 2024, Vol. 44
2024, Vol. 44文章信息
- 杨岚, 寇旭阳, 付晓, 郑栓宁, 吴钢, 陆兆华, 桑卫国
- YANG Lan, KOU Xuyang, FU Xiao, ZHENG Shuanning, WU Gang, LU Zhaohua, SANG Weiguo
- 长白山温带森林生态系统关键要素耦合及演替过程动态模拟
- Coupling key elements and simulating the succession dynamic in the Changbai Mountain temperate forest ecosystem
- 生态学报. 2024, 44(12): 5377-5388
- Acta Ecologica Sinica. 2024, 44(12): 5377-5388
- http://dx.doi.org/10.20103/j.stxb.202401050044
-
文章历史
- 收稿日期: 2024-01-05
- 网络出版日期: 2024-04-19





2. 中国科学院生态环境研究中心, 北京 100085;
3. 中国科学院城市环境研究所, 厦门 361021;
4. 中国矿业大学(北京), 北京 100083
2. Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
3. Institute of Urban Environment, Chinese Academy of Sciences, Xiamen 361021, China;
4. China University of Mining & Technology, Beijing 100083, China
面对生态修复和恢复中缺乏系统性、连续性、持续性等问题, 我国正在开展的“山水林田湖草沙一体化修复和保护”工程, 采用统筹、综合、整体的方法进行[1—2]。但是由于生态系统内和系统间耦合机制不清、理论基础缺乏等限制一体化保护和修复的推进, 迫切需要开展生态系统耦合理论、方法和技术的研究, 这对我国山水项目的实施和促进一体化修复和保护实践具有非常重要的意义[3]。目前在生态系统尺度上, 生态要素间耦合机制不清楚, 生态要素间的关系认识不深[4], 开展生态系统尺度上生态要素耦合研究显得尤其重要。
长白山温带森林生态系统是我国北方生态屏障带的重要组成部分[5—6]。探究森林生态系统动态过程要素的耦合机制, 以便揭示演替过程中组分与结构的变化, 对于了解生态系统功能至关重要[7]。森林生态系统的演替过程较复杂, 受到环境要素:太阳辐射、降水、风速、温度、土壤类型、土壤有机质含量, 和生物要素:树种本身的生理特性、树种与树种的相互作用以及土壤微生物的活动影响, 所有这些关键要素在演替的长时间尺度上也会发生变化[8]。可见, 森林生态系统的演替动态过程复杂, 所涉及的要素较多, 将森林生态系统演替过程中的多关键要素耦合, 体现“生命共同体”理念, 模拟森林生态系统的演替过程, 对于认识生态系统的生命一体化具有重要的科学意义。
在生态系统空间尺度, 以及百年以上的时间尺度上, 动态地考虑多关键要素的耦合较为复杂。研究从斑块的概念出发将关键生态系统因素进行耦合, 以一定时间段为步长, 探究在“水土气生”等多关键要素的耦合影响下, 斑块内单个树种更新、生长、死亡等主要过程的动态变化, 从而实现对整个森林生态系统百年以上的动态模拟[9—10]。LINKAGES系列是综合考虑生态关键要素最多林窗模型之一, 与其他林窗模型相比, 最大特征是充分考虑地下的生物地球循环过程, 充分体现了“生命共同体”的理念[11]。LINKAGES 3.0是最新的版本, 纠正了以往出现的问题, 并更新了过时的水文等模块, 从整体、综合、统筹的角度出发, 充分考虑了林分动态、碳氮循环、和水文过程之间的相互作用[12], 将森林生态系统的多要素进行耦合, 揭示了森林演替过程中树种组成和结构随时间的变化, 并量化了林分主要特征(蓄积量、生物量等指标)的变化过程, 使森林的动态过程得到了定量的预测[13]。
本研究旨在从生命共同体的理念出发, 以“统筹、综合、整体”角度聚焦于生态系统尺度, 运用LINKAGES 3.0模型探究森林生态系统动态演替过程中关键要素“水土气生”的耦合机制, 并模拟长白山森林生态系统的动态演替过程, 用系统性、交互性、动态性的模拟实例, 为我国“山水林田湖草”生态修复工程提供理论支撑以及科学工具。本研究将围绕以下两个科学问题进行开展:(1)LINKAGES 3.0模型中各子模型间是如何耦合的?森林生态系统关键生态要素间的耦合机制?(2)长白山温带针阔混交林树种组成与结构以及总生物量在演替过程中的变化?
1 研究地区与研究方法 1.1 研究区研究区位于我国东北部的长白山自然保护区的北坡(42°23′ N, 128°5′ E), 平均海拔801.5 m, 最高海拔809.5 m, 最低海拔791.8 m, 主要建群种为红松(Pinus koraiensis)、紫椴(Tilia amurensis)、蒙古栎(Fraxinus manshurica)、水曲柳(Quercus mongolica)、色木槭(Acer mono)等[14—15]。夏季短暂且温暖, 冬季漫长且寒冷, 年平均气温仅在3 ℃左右。年降水量约为700—800 m。太阳辐射为124—125 kcal cm-2 a-1。年平均风速小于3.9 m/s[16]。该区的地带性土壤为暗棕壤, 透水性好、氧化作用强, 表层腐殖质含量高, 具有高的土壤肥力[17]。
1.2 数据收集与整理气候数据来源于中国气象数据网(http://data.cma.cn/)的3.0中国地面气象站日值数据集1980—2010年的气候资料日值数据集(日最高温、日最低温、日降水量、日均风速)以及青藏高原科学数据中心共享的中国716个气象站太阳辐射日均值数据集1980—2010年的日均辐射数据[18]。利用Matlab软件, 提取气象站点数据, 并运用线性内插法对缺失值和错误值进行处理。运用ArcGIS 10.8的克里金插值法将以上5种日值数据集进行空间插值。
土壤数据来源于寒区旱区科学数据中心共享的面向陆面模拟的中国土壤数据集[19], 运用ArcGIS 10.8分层提取土壤剖面深度、土层厚度、砂粒、粉粒、粘粒、砾石百分比、容重、根密度、有机质和氮等数据, 同时参考了中国土壤数据库(http://vdb3.soil.csdb.cn/)对土壤颗粒组成和有效土层厚度进行调整。利用基于陆地生态系统生物地球化学循环的Century模型中的土壤计算器(Soil Calculator, https://www.nrel.colostate.edu/projects/century/soilCalculator.html)推算出各个土层的萎蔫系数和田间持水量, 其他土壤参数是通过查阅Brook 90水文模型(http://www.ecoshift.net/brook/b90doc.html)的相关文件进行计算获得的。
树种的最大积温和最小积温依据树种分布南界和北界的有效积温值, 并参考了中国植物志(http://frps.iplant.cn/)和前人在东北林区的研究成果[20—23]。树种的耐阴系数分为耐阴1和不耐阴2两个等级, 根冠比取值为0.5/0.8/1;滞叶周期分别取值为1/2/3, 耐氮系数也是在三个耐氮系数等级中取值, 最大寿命、耐旱系数、最低耐受温度、冠面积参数、树种的根冠比等来源于前人在中国东北地区温带森林的研究成果[24—26]。Richard方程参数B2、B3以及树木生长参数G的计算参考Botkin等人的相关研究[27]。
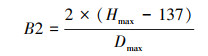
|
(1) |
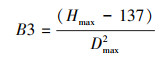
|
(2) |
式中, B2和B3分别是Richard方程中的相关参数, Hmax是树种的最大高度, Dmax是树种的最大胸径。
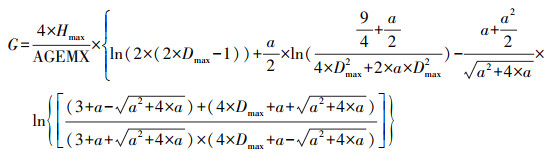
|
(3) |
式中, G是树木生长参数, a=1-137/Hmax, Hmax是树种的最大高度, Dmax是树种的最大胸径, AGEMX是最大寿命。公式中的Dmax和Hmax单位为cm, AGEMX单位为a。
| 树种 Species |
最大积温 Maximum degree day/℃ |
最小积温 Minimum degree day/℃ |
单位树冠 面积叶重 Leaf weight per unit crown area/ (100g/m2) |
最大寿命 Maximum age/a |
树木生 长参数 Tree growth parameter/cm |
耐旱系数 Shade tolerance |
最低耐 受温度 Temperature tolerated/℃ |
凋落物类型 Types of leaf litter |
叶片存 留周期 Foliage retention time/a |
| 红松Pinus koraiensis | 2600 | 800 | 440 | 400 | 118.93 | 0.276 | -35 | 12 | 2 |
| 水曲柳Fraxinus manshurica | 2700 | 900 | 440 | 250 | 119.59 | 0.189 | -31 | 2 | 1 |
| 蒙古栎Quercus mongolica | 3200 | 1100 | 440 | 300 | 90.92 | 0.514 | -34 | 9 | 1 |
| 春榆Ulmus davidiana | 2800 | 1000 | 440 | 250 | 105.35 | 0.176 | -33 | 5 | 1 |
| 色木槭Acer mono | 3100 | 1000 | 440 | 200 | 109.15 | 0.233 | -33 | 2 | 1 |
| 紫椴Tilia amurensis | 2600 | 1000 | 440 | 300 | 88.647 | 0.231 | -32 | 2 | 1 |
| 白桦Betula platyphylla | 3200 | 1100 | 250 | 140 | 173.04 | 0.412 | -37 | 4 | 1 |
| 山杨Populus davidiana | 3100 | 1000 | 250 | 140 | 148.02 | 0.333 | -35 | 7 | 1 |
图 1为LINKAGES 3.0模型基本流程图, 这是一个受气候和土壤条件限制的森林生态系统动态演替以及碳氮循环的一般模型。以面积为0.083 hm2的斑块为基本单元, 将演替过程中的关键要素进行耦合作为林分动态变化的驱动因素, 以年为模拟时间步长, 探究树种更新、生长、死亡动态变化, 循环累加到设定的时间尺度, 并输出每年斑块内树种组成和结构、斑块内总株树、树种生物量和蓄积量以及净初级生产力等, 进而实现对整个森林生态系统的动态模拟[11]。

|
| 图 1 LINKAGES3.0模型模拟基本流程图 Fig. 1 Basic flow chart of LINKAGES 3.0 model simulation |
图 1体现了该模型的设计原理, 树木与土壤可利用氮、干旱天数、有效积温以及可利用光的相互作用均取决于土壤条件和气候条件。模型的主要过程是树木的更新、生长和死亡, 这些过程受到立地条件的综合影响, 并且将会反作用于立地条件。总的来说, LINKAGES 3.0模型主要包括了四个大过程:决定单木生长的生理学过程, 决定林木种群动态的种群统计学过程, 决定氮有效性的微生物过程以及决定水分有效性的环境过程四种生物学过程的交互作用, 是一个综合考虑树木种群动态、水分养分循环以及气候之间的相互作用的模型。
1.3.2 LINAKGES 3.0基本结构LINKAGES 3.0模型将FORENA林分动态模块、Brook 90土壤水文模块、SUCSIM土壤养分模块和FORTNITE凋落物分解模块等进行耦合以实现模拟立地水平上种群动态、养分循环和水文过程之间的相互作用。LINKAGES 3.0模型包含以下七个子模块:温度子模块、湿度子模块、土壤分解子模块、累积子模块以及更新子模块、生长子模块、死亡子模块。其中温度子模块、湿度子模块、土壤分解子模块用于确定立地条件, 更新子模块、生长子模块、死亡子模块用于计算树木生长和群落动态, 两个部分通过累积子模块进行联结。温度子模块计算得出有效积温, 湿度子模块计算得出的干旱天数(低于土壤凋萎点的天数), 土壤分解子模块计算得出土壤可利用氮, 累积子模块将有效积温、干旱天数以及土壤可利用氮转化为相应的生长乘数。可利用的光根据个体树木的冠层结构计算得到。
1.3.3 模型中关键要素耦合的基本机理本研究通过图 2先介绍立地环境条件如何确定, 再说明模型的核心过程(更新、生长以及死亡)的模拟方法, 以探讨关键生态要素的耦合机制。

|
| 图 2 森林生态系统生态关键要素耦合框架图 Fig. 2 Frame diagram of coupling the key ecological elements of forest ecosystem DEGDF:有效积温生长乘数;SMGF:土壤水分生长乘数;SNGF:可利用氮生长乘数;ALGF:可用光乘数;G:树木生长参数;B2、B3:Richard生长方程的相关系数 |
立地环境条件由温度子模块、湿度子模块以及土壤子模块决定。其中温度子模块根据月均温度和指定的标准差计算得到有效积温。在湿度子模块中计算得出AET(实际蒸散量)以及干旱天数。AET根据Thornthwaite和Mather水分估算模型[28]得出, 该要素受到月均温度、月均降水、土壤理化性质以及树冠开闭程度的影响, 并影响着土壤理化性质以及腐殖质分解速率;干旱天数是指树木生长时土壤水分不足的天数, 在累积子模块转化为土壤可利用水分生长乘数, 该要素受到月降水、月均温、生长季开始日和结束日以及土壤理化性质(田间持水力和萎蔫系数等)等多生态要素的影响。月均温度, 月均降水由子模块LININT在不同气候年份之间进行线性插值。土壤子模块模拟土壤生物地球化学循环过程中的碳氮流动, 计算土壤可利用氮。该模块由两个主要部分组成:第一部分计算凋落物的氮固定和二氧化碳损失;第二部分计算腐殖质的氮矿化。凋落物的分解以及腐殖质的矿化受到实际蒸散量、凋落物化学成分和冠层覆盖度程度的影响。凋落物来自死亡子模块, 根据凋落物木质素和氮比例分为12种类型, 并匹配相应的分解和矿化速率;当凋落物的氮浓度达到该组的临界氮浓度时, 该组的有机物和氮含量会转移到土壤腐殖质池中。临界氮浓度是一个阈值, 低于该阈值被认为是氮限制, 高于则视为碳限制。AET、月均温以及树冠封闭程度均会影响氮矿化速率。
更新子模块模拟树种进入林分的过程。根据五个逻辑是非问题, 筛选出该年度适合种植的树种, 树种实际更新的量是最大更新值随着可用光、土壤湿度、有效积温低于树种最适条件而减少后的量。生长子模块通过生长乘数减少最大直径增量来计算得出每棵树实际直径增长量。每棵树的最大直径增量是根据树种参数G、B2和B3计算得到, 这三个参数均与树种最大高度以及直径有关。接着, 通过累积子模块将有效积温、干旱天数、土壤可利用氮以及可用光转化为有效积温生长乘数、土壤可利用水分生长乘数以及土壤可利用氮生长乘数。最后, 树种的最大直径增量会根据相对最不充足部分的生长乘数而减少, 从而得到实际的直径增量。死亡子模块显示树木有两种死亡方式, 一是与年龄有关的死亡率, 树种只有大约1%的个体能够存活到最大年龄;二是自然死亡率, 随着年龄的增长或生长速率放缓而增加。每年结束时, 叶、根系和木质凋落物会返回到土壤分解子中, 在下一年进行分解。
模型中的重要的双向耦合过程:(1)冠层的叶重量和叶面积决定了每棵树的光可用性, 进而影响树木的更新和生长;(2)土壤可利用氮影响树木生长, 进而通过凋落物的数量和类型影响分解。
2 结果 2.1 树种生物量随时间的变化随机布设100个点为模拟的林分斑块, 模拟总样地面积为8.3 hm2, 每个斑块从裸地开始模拟独立运行10次。由各树种生物量随演替时间的变化的图 3可知, 白桦、山杨在演替初期处于竞争优势地位, 随着演替的进行其生物量先增大后减少, 在70 a左右达到最大生物量, 在170 a左右完全消失;紫椴生物量随着演替的进行先增加后下降, 最后保持相对稳定的较高生物量;蒙古栎在演替的前60 a生物量增长的较慢, 在60—140 a左右, 蒙古栎生物量增加的较快, 之后生物量开始逐渐减少;水曲柳在前80 a, 生物量增速较大, 随后逐渐降低, 260 a之后维持较低且稳定的生物量;色木槭、春榆生物量先增后减少;红松在演替初期存在, 但生物量最少, 在80 a左右, 红松生物量才有存在感, 并随着演替的发生其生物量增速持续增大, 在280 a左右成为了生物量最高的优势树种。

|
| 图 3 树种生物量随时间的变化 Fig. 3 The change of tree species biomass with time |
在长白山森林生态系统的演替过程中, 各树种生物量的变化呈现较为明显的阶段性。白桦、山杨在演替前60 a左右占据优势, 生物量增长最快, 随演替的进行生物量先增大后减少, 在160—180 a左右完全消失。紫椴、蒙古栎水曲柳等在演替前期较少, 中期生物量较多, 最后逐渐减少甚至消失, 这些树种进一步的改变了生长环境, 为红松的迅速增长提供了合适的环境。顶级树种红松最开始处于劣势, 但随演替的发展生物量持续增加, 成为演替后期的主要树种。
2.2 代表性演替阶段的树木组成根据树种生物量变化明显的年份划分为四个阶段:0—70 a, 70—170 a;170—280 a;280—400 a。图 4以四个阶段为横坐标, 以树种在该时间段的生物量累积为纵坐标。根据图 4可知, 白桦、山杨, 在0—70 a, 生物量所占比例最大, 白桦比山杨生物量比例更大;在70—170 a, 白桦和山杨生物量比例降低, 此时两个树种的生物量相接近;在170 a之后, 两个树种没有生物量的存在;紫椴在70—170 a生物量占比最多, 随后所占比例逐渐减少, 但仍然占据第二大生物量;水曲柳在0—70 a生物量仅次于紫椴, 70—170 a生物量有所增加, 但在林分中所占比例下降, 随后生物量所占比例不断减少, 趋向于消失;蒙古栎在0—70 a生物量非常少, 所占比例最低, 70—170 a, 生物量迅速增多, 成为拥有第二大生物量所占比的树种, 170—280 a, 生物量有所降低, 但仍然是林分中第二大生物量的树种, 280 a后, 蒙古栎在林分中生物量占比继续降低;色木槭、春榆生物量在整个演替期间存在较少, 变化如倒U形;红松0—170 a生物量所占比例最小, 在170—280 a期间, 具有一定的生物量, 次于第二大生物量树种蒙古栎, 280 a之后, 生物量所占比例在所有树种中最大。

|
| 图 4 各阶段树种累积的生物量 Fig. 4 Accumulated Biomass of Tree species at different periods |
由图 5可知, 树种生物量所占比例从大到小排序, 在0—70 a, 白桦33%、山杨22%、紫椴16%、水曲柳14%、春榆6%、色木槭6%, 蒙古栎3%, 红松3%;在70—170 a, 紫椴31%、蒙古栎19%、水曲柳13%、白桦11%、山杨10%、色木槭8%、春榆4%、红松4%;在170—280 a, 紫椴36%、蒙古栎28%、红松15%、水曲柳10%、色木槭7%、春榆4%;在280 a之后, 红松50%、紫椴29%、蒙古栎11%, 水曲柳4%、色木槭3%、春榆3%。

|
| 图 5 各阶段树种生物量比例 Fig. 5 Proportion of tree species biomass at different periods |
由图 6可知, 地上树木总生物量的变化趋势为“先增加, 后减少, 最后趋于平缓”。在森林动态演替的0—70 a期间, 地上树木总生物量逐渐上升, 在末期增长到最大值181.25 mg/hm2;在70—250 a期间, 地上树木生物量逐渐减少至50 mg/hm2左右;在之后的演替期间, 地上总生物量在53 mg/hm2左右浮动。由图 7可知, 地上净初级生产力随演替时间的变化, 在0—30 a, 地上总净初级生产力迅速增加至11.42 kg/hm2, 在40—120 a期间, 地上净初级生产力的值不规则下降, 在之后的演替阶段, 地上净初级生产力值较为稳定, 在30 kg/hm2左右浮动。

|
| 图 6 地上总生物量的变化 Fig. 6 Changes in aboveground biomass |

|
| 图 7 地上净初级生产力的变化 Fig. 7 Changes in aboveground net primary productivity |
生命共同体中“水土气生”在生态系统尺度上体现为水分条件、土壤条件、气候条件(温度与光照)以及树木, 即本研究的有效积温、干旱天数、土壤可利用氮, 树木可利用光以及树木, 这些多关键要素是通过7个子模块以及各子模块内部相关方程耦合联系起来。有效积温、干旱天数和实际年蒸散量是根据月降水量、月均温度以及土壤的田间持水力和萎蔫系数来计算的, 土壤可利用氮是根据土壤有机质含量、碳化学成分、实际蒸散量和树冠层封闭度来计算的。每棵树的可利用光取决于所有更高树木的叶片生物量。有效积温、干旱天数、土壤可利用氮、可利用光以及树木相互作用、相互影响, 共同决定着每棵树木更新、生长、死亡过程。有效积温影响树木的更新, 并通过累积子模块生成的有效积温生长乘数影响树木直径生长量;干旱天数受到温度、降水、物候以及土壤质地类型等的影响, 同时也反作用于土壤的理化性质, 进而影响着树木动态过程;AET受到上述要素以及冠层的开闭程度的影响, 并且直接影响土壤分解模块中凋落物分解以及腐殖质矿化速率;土壤可利用氮的计算涉及土壤有机质含量、碳化学成分、实际蒸散量和树冠层封闭度, 对树木的生长、更新产生影响;每棵树的可利用光取决于树冠层所有更高树木的叶片生物量, 同样影响生长和更新过程。此外, 冠层结构决定着AET、土壤理化性质、分解速率以及矿化速率。树木死亡的概率随着年龄增加和生长缓慢而增加。每年结束时, 叶、根系和木质凋落物会返回到土壤(凋落物)中, 在下一年进行分解。树木的死亡分为自然死亡和年龄死亡两种。
整个模型的设计原理以及运行机制高度契合“生命共同体”的核心特征——“系统性、交互性、动态性、空间性”, 是一个高效、统筹、系统、综合的科学模型, 其设计原理以及运行机制在生态系统尺度对于“生命共同体”提供着理论支撑。
3.2 模型结果验证林窗模型结果的验证方法通常集中在两个方面:模型假设与结构的合理性验证以及模拟结果的真实性验证。模型假设与结构的合理性的方面在LINKAGES模型建立以来已经在北美以及中国东北林区等地得到了充分验证。最新版本LINAKGES 3.0在中国东北已有的林分基础上进行模拟, 得到了可靠的结果, 但该模型目前没有对针阔混交林的原生演替进行模拟[25—26]。因此本研究应用LINAKGES 3.0对长白山北坡平均海拔为800 m的红松针阔混交林进行模拟, 并通过与真实演替过程进行比较以及与其他林窗模型预测结果进行比较来验证其模拟结果的真实性。
与真实演替过程进行比较验证。已有的研究表明, 由裸地开始的真实演替过程通常呈现出一定的阶段性。我国学者将针阔混交林的演替过程总结为3个阶段:前期山杨白桦林阶段、中期阔叶红松林阶段和后期针阔混交林阶段[29]。本研究以现有真实气候作为气候情景输入, 对长白山北坡平均海拔为800 m的8个主要树种进行了由裸地开始的模拟。预测结果将总的演替动态概括为:阳性先锋树种阶段(0—70 a);软阔叶林阶段(70—170 a);硬混叶林阶段(170—280 a);针阔混交林阶段(280 a以后)。其结果与经典的演替理论以及经验性的观测规律基本相符。
与其他林窗模型预测结果的比较验证。国内已有许多关于红松针阔混交林动态过程模拟的研究成果, 这些成果可以为本研究提供模型结果的验证依据。但由于本研究的重点为关键要素耦合机制的探索, 因此只选择与本文研究地最为贴切的模型进行比较验证。邵国凡开发的KOPIDE模型对长白山北坡海拔650—850 m的针阔阔叶混交林进行了动态演替模拟[30]。图 8研究结果显示, 在演替开始的前50 a内, 山杨成为优势树种。随着山杨的地位趋于稳定或下降, 在50 a至约200 a间, 其他硬阔树种和红松在林分中的比例逐渐增加。这一时期的林分可以称为硬阔林。在200 a之后, 林分以红松为主, 标志着阔叶红松林的开始。演替过程与描述的红松林演替过程基本一致, 模拟与实际有较好地相符。

与以上的模型进行对比可以发现, 尽管在一些具体输出指标上存在差异, 例如林分总生物量峰值的出现时间、先锋树种的衰退时间等, 但模拟结果基本相似。因此, 可以确认本研究采用的LINKAGES3.0模型能够较好地模拟长白山针阔混交林的实际演替情况。
3.3 森林生态系统动态的模拟在掌握了真实演替过程规律的基础上, 应用林窗斑块分析了森林生态系统中的关键生态因素“水土气生”耦合机制, 以现有真实气候作为气候情景输入, 对长白山的主要8个树种进行了由裸地开始的模拟。预测结果表明, 在现实气候条件下, 以裸地为起点预测的森林演替过程与经典的演替理论以及经验性的观测规律基本相符[31]。总的演替动态可概括为:阳性先锋树种阶段(0—70 a);软阔叶林阶段(70—170 a);硬混叶林阶段(170—280 a);针阔混交林阶段(280 a以后)。长白山针阔混交林初生演替模拟的结果在一定程度上体现树种的特性:白桦和山杨为阳性树种, 在演替初期占据优势, 随着演替的进行, 林分郁闭程度变高, 而不耐阴的阳性树种白桦山杨逐渐衰退, 直至消失了;紫椴、水曲柳、蒙古栎均属于较为耐阴的阔叶树种, 所以在白桦山杨先锋树种创造了一定的荫蔽条件后, 开始从林下层不断生长;耐阴阔叶树种之间由于生理特性其生长速率也不同, 紫椴、水曲柳为软木, 而蒙古栎为硬木, 软木紫椴、水曲柳生长速率会高于硬木蒙古栎[32]。
在模型中将原先版本设定的5个耐阴性等级改为了1(不耐阴), 2(耐阴)两个等级, 这样的设定使得红松与其他阔叶树种的耐阴性在模型中相当, 但红松依旧能在后期占据优势地位, 主要是红松的最大寿命最高, 耐氮性、耐旱性等性能均强于其他树种。与其他的林窗模型比较, 该模型充分考虑了地下的生物地球化学循环, 对于生态要素考虑的更加统筹、综合。鉴于上述优点, 该模型应更多地被用于研究气候因素、土壤理化性质以及土壤碳氮循环对森林生态系统动态演替的影响。
3.4 不足与展望与输入月度或年度气候数据的林窗模型相比, 本研究采用的模型输入日值气候数据, 对极端降水和干旱更敏感。此外, 将水文模块更新到Brook 90水文软件的最新版本(4.7版本)[33], 考虑土层厚度以及每层土壤的理化性质, 对土壤的理化性质更加敏感。
但是, 本研究的林窗模型存在固有的局限, 它是一个理论经验的模型, 其设计基于一定的生态理论假设。应根据研究目的和研究对象, 考虑不同方法的可能性以及限制, 做出合适的假设, 有针对性地改进模型。本研究建议:学习FORCEEP模型将树冠模拟为“倒圆锥形”, 更加逼真地模拟树木对光的获取以及树木之间的竞争[34];将LINKAGSE 3.0与森林景观模拟模型LANDIS RRO耦合[35], 从不同尺度研究生态关键要素之间耦合机制以及森林动态;对土壤可利用氮、土壤可利用水和土壤有机物分解的过程模拟更加详细, 例如HYBRID[36]和FIRE-BGC[37];将“人”的活动考虑进去等, 例如动态植被模型(DVM)是探究人为驱动因素对森林动态影响的有价值的工具[38]。
4 结论以“生命共同体”的理念为指导思想, 本研究聚焦于生态系统尺度, 运用林窗模型将森林生态系统的关键要素“水土气生”进行耦合, 分析关键要素之间的耦合机制, 并探讨长白山针阔混交林树种组成和结构的动态变化。森林生态系统中的生态要素之间相互作用和相互影响:冠层特征决定树木可利用光, 并影响树木动态以及微生物分解速率等;土壤可利用氮由复合氮类型以及气候和土壤条件决定, 影响树木生长过程, 并通过凋落物的数量和类型影响微生物分解;气候条件不仅直接对森林动态演替过程产生影响, 也间接影响地下的营养循环。在400 a的初生演替期间, 白桦、山杨为演替的先锋树种, 最先侵入生境, 随演替的进行生物量先增大后减少, 最后消失;紫椴、蒙古栎、水曲柳等为过渡树种, 这些树种进一步改变了生长环境。顶级树种红松最开始处于劣势地位, 但随演替的发展, 其生物量持续增加, 成为演替后期的优势树种。经验证, 本研究对长白山森林生态系统动态的模拟可靠, 说明了关键要素“水土气生”在生态系统尺度耦合机制设定的合理性, 也将生命共同体“系统、统筹, 综合”的核心观念落于实处。为今后在生态系统尺度, 对“水土气生”多生态要素之间的耦合研究提供了科学依据。
| [1] |
赵金羽, 萨娜, 付晓, 郑拴宁, 吴钢, 何霄嘉, 陆兆华, 桑卫国. 以疏勒河流域为例的"山水林田湖草" 生态修复多尺度耦合框架及方法. 生态学报, 2023, 43(10): 3841-3854.
|
| [2] |
范应龙. 中国山水林田湖草研究进展综述. 中南林业调查规划, 2023, 42(3): 51-54, 72.
|
| [3] |
萨娜, 赵金羽, 寇旭阳, 郑拴宁, 陆兆华, 付晓, 何霄嘉, 吴钢, 桑卫国. "山水林田湖草沙生命共同体" 耦合框架、模型与展望. 生态学报, 2023, 43(11): 4333-4343.
|
| [4] |
吴浓娣, 吴强, 刘定湘. 系统治理--坚持山水林田湖草是一个生命共同体. 水利发展研究, 2018, 18(9): 25-32.
|
| [5] |
吴钢, 肖寒, 赵景柱, 邵国凡, 李静. 长白山森林生态系统服务功能. 中国科学: C辑: 生命科学, 2001, 31(5): 471-480.
|
| [6] |
罗唯学. 东北温带森林群落结构与生态功能的空间格局及其驱动因子. 北京: 北京林业大学, 2021.
|
| [7] |
Xie J Q, Zhang G Q, Li Y L, Yan X Y, Liu Q F, Chen D M, Sui M Z, He Y J. A bibliometric analysis of forest gap research during 1980-2021. Sustainability, 2023, 15(3): 1-16.
|
| [8] |
Morin X, de Coligny F, Martin-StPaul N K, Bugmann H, Cailleret M, Limousin J, Ourcival J, Prévosto B, Simioni G, Vennetier M, Guillemot J. Beyond forest succession: A gap model to study ecosystem functioning and tree community composition under climate change. Functional Ecology, 2021, 35(4): 955-975.
|
| [9] |
König L, Mohren F, Schelhaas M, Bugmann H, Nabuurs G. Tree regeneration in models of forest dynamics - suitability to assess climate change impacts on european forests. Forest Ecology and Management, 2022, 520(2): 120-390.
|
| [10] |
Greenwood S, Ruiz-Benito P, Martínez-Vilalta J, Lloret F, Kitzberger T, Allen C D, Fensham R, Laughlin D C, Kattge J, Bínisch G, Kraft N J B, Jump A S. Tree mortality across biomes is promoted by drought intensity, lower wood density and higher specific leaf area. Ecology Letters, 2017, 20(4): 539-553.
|
| [11] |
Post W M, Pastor J. Linkages: an individual-based forest ecosystem model. Climatic Change, 1996, 34(2): 253-261.
|
| [12] |
Dijak W D, Hanberry B B, Fraser J S, He H S, Wang W J, Thompson F R. Revision and application of the LINKAGES model to simulate forest growth in central hardwood landscapes in response to climate change. Landscape Ecology, 2016, 32(7): 1365-1384.
|
| [13] |
Shugart H H, Wang B, Fischer R, Ma J Y, Fang J, Yan X D, Huth A, Armstrong A H. Gap models and their individual-based relatives in the assessment of the consequences of global change. Environmental Research Letters, 2018, 13(3): 033001.
|
| [14] |
郝占庆, 李步杭, 张健, 王绪高, 叶吉, 姚晓琳. 长白山阔叶红松林样地(CBS): 群落组成与结构. 植物生态学报, 2008, 32(2): 238-250.
|
| [15] |
徐化成. 中国红松天然林. 北京: 中国林业出版社, 2001.
|
| [16] |
孔庆文, 黄利亚, 冯秀香. 长白山保护开发区自然环境分析. 农业与技术, 2007, 27(1): 70-74.
|
| [17] |
沈海燕. 不同类型红松林土壤物理性质及土壤有机质空间异质性研究. 哈尔滨: 东北林业大学, 2011.
|
| [18] |
Tang W J, Yang K, Qin J, Min M. Development of a 50-year daily surface solar radiation dataset over China. Science China Earth Sciences, 2013, 56(9): 1555-1565.
|
| [19] |
Shangguan W, Dai Y J, Liu B Y, Zhu A X, Duan Q Y, Wu L Z, Ji D Y, Ye A Z, Yuan H, Zhang Q, Chen D D, Chen M, Chu J T, Dou Y J, Guo J X, Li H Q, Li J J, Liang L, Liang X, Liu H P, Liu S Y, Miao C Y, Zhang Y Z. A China data set of soil properties for land surface modeling. Journal of Advances in Modeling Earth Systems, 2013, 5(2): 212-224.
|
| [20] |
He H S, Mladenoff D J, Crow T R. Linking an ecosystem model and a landscape model to study forest species response to climate warming. Ecological Modelling, 1999, 114(2/3): 213-233.
|
| [21] |
郝占庆, 代力民, 贺红士, David J. Malandnoff, 邵国凡. 气候变暖对长白山主要树种的潜在影响. 应用生态学报, 2001, 12(5): 653-658.
|
| [22] |
桑卫国, 陈灵芝, 马克平. 蒙古栎红松林演替模型FOROAK的研究. 植物学报, 1999, 41(6): 658-668.
|
| [23] |
于振良, 于贵瑞, 赵士洞, Steve Garman. 长白山阔叶红松林动态及经营管理模型研究. 资源科学, 2001, 23(6): 59-63.
|
| [24] |
浮媛媛. 中国东北林区主要树种地上生物量与密度的遥感估算与模拟研究. 长春: 东北师范大学, 2020.
|
| [25] |
刘凯. 气候变化与人为干扰对小兴安岭阔叶红松林树种组成和结构的影响. 长春: 东北师范大学, 2020.
|
| [26] |
桑卫国, 李景文. 小兴安岭南坡红松林动态模拟. 生态学报, 1998, 18(1): 38-47.
|
| [27] |
Botkin D B, Janak J F, Wallis J R. Some ecological consequences of a computer model of forest growth. The Journal of Ecology, 1972, 60(3): 849-872.
|
| [28] |
Thornthwaite C W, Mather J R. Instructions and table for computing potentialevapotranspiration and the water balance. Publications in Climatology, 1957.10: 183311.
|
| [29] |
李景文. 天然红松混交林恢复的基本对策. 林业月报, 1997, 6(4): 3-3.
|
| [30] |
Shao G F, Schall P, Weishampel J F. Dynamic simulations of mixed broadleaved-Pinus koraiensis forests in the Changbaishan biosphere reserve of China. Forest Ecology and Management, 1994, 70(1/2/3): 169-181.
|
| [31] |
葛剑平. 森林生态学建模与仿真. 哈尔滨: 东北林业大学出版社, 1996.
|
| [32] |
周以良, 董世林, 聂绍荃. 黑龙江树木志. 哈尔滨: 黑龙江科学技术出版社, 1986.
|
| [33] |
Schmidt-Walter P, Trotsiuk V, Meusburger K, Zacios M, Meesenburg H. Advancing simulations of water fluxes, soil moisture and drought stress by using the LWF-Brook90 hydrological model in R. Agricultural and Forest Meteorology, 2020, 291, 108-023.
|
| [34] |
Morin X, de Coligny F, Martin-StPaul N, Bugmann H, Cailleret M, Limousin J M, Ourcival J M, Prevosto B, Simioni G, Vennetier M, Guillemot J. Beyond forest succession: a gap model to study ecosystem functioning and tree community composition under climate change. . Functional Ecology, 2021, 35(4): 955-975.
|
| [35] |
Wang W J, He H S, Spetich M A, Shifley S R, Larsen D R, Fraser J S, Yang J. A large-scale forest landscape model incorporating multi-scale processes and utilizing forest inventory data. Ecosphere, 2013, 4(9): 1-22.
|
| [36] |
Friend A D, Stevens A K, Knox R G, Cannell M G R. A process-based, terrestrial biosphere model of ecosystem dynamics (Hybrid v3.0). Ecological Modelling, 1997, 95(2/3): 249-287.
|
| [37] |
Keane R E, Morgan P. Running-SW FIRE-BGC: A Mechanistic Ecological Process Model for Simulating Fire Succession on Coniferous Forest Landscapes of the Northern Rocky Mountains. USDA Forest Service Research Paper, 1996, 16(3): 484-122.
|
| [38] |
Maŕechaux I, Langerwisch F, Huth A, Bugmann H, Morin X, Reyer C P O, Seidl R, Collalti A, Dantas de Paula M, Fischer R, Gutsch M, Lexer M J, Lischke H, Rammig A Rödig E, Sakschewski B, Taubert F, Thonicke K, Vacchiano G, Bohn F J. Tackling unresolved questions in forest ecology: the past and future role of simulation models. Ecology and Evolution, 2021, 11(9): 3746-3770.
|