F, Liu M Q. Management effects on soil nematode abundance differ among functional groups and land-use types at a global scale. The Journal of Animal Ecology, 2022, 91(9): 1770-1780. |
| [40] |
Lucas-Borja M E, Candel D, Jindo K, Moreno J L, Andrés M, Bastida F. Soil microbial community structure and activity in monospecific and mixed forest stands, under Mediterranean humid conditions. Plant and Soil, 2012, 354(1): 359-370. |
| [41] |
Quist C W, Gort G, Mooijman P, Brus D J, van den Elsen S, Kostenko O, Vervoort M, Bakker J, van der Putten W H, Helder J. Spatial distribution of soil nematodes relates to soil organic matter and life strategy. Soil Biology and Biochemistry, 2019, 136: 107542. |
| [42] |
周程爱, 张于光, 肖烨, 张小全, 李迪强. 土地利用变化对川西米亚罗林土壤活性碳库的影响. 生态学报, 2009, 29(8): 4542-4547. |
| [43] |
Pan F J, Xu Y L, McLaughlin N B, Xue A G, Yu Q, Han X Z, Liu W, Zhan L L, Zhao D, Li C J. Response of soil nematode community structure and diversity to long-term land use in the black soil region in China. Ecological Research, 2012, 27(4): 701-714. |
| [44] |
Su L X, Bai T Y, Qin X W, Yu H, Wu G, Zhao Q Y, Tan L H. Organic manure induced soil food web of microbes and nematodes drive soil organic matter under jackfruit planting. Applied Soil Ecology, 2021, 166: 103994. |