 2024, Vol. 44
2024, Vol. 44文章信息
- 张佳佳, 曾立雄, 雷蕾, 肖文发
- ZHANG Jiajia, ZENG Lixiong, LEI Lei, XIAO Wenfa
- 丛枝菌根途径的土壤有机碳固存机制研究进展
- Research progress of soil organic carbon sequestration mechanism by arbuscular mycorrhizal pathway
- 生态学报. 2024, 44(12): 4945-4955
- Acta Ecologica Sinica. 2024, 44(12): 4945-4955
- http://dx.doi.org/10.20103/j.stxb.202208112282
-
文章历史
- 收稿日期: 2022-08-11
- 采用日期: 2024-04-23

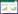




2. 南京林业大学南方现代林业协同创新中心, 南京 210037
2. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China
近几十年全球变暖一直是人类面临的最大环境问题之一。为了将全球变暖控制在与工业化前水平相比不超过2℃的范围内, 第21届联合国气候变化大会启动了4‰计划, 目标是将全球土壤有机质(Soil organic matter, SOM)储量每年增加0.4%, 以补偿人为造成的全球温室气体排放[1]。全球土壤含有2500 Gt的碳, 是大气碳库的3.3倍, 其中森林、草地和农业土壤碳库达1300 Gt左右, 约占全球土壤碳库的52%[2—3]。退化农田的土壤碳库每增加1 t/hm2, 小麦产量可增加20—40 kg/hm2, 玉米增加10—20 kg/hm2, 抵消了0.4—12 Gt的化石燃料排放, 即全球化石燃料排放的5%—15%[2]。因此, 增加陆地生态系统土壤有机碳(Soil organic carbon, SOC)含量是应对气候变暖的有效措施。
SOC的稳定来源一部分来自植物, 如含有软木脂、角质的植物残体, 甚至根系分泌物中一些有机酸(如草酸)可与铁铝氧化物结合形成稳定SOC组分, 作为稳定性碳源保存在土壤中[4—6]。另一部分来自微生物, 微生物碳泵(Microbial carbon pump, MCP)的“续埋效应”对SOC的形成和稳定具有重大作用, 其中真菌残体对SOC的贡献远大于细菌残体[7—11]。菌根真菌的菌丝能到达比根系更大的范围, 不仅利用其特性促进矿物结合有机质(Mineral-associated organic matter, MAOM)的形成[12], 而且可以暂时储存植被净初级生产力的1%—13%, 周转死亡后还成为真菌残体沉积在土壤中, 故被认为是土壤碳库之一[13—14]。因此研究菌根真菌途径的土壤碳固存机制对提高土壤碳固存潜力具有重要意义[15]。
丛枝菌根是分布最为广泛的一种菌根(高等植物的根系与菌根真菌形成互惠互利的共生体), 维管植物中约78%的菌根为丛枝菌根[15]。丛枝菌根真菌(Arbuscular mycorrhizal Fungi, AMF)从植物获取碳, 通过菌丝分泌、周转等途径又将碳归还到土壤, 形成AMF途径的土壤固碳机制, 是当下关注的一大热点[16—17], 但缺乏系统探讨AMF途径的SOC固存机制。在全球背景下, 如何理解AMF在土壤碳固存中扮演的角色是目前面临的重大科学挑战。因此, 本文主要以AMF的功能特性、AMF途径的SOC固存机制以及主要影响因素三方面阐明AMF途径的SOC固存研究进展, 旨在为菌根真菌提高生态系统碳汇的研究提供一定的理论参考。
1 AMF的功能特性 1.1 AMF是土壤碳库之一AMF有利于SOC的长期积累[18]。全球丛枝菌根植物每年分配给AMF的碳约为1.07 Gt, 其中分配给草本类植物的AMF碳量高于木本类植物约3倍[14]。AMF吸收碳后再贡献给SOM组分可达1%—11%[13, 19—20]。Drigo等人[21]也提出当大气中的CO2浓度升高时, 植物吸收的碳可快速转移给AMF, AMF再缓慢地释放给菌丝际(菌丝所能到达范围的土壤区域)的细菌和真菌群落, 起到固碳的作用。AMF能起到固碳作用主要有三方面原因, 首先, AMF的菌丝密度和周转速率远高于根系。AMF菌丝平均密度为2000 cm/cm3, 周转速率高达每月5次, 而细根密度的全球估计平均值仅为6.8 cm/cm3, 周转速率为每月0.07次[12, 22]。其次, AMF菌丝延伸影响土壤物理结构, 在土壤中形成通路, 与之相适配的微生物借助胞外聚合物在AMF通路上形成菌落, 利用AMF释放的源碳, 经代谢周转成为微生物源碳积累在土壤中[12, 23]。不仅如此, 菌根的菌丝对MAOM的结合位点远高于根系, 微生物源碳在MAOM中占主导地位, 故AMF的根外菌丝对土壤碳形成和稳定具有重要性[12, 24]。最后, AMF分泌的球囊霉素类相关蛋白(Glomalin-related soil proteins, GRSPs)及AMF周转残体, 不仅是SOC的稳定组成部分之一, 还通过其功能特性保护SOM不易被其他微生物分解[25—26]。因此, AMF的特性不仅对SOC的形成和稳定具有重要作用, 还是土壤碳库的重要组成部分。
1.2 植物与AMF形成营养共生关系大部分的被子植物都与AMF之间建立了营养获取的共生关系, 即AMF向植物提供超过50%的氮, 90%的磷以及部分钾、微量矿物质和水, 交换植物为其提供的4%—20%的糖和脂肪酸, 作为生长发育的碳源[27—29]。AMF从植物根系侵入, 在根内形成丛枝、根内菌丝、泡囊等结构, 菌丝再伸展至根外土壤形成大型菌丝网络[30]。AMF的菌丝薄, 能到达根系不能到达的地方, 增加了植物的营养吸收表面积[31]。如在低投入的农田, AMF接种后的薏米(Coix lacryma-jobi)磷含量显著增加[32];接种AMF的羊草(Leymus chinensis)氮素吸收增强, 氮含量提高107%[33]。
植物间AMF根外菌丝相互连接形成的地下共有菌丝网络(Common mycorrhizal network, CMN)[31], 促进植物种内和种间的物质交流, 提高稳定性和生产力[34]。如白三叶草(Trifolium repens)与枳(Poncirus trifoliata)的CMN存在激素交流, 促进双方生物量增加[35], 提高了植物的生产量。植物种间和种内还可以利用CMN进行资源再分配, 如树种群落之间通过CMN分配碳和营养物质给竞争优势寄主, 植物间出现“接受体”细根4%—29%的碳来源于“捐赠体”, 氮素转移可达2%—50%[36—38]。植物个体生长和生理性状也会响应CMN介导的资源再分配, 如香樟(Cinnamomum camphora)通过CMN提高了同一物种的叶片氮素摄取量, 促进其生物量积累, 降低了相邻构树(Broussonetia papyrifera)的根长、根表面积[39]。AMF与植物的营养共生促进植物的生产力增加, 反作用植物对AMF分配的碳量, 进而影响AMF途径的SOC固存。但目前对不同气候区或生态系统分配给AMF碳量的动态曲线研究甚少, 却是一个亟待突破的科学问题。
2 AMF途径对土壤碳库的作用机制 2.1 AMF对土壤碳形成的作用 2.1.1 植物碳是AMF对土壤碳输入的根本来源AMF位于植物根系和土壤之间, 通过根内菌丝从寄主植物获取碳源是控制植物碳进入土壤的一个关键输入点。AMF从根系获得碳源后, 经过菌丝分泌(如有机酸、GRSPs)或者残体释放到土壤中[40—44], 成为了AMF源碳进入土壤碳库。而且植物以糖和脂肪酸形式作为碳源供应给AMF进行菌丝生长和孢子萌发时, 脂类和几丁质是菌丝的结构主要成分[27, 29], 这两者也是SOC积累的重要成分[45]。因此, 在经过AMF代谢周转后释放的脂类和几丁质也是AMF途径的SOC积累来源之一, 但目前仍缺乏相关的追踪手段和量化其对SOC贡献的直接证据。
植物碳通过AMF进入土壤的另一重要途径为AMF根外菌丝参与凋落物分解。AMF缺乏植物细胞壁降解酶和过氧化物酶的基因, 不能直接通过酶作用分解凋落物, 故AMF利用菌丝体释放的碳诱导腐生微生物沉积于菌丝通路, 使腐生微生物分泌酶来降解凋落物, 将植物碳输入土壤[46—48]。
2.1.2 AMF源碳对土壤碳库的贡献AMF生物量、代谢产物和残体形成AMF源碳, 作用土壤碳库, 如草地土壤中AMF生物量产生碳速率为153 g m-2 a-1, 周转迅速(36—75 d), AMF残体周转较慢(每年1.4次)[49]。AMF生物量对颗粒有机质(Particular organic matter, POM)的贡献为1%—4%, 对MAOM贡献为5%—11%[20]。AMF分泌物中的GRSPs, 不仅在土壤中的周转时间为6—42 a, 对土壤碳库的贡献可达20%, 积累速率(约22.6%)高于SOC(15.9%), 还是一种持久性的有机胶结物质, 比其他的土壤化合物胶结土壤颗粒的能力强3—10倍[50—51], 因此GRSPs具有碳源和稳定碳库的双重作用。此外, 微生物残体碳是SOC形成和稳定的主要因素。微生物残体对温带农业、草地和森林土壤总SOC的贡献分别为55.6%、61.8%和30%, 其中真菌残体占了70%以上[42]。AMF的菌丝密度和周转速率远高于根系, 且生物量在草地和农田生态系统中占土壤微生物总生物量的很大比例, 但残体周转慢, 故AMF残体成为了真菌残体贡献了土壤微生物残体碳[7, 49]。
2.2 AMF的菌丝生长伴随其他碳源进入土壤碳库AMF的根外菌丝可以在土壤孔隙中自由移动, 为单细胞微生物提供一个可在其上移动的水化表面, 从而使微生物以菌落形式附着于矿物表面[12, 43]。土壤微生物中70%—90%是细菌, 细菌不仅与AMF协同提高凋落物的分解效率, 而且能够消耗菌丝损伤或死亡后释放的真菌渗出物, 并通过菌丝侵噬积极攻击活菌丝, 促进碳从生物形态向CO2、颗粒有机碳(Particular organic carbon, POC)和矿物结合有机碳(Mineral-associated organic carbon, MAOC)的转移[44, 52—53]。不仅如此, 植物碳经菌丝分泌进入微生物群落后, 存在一个微生物的食物网传递, 如微生物矿化碳源时, 真菌明显比其他微生物更早接触和分解细菌残体, 然后自身再作为碳源被下一级消费者所分解, 细菌和真菌的重叠利用底物和平行周转, 保证了MCP的续埋效应[54]。简单来说, 就是AMF与相适配的其他微生物形成微生物组进行土壤生物地球化学循环, 伴随着微生物相关产物分泌到土壤, 发挥MCP作用, 源源不断地将微生物碳输入土壤碳库, 从而提升了微生物固碳的潜力。另外, AMF与腐生真菌合作在促进高碳氮比的凋落物分解的同时, 可以抑制土壤碳的分解以及将分解的凋落物富集于大团聚体中[55—56], 实现土壤碳的净积累。故植物源碳也伴随着AMF的菌丝生长进入土壤碳库。
2.3 AMF途径对土壤碳的稳定作用团聚体是加强SOC保护的重要结构, 而且土壤团聚的能力比团聚的状态更能促进SOM的形成[57]。AMF对团聚体的形成和稳定具有重要作用[58]。AMF的菌丝是团聚体表面稳定网络的“主干”, 提供抗拉强度, 促进植物源碳沉积到团聚体矿物表面[59]。待微生物周转后又将碳输入孔隙, AMF产物GRSPs因其胶结作用, 将有机矿物结合形成团聚体以及改变团聚体表面极性来提高团聚体稳定性的作用[60], 将碳保护在团聚体中[61—62]。不仅如此, AMF及其相适配的微生物都会向周围环境释放有机化合物, 这些有机化合物与矿物质相互作用, 如菌丝在释放有机物或营养物质的胞外酶可以与粘土矿物形成稳定的键[12], 从而保护土壤碳的稳定。
总的来说, AMF从根系获取植物碳, 通过生物量、菌丝生长、代谢产物(尤其GRSPs)和残体以及与相适配的微生物相互作用, 分解凋落物及微生物的合成代谢、周转等复杂过程, 除了释放部分碳到大气中, 还将分解的植物残体和产生的微生物源碳输入土壤, 并形成团聚体结构保护土壤中的碳不被分解, 实现了大体上AMF途径的SOC固存内在机制(图 1)。

|
| 图 1 AMF途径的SOC固存机制概念图 Fig. 1 Conceptual diagram of soil organic carbon sequestration mechanism by the AMF pathway BD:土壤容重;pH:土壤酸碱度;N:氮;P:磷;C:碳;CO2:二氧化碳;CMN:共有菌根网络; (1)丛枝菌根结构示意图; (2)AMF的直接途径, 即AMF对土壤碳的形成和稳定的机制:AMF从根系获得碳源, 经菌丝分泌、GRSPs以及周转残体沉积在土壤中形成AMF源碳。菌丝和GRSPs有助于团聚体的形成和稳定, 从而保护碳不被分解; (3)AMF的间接途径, 即菌丝的生长伴随着其他源碳的输入:菌丝生长过程通过分泌碳吸引相适配的微生物形成菌落沉积在菌丝上, 通过分解凋落物以及微生物的合成代谢等过程, 产生植物源碳和微生物源碳沉积在矿物表面, 利于矿物结合有机质的形成; (4)—(7)分别为影响AMF途径SOC固存的因素(气候、土壤非生物因子、CMN以及人类活动); 箭头表示碳的流向 |
AMF对土壤碳库直接贡献的两个性状, AMF的菌丝和GRSPs, 会因气候变化、土壤类型、人类活动等因素产生差异。AMF根外菌丝固碳量可利用菌丝密度乘以一个常数计算, 如亚北极高山植物群落的AMF根外菌丝固定碳含量约50 g/m2[61]。所以AMF的菌丝密度在农田、草原和森林生态系统分别为0.5—8 m/g、2—10 m/g和2—34 m/g[59, 62—63], 不仅使AMF对SOC的直接贡献造成差异, 还能通过这些菌丝死亡, 经其他土壤微生物迅速分解其组织中的碳及内容物, 或者在土壤中停留数年至数十年[13], 从而影响土壤碳含量。GRSPs含量因土地利用类型的差异, 直接造成0—100 cm的土层总GRSPs含量在森林生态系统(约20 g/kg)最高, 草地(约18 g/kg)次之, 农田(13 g/kg)最低[64];GRSPs在传统耕作的农田生态系统的表层(0—10 cm)约为3.5 mg/g, 减少耕作后GRSPs含量约为4.1 mg/g, 但仍显著低于森林(6.5—11.8 mg/g)和草原(0.87—15 mg/g)的相对应土层的GRSPs含量[65—66]。虽然已有研究表明我国东北地区的农田生态系统80—100 cm的土层GRSPs对SOC的贡献是0—20 cm的1.34—1.5倍[67], 但不同生态系统、不同土层深度AMF直接对SOC贡献的研究甚少, 深入探索AMF对SOC贡献的时空分布特征对提高生态系统碳汇具有重要意义。
迄今为止, 有部分研究量化了AMF途径的总SOC固存量, 如氮沉降下, 在C3土壤上种植四年的C4植物经AMF途径对土壤新碳积累的贡献为28%, 约0.18 g/kg[65];经过增温三年, 草地通过AMF途径衍生的新碳有107 g/m2, 其中GRSPs贡献12%[49]等。因研究较少而不能系统探讨三大系统的AMF途径衍生碳量的差异以及影响因素, 但不同生态系统, 植物多样性、树种的合作与竞争如何调控AMF途径的SOC固存, 分别量化在不同环境下AMF直接、间接和总作用的差异及AMF自身激发效应等与土壤碳积累或损失的内在联系还是有待考究。
3.2 影响AMF途径SOC固存的主要因素 3.2.1 全球气候变化对AMF性状的调控气温上升、氮沉降等气候因素会影响AMF途径SOC的固存。具体来说, 变暖现象可分别显著提升草地19.1%与森林18.7%的AMF丰度, 但是当变暖超过4℃, 会对AMF丰度产生负效应[66]。温带草甸通过增加了AMF的孢子密度和直径、菌丝密度等响应温度升高和氮沉降[68]。大气CO2浓度升高显著增加了亚热带森林0—10 cm土壤的总GRSPs(约35%), 同时在氮添加的背景下, 土壤总GRSPs含量还能增加1.72%—48.49%[51]。降水也是影响GRSPs的一个因素, 如滨海生态系统中的GRSPs随着雨季的年降雨量升高而增加[69]。故气候变化可直接影响AMF自身性状来调控AMF途径的SOC固存。陆地上大约78%的维管植物为AM, 即至少78%的生态系统AMF途径SOC固存受到气候因子影响, 故可利用AMF对气候变化的响应策略来制定提高生态系统固碳潜力的策略。
3.2.2 土壤非生物因子影响AMF途径SOC固存GRSPs具有增加团聚体大小和稳定性、提高AMF存活率、保护菌丝、有助于碳固存等优点, 但其含量会受到土壤非生物因子的影响[67, 70]。土壤pH和容重会直接影响GRSPs含量, 如相比中性和钙质土壤, 酸性土壤中GRSPs含量最高[70], 同时电导率和土壤水分也可以间接通过pH和容重影响GRSPs含量[71]。此外, AMF的菌丝丰度会受到海拔、SOC和碱解氮等土壤因子的影响[25], 而AMF的菌丝丰度会影响GRSPs含量[72]。因此物理性质可以直接影响AMF性状来调控GRSPs对SOC的贡献。
除土壤物理性质外, 土壤养分限制也会影响AMF途径的SOC固存, 如AMF丰度和多样性对氮、磷素含量相当敏感[73];低磷环境会促进AMF对SOM的分解[74]。已有研究表明, AMF主导的森林和草地为将更多的碳储存在MAOC, 对土壤氮的需求高于外生菌根主导的生态系统[24], 而且微生物会重新利用微生物残体来响应土壤碳氮比失衡[75]。但是在不同生态系统中, AMF如何权衡植物-土壤响应养分变化, 并调控植物源碳、微生物源碳对SOC的贡献, 还需进一步研究。
3.2.3 土壤微生物与AMF的互相作用调控土壤碳动态变化AMF与根瘤菌的同时存在对植物的促进效果大于单一的AMF或根瘤菌, AMF和根瘤菌共存可增加植物的地上、地下及总生物量等[76—77], 而生物量的增加反过来影响植被对AMF分配的碳量。不仅如此, AMF与根瘤菌的共生还可以经其他因素调控微生物群落来增加或降低微生物残体的含量[78—79]。所以AMF和根瘤菌的共生可直接或间接的影响土壤碳固存。
木质素是植物材料中最复杂的成分, 也是SOM中难以降解的部分。AMF不能产生相关酶分解凋落物, 但是可以与腐生真菌合作, 借助腐生真菌产生的过氧化物酶和多酚氧化酶来降解凋落物或者分解SOM中的木质素, 从而调控SOC动态变化[48, 80]。腐生真菌与AMF互作引起土壤碳动态变化还需进一步探讨植物碳输入与土壤老碳分解、微生物生长代谢产生的呼吸和残体的联系。
3.2.4 CMN调节植物种群和群落间接影响AMF途径的SOC固存AMF介导的CMN通过资源再分配调节植物种群和群落的结构[34]。植物种内和种间通过CMN进行碳转移, 如地中海柏木(Cupressus sempervirens)通过CMN在同种内碳转移可达118.88%, 作为捐赠体可通过CMN转移62.87%的碳给乳香树(Pistacia lentiscus)。除碳以外, 植物间的营养物质竞争也会通过CMN优先分配给竞争优势个体, 如相邻的大须芒草(Andropogon gerardii)通过CMN会优先将矿物营养提供给那些最能固定碳或代表最强碳汇的寄主个体, 相邻个体之间的养分差异可高达32%[81—82]。虽然CMN存在碳和营养物质的不均等分配, 但也能维持竞争力差的植物, 如遮阴植物的生长[83]。因此共生植物对菌丝投资的碳量会根据植被光合能力所影响, 如通过13C标记量化高粱和亚麻共存下, CMN约70%的碳来自高粱, 30%来自亚麻[84]。也就是说, CMN能够均衡植物种间的竞争, 使植物间的共存增强, 从而调控对AMF菌丝的碳分配量。
但植物也会通过AMF扩大化感范围, 减少植物群落的多样性, 如胡桃(Juglans)利用AMF菌丝向外延伸分泌胡桃酮, 抑制番茄(Lycopersicon lycopersicum)的生长, 从而影响群落结构[85]。植物种群和群落变化可直接调节林分结构, 林分结构反过来影响植物的生长, 植物生长一方面通过调控对AMF菌丝的碳输入, 另一方面通过凋落物质量调控微生物分解来影响植物碳的输入, 再经过AMF途径的微生物周转产生残体, 进而影响植物源碳和微生物源碳的沉积以及MAOM和POM的形成和稳定[86—88]。因此从植物群落结构到调控AMF途径的碳输入以及该途径的微生物群落响应是探索AMF途径SOC固存的关键环节, 缺一不可。
3.2.5 人类活动对AMF途径SOC固存的干预人类活动如造林、耕作、退耕还林、再造林等对AMF途径SOC固存的影响是一个不可忽视的因素, 如耕作、施肥等措施会破坏AMF的菌丝网络和真菌多样性, 影响GRSPs含量[72, 89];天然林的根内球囊相关蛋白(Glomalin-related root protein, GRRP, 植物根内AMF产生的球囊蛋白)含量高于人工林, GRRP的增加伴随着GRSPs的增加, 进而促进SOC积累[90]等。相比于森林, 草地和农田是人为活动频繁的生态系统。传统耕作是农田长期的管理手段, 但耕作会破坏AMF的菌丝和生物量, 少耕或免耕可以提高AMF的生物量[91—92]。施肥是维持草地产量的一大经营措施, 但施肥会抑制AMF生物量, 导致土壤碳储量减少20%[93]。故寻求新的经营管理措施来代替传统的经营措施(耕作、施肥)有利于提高农业和草地生态系统的AMF介导的SOC固存, 从而缓解全球变暖问题。
4 总结与展望AMF与近80%的植物形成共生关系, 就决定了其对提高生态系统的固碳潜力具有重要意义。综上所述, AMF从植物根系获取碳源, 在其进行生长代谢和周转过程转化为AMF源碳进入土壤碳库, 伴随着与其他微生物的相互作用, 分解凋落物以及微生物的代谢周转, 增加了土壤碳库的微生物源碳和植物源碳的输入, 同时根外菌丝的生长和球囊霉素的胶结特性促进土壤团聚体的形成, 保留碳在土壤中, 即是AMF途径的SOC固存主要机制。气候变化、土壤非生物和生物因子、植物间的CMN以及人类活动等因素能够直接或间接地调控AMF途径的SOC固存, 但是在这个错综复杂的固碳过程, 仍有许多内在机理未探索清晰。比如:(1)尽管植被能够将光合产物分配给AMF, 但是不同气候带或生态系统中AMF能够获得的碳量时空动态曲线或者峰值仍不清楚。(2)不同生态系统AMF途径固存的SOC量在不同土层分布特征尚不清楚, 且探索AMF对土壤碳的直接、间接和总贡献差异与AMF途径的激发效应之间的联系, 对提高生态系统碳汇具有重要意义。(3)植物共存会影响分配给AMF菌丝的碳量, 系统探索植物群落结构调控AMF途径的植物碳输入以及微生物群落响应, 分析植物碳输入与土壤老碳之间的“收-支”动态, 有助于深刻理解AMF途径的SOC固存机制, 提高土壤固碳潜力。
| [1] |
Minasny B, Malone B P, McBratney A B, Angers D A, Arrouays D, Chambers A, Chaplot V, Chen Z S, Cheng K, Das B S, Field D J, Gimona A, Hedley C B, Hong S Y, Mandal B, Marchant B P, Martin M, McConkey B G, Mulder V L, O'Rourke S, Richer-de-Forges A C, Odeh I, Padarian J, Paustian K, Pan G X, Poggio L, Savin I, Stolbovoy V, Stockmann U, Sulaeman Y, Tsui C C, Vågen T G, van Wesemael B, Winowiecki L. Soil carbon 4 per mille. Geoderma, 2017, 292: 59-86. DOI:10.1016/j.geoderma.2017.01.002 |
| [2] |
Lal R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science, 2004, 304(5677): 1623-1627. DOI:10.1126/science.1097396 |
| [3] |
Panchal P, Preece C, Peñuelas J, Giri J. Soil carbon sequestration by root exudates. Trends in Plant Science, 2022, 27(8): 749-757. DOI:10.1016/j.tplants.2022.04.009 |
| [4] |
Shahzad T, Chenu C, Genet P, Barot S, Perveen N, Mougin C, Fontaine S. Contribution of exudates, arbuscular mycorrhizal fungi and litter depositions to the rhizosphere priming effect induced by grassland species. Soil Biology and Biochemistry, 2015, 80: 146-155. DOI:10.1016/j.soilbio.2014.09.023 |
| [5] |
Jeewani P H, Gunina A, Tao L, Zhu Z K, Kuzyakov Y, Van Zwieten L, Guggenberger G, Shen C C, Yu G H, Singh B P, Pan S T, Luo Y, Xu J M. Rusty sink of rhizodeposits and associated keystone microbiomes. Soil Biology and Biochemistry, 2020, 147: 107840. DOI:10.1016/j.soilbio.2020.107840 |
| [6] |
冯晓娟, 王依云, 刘婷, 贾娟, 戴国华, 马田, 刘宗广. 生物标志物及其在生态系统研究中的应用. 植物生态学报, 2020, 44(4): 384-394. |
| [7] |
Yang Y L, Xie H T, Mao Z, Bao X L, He H B, Zhang X D, Liang C. Fungi determine increased soil organic carbon more than bacteria through their necromass inputs in conservation tillage croplands. Soil Biology and Biochemistry, 2022, 167: 108587. DOI:10.1016/j.soilbio.2022.108587 |
| [8] |
Liang C, Schimel J P, Jastrow J D. The importance of anabolism in microbial control over soil carbon storage. Nature Microbiology, 2017, 2: 17105. DOI:10.1038/nmicrobiol.2017.105 |
| [9] |
梁超, 朱雪峰. 土壤微生物碳泵储碳机制概论. 中国科学: 地球科学, 2021, 51(5): 680-695. |
| [10] |
Buckeridge K M, Mason K E, Ostle N, McNamara N P, Grant H K, Whitaker J. Microbial necromass carbon and nitrogen persistence are decoupled in agricultural grassland soils. Communications Earth & Environment, 2022, 3: 114. |
| [11] |
Liang C. Soil microbial carbon pump: mechanism and appraisal. Soil Ecology Letters, 2020, 2(4): 241-254. DOI:10.1007/s42832-020-0052-4 |
| [12] |
See C R, Keller A B, Hobbie S E, Kennedy P G, Weber P K, Pett-Ridge J. Hyphae move matter and microbes to mineral microsites: integrating the hyphosphere into conceptual models of soil organic matter stabilization. Global Change Biology, 2022, 28(8): 2527-2540. DOI:10.1111/gcb.16073 |
| [13] |
Treseder K K, Holden S R. Ecology. Fungal carbon sequestration. Science, 2013, 339(6127): 1528-1529. DOI:10.1126/science.1236338 |
| [14] |
Hawkins H J, Cargill R I M, Van Nuland M E, Hagen S C, Field K J, Sheldrake M, Soudzilovskaia N A, Kiers E T. Mycorrhizal mycelium as a global carbon pool. Current Biology: CB, 2023, 33(11): R560-R573. DOI:10.1016/j.cub.2023.02.027 |
| [15] |
Brundrett M C, Tedersoo L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytologist, 2018, 220(4): 1108-1115. DOI:10.1111/nph.14976 |
| [16] |
Zhang Z L, Xiao J, Yuan Y S, Zhao C Z, Liu Q, Yin H J. Mycelium- and root-derived C inputs differ in their impacts on soil organic C pools and decomposition in forests. Soil Biology and Biochemistry, 2018, 123: 257-265. DOI:10.1016/j.soilbio.2018.05.015 |
| [17] |
Ma X M, Zhu B, Nie Y X, Liu Y, Kuzyakov Y. Root and mycorrhizal strategies for nutrient acquisition in forests under nitrogen deposition: a meta-analysis. Soil Biology and Biochemistry, 2021, 163: 108418. DOI:10.1016/j.soilbio.2021.108418 |
| [18] |
Verbruggen E, Veresoglou S D, Anderson I C, Caruso T, Hammer E C, Kohler J, Rillig M C. Arbuscular mycorrhizal fungi-short-term liability but long-term benefits for soil carbon storage?. New Phytologist, 2013, 197(2): 366-368. DOI:10.1111/nph.12079 |
| [19] |
Barto E K, Weidenhamer J D, Cipollini D, Rillig M C. Fungal superhighways: do common mycorrhizal networks enhance below ground communication?. Trends in Plant Science, 2012, 17(11): 633-637. DOI:10.1016/j.tplants.2012.06.007 |
| [20] |
Klink S, Keller A B, Wild A J, Baumert V L, Gube M, Lehndorff E, Meyer N, Mueller C W, Phillips R P, Pausch J. Stable isotopes reveal that fungal residues contribute more to mineral-associated organic matter pools than plant residues. Soil Biology and Biochemistry, 2022, 168: 108634. DOI:10.1016/j.soilbio.2022.108634 |
| [21] |
Drigo B, Pij A S, Duyts H, Kielak A M, Gamper H A, Houtekamer M J, Boschker H T S, Bodelier P L E, Whiteley A S, van Veen J A, Kowalchuk G A. Shifting carbon flow from roots into associated microbial communities in response to elevated atmospheric CO2. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(24): 10938-10942. |
| [22] |
Jackson R B, Mooney H A, Schulze E D. A global budget for fine root biomass, surface area, and nutrient contents. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(14): 7362-7366. |
| [23] |
Scheublin T R, Sanders I R, Keel C, van der Meer J R. Characterisation of microbial communities colonising the hyphal surfaces of arbuscular mycorrhizal fungi. The ISME Journal, 2010, 4(6): 752-763. DOI:10.1038/ismej.2010.5 |
| [24] |
Cotrufo M F, Ranalli M G, Haddix M L, Six J, Lugato E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nature Geoscience, 2019, 12: 989-994. DOI:10.1038/s41561-019-0484-6 |
| [25] |
Wang P, Wang Y, Shu B, Liu J F, Xia R X. Relationships between arbuscular mycorrhizal symbiosis and soil fertility factors in Citrus orchards along an altitudinal gradient. Pedosphere, 2015, 25(1): 160-168. DOI:10.1016/S1002-0160(14)60086-2 |
| [26] |
王建, 周紫燕, 凌婉婷. 球囊霉素相关土壤蛋白的分布及环境功能研究进展. 应用生态学报, 2016, 27(2): 634-642. |
| [27] |
Keymer A, Pimprikar P, Wewer V, Huber C, Brands M, Bucerius S L, Delaux P M, Klingl V, von Röpenack-Lahaye E, Wang T L, Eisenreich W, Dörmann P, Parniske M, Gutjahr C. Lipid transfer from plants to arbuscular mycorrhiza fungi. eLife, 2017, 6: e29107. DOI:10.7554/eLife.29107 |
| [28] |
Smith S E, Read D. Mycorrhizas in ecological interactions. Mycorrhizal Symbiosis. Amsterdam: Elsevier, 2008: 573.
|
| [29] |
Jiang Y N, Wang W X, Xie Q J, Liu N, Liu L X, Wang D P, Zhang X W, Yang C, Chen X Y, Tang D Z, Wang E T. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science, 2017, 356(6343): 1172-1175. DOI:10.1126/science.aam9970 |
| [30] |
田蜜, 陈应龙, 李敏, 刘润进. 丛枝菌根结构与功能研究进展. 应用生态学报, 2013, 24(8): 2369-2376. |
| [31] |
Bennett A E, Groten K. The costs and benefits of plant-arbuscular mycorrhizal fungal interactions. Annual Review of Plant Biology, 2022, 73: 649-672. DOI:10.1146/annurev-arplant-102820-124504 |
| [32] |
Bei S K, Xu M, Lyu X K, Chen C X, Li A L, Qiao X. Arbuscular mycorrhizal fungi enhanced coix responses to phosphorous forms but not for faba bean in intercropping systems, under controlled environment. Agronomy Journal, 2021, 113(3): 2578-2590. DOI:10.1002/agj2.20643 |
| [33] |
高志强, 谢腾龙, 艾俊国, 梅琳琳. 施氮下丛枝菌根真菌对羊草化学计量特征的影响. 中国草地学报, 2022, 44(3): 113-118. |
| [34] |
Tedersoo L, Bahram M, Zobel M. How mycorrhizal associations drive plant population and community biology. Science, 2020, 367(6480): eaba1223. DOI:10.1126/science.aba1223 |
| [35] |
Wu Q S, Zhang Y C, Zhang Z Z, Srivastava A K. Underground communication of root hormones by common mycorrhizal network between trifoliate orange and white clover. Archives of Agronomy and Soil Science, 2017, 63(9): 1187-1197. DOI:10.1080/03650340.2016.1276570 |
| [36] |
Avital S, Rog I, Livne-Luzon S, Cahanovitc R, Klein T. Asymmetric belowground carbon transfer in a diverse tree community. Molecular Ecology, 2022, 31(12): 3481-3495. DOI:10.1111/mec.16477 |
| [37] |
Wahbi S, Maghraoui T, Hafidi M, Sanguin H, Oufdou K, Prin Y, Duponnois R, Galiana A. Enhanced transfer of biologically fixed N from faba bean to intercropped wheat through mycorrhizal symbiosis. Applied Soil Ecology, 2016, 107: 91-98. DOI:10.1016/j.apsoil.2016.05.008 |
| [38] |
Fang L F, He X H, Zhang X L, Yang Y H, Liu R, Shi S M, Shi X J, Zhang Y T. A small amount of nitrogen transfer from white clover to Citrus seedling via common arbuscular mycorrhizal networks. Agronomy, 2020, 11(1): 32. DOI:10.3390/agronomy11010032 |
| [39] |
杨应, 何跃军, 董鸣, 王鹏鹏, 司建朋, 谢佩耘. 丛枝菌根网络对不同喀斯特适生植物生长及氮摄取的影响. 生态学报, 2017, 37(24): 8477-8485. |
| [40] |
Rillig M, Wright S, Nichols K, Schmidt W, Torn M. Large contribution of arbuscular mycorrhizal fungi to soil carbon pools in tropical forest soils. Plant and Soil, 2001, 233(2): 167-177. DOI:10.1023/A:1010364221169 |
| [41] |
Holátko J, Brtnický M, Kučerík J, Kotianová M, Elbl J, Kintl A, Kynický J, Benada O, Datta R, Jansa J. Glomalin—Truths, myths, and the future of this elusive soil glycoprotein. Soil Biology and Biochemistry, 2020, 153: 108116. |
| [42] |
Liang C, Amelung W, Lehmann J, Kästner M. Quantitative assessment of microbial necromass contribution to soil organic matter. Global Change Biology, 2019, 25(11): 3578-3590. DOI:10.1111/gcb.14781 |
| [43] |
Kakouridis A, Hagen J A, Kan M P, Mambelli S, Feldman L J, Herman D J, Weber P K, Pett-Ridge J, Firestone M K. Routes to roots: direct evidence of water transport by arbuscular mycorrhizal fungi to host plants. New Phytologist, 2022, 236(1): 210-221. DOI:10.1111/nph.18281 |
| [44] |
Kaiser K, Wemheuer B, Korolkow V, Wemheuer F, Nacke H, Schöning I, Schrumpf M, Daniel R. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Scientific Reports, 2016, 6: 33696. DOI:10.1038/srep33696 |
| [45] |
Dai G H, Zhu S S, Cai Y, Zhu E X, Jia Y F, Ji C J, Tang Z Y, Fang J Y, Feng X J. Plant-derived lipids play a crucial role in forest soil carbon accumulation. Soil Biology and Biochemistry, 2022, 168: 108645. DOI:10.1016/j.soilbio.2022.108645 |
| [46] |
Tang N W, San Clemente H, Roy S, Bécard G, Zhao B, Roux C. A survey of the gene repertoire of Gigaspora rosea unravels conserved features among glomeromycota for obligate biotrophy. Frontiers in Microbiology, 2016, 7: 233. |
| [47] |
Tisserant E, Malbreil M, Kuo A L, Kohler A, Symeonidi A, Balestrini R, Charron P, Duensing N, Frey N F D, Gianinazzi-Pearson V, Gilbert L B, Handa Y, Herr J R, Hijri M, Koul R, Kawaguchi M, Krajinski F, Lammers P J, Masclaux F G, Murat C, Morin E, Ndikumana S, Pagni M, Petitpierre D, Requena N, Rosikiewicz P, Riley R, Saito K, Clemente H S, Shapiro H, van Tuinen D, Bécard G, Bonfante P, Paszkowski U, Shachar-Hill Y Y, Tuskan G A, Sanders I R, Henrissat B, Rensing S A, Grigoriev I V, Corradi N, Roux C, Martin F. Genome of an arbuscular mycorrhizal fungus provides insight into the oldest plant symbiosis. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(50): 20117-20122. |
| [48] |
段嘉靖, 张勇, 郝龙飞, 王庆成, 剡丽梅, 何瑞雪. 丛枝菌根真菌和外生菌根真菌对凋落物分解的影响机制. 世界林业研究, 2022, 35(2): 21-27. |
| [49] |
Zhang J, Ekblad A, Sigurdsson B D, Wallander H. The influence of soil warming on organic carbon sequestration of arbuscular mycorrhizal fungi in a sub-arctic grassland. Soil Biology and Biochemistry, 2020, 147: 107826. DOI:10.1016/j.soilbio.2020.107826 |
| [50] |
Gomes V M, Assis I R, Hobbs R J, Fernandes G W. Glomalin-Related Soil Protein Reflects the Heterogeneity of Substrate and Vegetation in the campo rupestre Ecosystem. Journal of Soil Science and Plant Nutrition, 2021, 21(1): 733-743. DOI:10.1007/s42729-020-00396-7 |
| [51] |
Zhang J, Tang X L, He X H, Liu J X. Glomalin-related soil protein responses to elevated CO2 and nitrogen addition in a subtropical forest: potential consequences for soil carbon accumulation. Soil Biology and Biochemistry, 2015, 83: 142-149. DOI:10.1016/j.soilbio.2015.01.023 |
| [52] |
Yuan M M, Kakouridis A, Starr E, Nguyen N, Shi S J, Pett-Ridge J, Nuccio E, Zhou J Z, Firestone M. Fungal-bacterial cooccurrence patterns differ between arbuscular mycorrhizal fungi and nonmycorrhizal fungi across soil niches. mBio, 2021, 12(2): e03509-e03520. |
| [53] |
Boer W D, Folman L B, Summerbell R C, Boddy L. Living in a fungal world: impact of fungi on soil bacterial niche development. FEMS Microbiology Reviews, 2005, 29(4): 795-811. DOI:10.1016/j.femsre.2004.11.005 |
| [54] |
Zheng T T, Miltner A, Liang C, Nowak K M, Kästner M. Turnover of gram-negative bacterial biomass-derived carbon through the microbial food web of an agricultural soil. Soil Biology and Biochemistry, 2021, 152: 108070. DOI:10.1016/j.soilbio.2020.108070 |
| [55] |
Verbruggen E, Jansa J, Hammer E C, Rillig M C. Do arbuscular mycorrhizal fungi stabilize litter-derived carbon in soil?. Journal of Ecology, 2016, 104(1): 261-269. DOI:10.1111/1365-2745.12496 |
| [56] |
Wei L L, Vosátka M, Cai B P, Ding J, Lu C Y, Xu J H, Yan W F, Li Y H, Liu C X. The role of arbuscular mycorrhiza fungi in the decomposition of fresh residue and soil organic carbon: a mini-review. Soil Science Society of America Journal, 2019, 83(3): 511-517. DOI:10.2136/sssaj2018.05.0205 |
| [57] |
Even R J, Cotrufo M F. The ability of soils to aggregate, more than the state of aggregation, promotes protected soil organic matter formation. Geoderma, 2024, 442: 116760. DOI:10.1016/j.geoderma.2023.116760 |
| [58] |
Leifheit E F, Veresoglou S D, Lehmann A, Morris E K, Rillig M C. Multiple factors influence the role of arbuscular mycorrhizal fungi in soil aggregation—a meta-analysis. Plant and Soil, 2014, 374(1): 523-537. |
| [59] |
Xiang D, Verbruggen E, Hu Y J, Veresoglou S D, Rillig M C, Zhou W P, Xu T L, Li H, Hao Z P, Chen Y L, Chen B D. Land use influences arbuscular mycorrhizal fungal communities in the farming-pastoral ecotone of Northern China. New Phytologist, 2014, 204(4): 968-978. DOI:10.1111/nph.12961 |
| [60] |
Rillig M C, Mummey D L. Mycorrhizas and soil structure. New Phytologist, 2006, 171(1): 41-53. DOI:10.1111/j.1469-8137.2006.01750.x |
| [61] |
Soudzilovskaia N A, van der Heijden M G A, Cornelissen J H C, Makarov M I, Onipchenko V G, Maslov M N, Akhmetzhanova A A, van Bodegom P M. Quantitative assessment of the differential impacts of arbuscular and ectomycorrhiza on soil carbon cycling. New Phytologist, 2015, 208(1): 280-293. DOI:10.1111/nph.13447 |
| [62] |
Camenzind T, Rillig M C. Extraradical arbuscular mycorrhizal fungal hyphae in an organic tropical montane forest soil. Soil Biology and Biochemistry, 2013, 64: 96-102. DOI:10.1016/j.soilbio.2013.04.011 |
| [63] |
Ren H Y, Gui W Y, Bai Y F, Stein C, Rodrigues J L M, Wilson G W T, Cobb A B, Zhang Y J, Yang G W. Long-term effects of grazing and topography on extra-radical hyphae of arbuscular mycorrhizal fungi in semi-arid grasslands. Mycorrhiza, 2018, 28(2): 117-127. DOI:10.1007/s00572-017-0812-x |
| [64] |
Huang B T, Zhang L, Cao Y P, Yang Y R, Wang P, Li Z X, Lin Y. Effects of land-use type on soil organic carbon and carbon pool management index through arbuscular mycorrhizal fungi pathways. Global Ecology and Conservation, 2023, 43: e02432. DOI:10.1016/j.gecco.2023.e02432 |
| [65] |
Huang J S, Liu W X, Yang S, Yang L, Peng Z Y, Deng M F, Xu S, Zhang B B, Ahirwal J, Liu L L. Plant carbon inputs through shoot, root, and mycorrhizal pathways affect soil organic carbon turnover differently. Soil Biology and Biochemistry, 2021, 160: 108322. DOI:10.1016/j.soilbio.2021.108322 |
| [66] |
Hu H, He L Y, Ma H F, Wang J Y, Li Y, Wang J, Guo Y X, Ren C J, Bai H Y, Zhao F Z. Responses of AM fungal abundance to the drivers of global climate change: a meta-analysis. The Science of the Total Environment, 2022, 805: 150362. DOI:10.1016/j.scitotenv.2021.150362 |
| [67] |
Wang W J, Zhong Z L, Wang Q, Wang H M, Fu Y J, He X Y. Glomalin contributed more to carbon, nutrients in deeper soils, and differently associated with climates and soil properties in vertical profiles. Scientific Reports, 2017, 7: 13003. DOI:10.1038/s41598-017-12731-7 |
| [68] |
Zhang T, Yang X, Guo R, Guo J X. Response of AM fungi spore population to elevated temperature and nitrogen addition and their influence on the plant community composition and productivity. Scientific Reports, 2016, 6: 24749. DOI:10.1038/srep24749 |
| [69] |
Adame M F, Wright S F, Grinham A, Lobb K, Reymond C E, Lovelock C E. Terrestrial-marine connectivity: patterns of terrestrial soil carbon deposition in coastal sediments determined by analysis of glomalin related soil protein. Limnology and Oceanography, 2012, 57(5): 1492-1502. DOI:10.4319/lo.2012.57.5.1492 |
| [70] |
Agnihotri R, Sharma M P, Prakash A, Ramesh A, Bhattacharjya S, Patra A K, Manna M C, Kurganova I, Kuzyakov Y. Glycoproteins of arbuscular mycorrhiza for soil carbon sequestration: review of mechanisms and controls. The Science of the Total Environment, 2022, 806(Pt 2): 150571. |
| [71] |
Zhong Z L, Wang W J, Wang Q, Wu Y, Wang H M, Pei Z X. Glomalin amount and compositional variation, and their associations with soil properties in farmland, northeastern China. Journal of Plant Nutrition and Soil Science, 2017, 180(5): 563-575. DOI:10.1002/jpln.201600579 |
| [72] |
Gianinazzi S, Gollotte A, Binet M N, van Tuinen D, Redecker D, Wipf D. Agroecology: the key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza, 2010, 20(8): 519-530. DOI:10.1007/s00572-010-0333-3 |
| [73] |
Xiao D, Che R X, Liu X, Tan Y J, Yang R, Zhang W, He X Y, Xu Z H, Wang K L. Arbuscular mycorrhizal fungi abundance was sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in Karst ecosystems. Biology and Fertility of Soils, 2019, 55(5): 457-469. DOI:10.1007/s00374-019-01362-x |
| [74] |
Xu J, Liu S J, Song S R, Guo H L, Tang J J, Yong J W H, Ma Y D, Chen X. Arbuscular mycorrhizal fungi influence decomposition and the associated soil microbial community under different soil phosphorus availability. Soil Biology and Biochemistry, 2018, 120: 181-190. DOI:10.1016/j.soilbio.2018.02.010 |
| [75] |
Cui J, Zhu Z K, Xu X L, Liu S L, Jones D L, Kuzyakov Y, Shibistova O, Wu J S, Ge T D. Carbon and nitrogen recycling from microbial necromass to cope with C: N stoichiometric imbalance by priming. Soil Biology and Biochemistry, 2020, 142: 107720. DOI:10.1016/j.soilbio.2020.107720 |
| [76] |
高文礼, 陈晓楠, 伊力努尔·艾力, 马晓东. 不同水分处理下双接种丛枝菌根真菌和根瘤菌对疏叶骆驼刺生长及氮素转移的影响. 生态学报, 2022, 42(16): 6816-6826. |
| [77] |
刘倩, 高娅妮, 柳旭, 周文楠, 王佺珍. 混合盐碱胁迫下接种丛枝菌根真菌和根瘤菌对紫花苜蓿生长的影响. 生态学报, 2018, 38(17): 6143-6155. |
| [78] |
Schmidt J, Fester T, Schulz E, Michalzik B, Buscot F, Gutknecht J. Effects of plant-symbiotic relationships on the living soil microbial community and microbial necromass in a long-term agro-ecosystem. The Science of the Total Environment, 2017, 581/582: 756-765. DOI:10.1016/j.scitotenv.2017.01.005 |
| [79] |
Zhou R R, Liu Y, Dungait J A J, Kumar A, Wang J S, Tiemann L K, Zhang F S, Kuzyakov Y, Tian J. Microbial necromass in cropland soils: A global meta‐analysis of management effects. Global Change Biology, 2023, 29(7): 1998-2014. DOI:10.1111/gcb.16613 |
| [80] |
Frey S D. Mycorrhizal fungi as mediators of soil organic matter dynamics. Annual Review of Ecology, Evolution, and Systematics, 2019, 50: 237-259. DOI:10.1146/annurev-ecolsys-110617-062331 |
| [81] |
Weremijewicz J, da Silveira Lobo O'Reilly Sternberg L, Janos D P. Common mycorrhizal networks amplify competition by preferential mineral nutrient allocation to large host plants. New Phytologist, 2016, 212(2): 461-471. DOI:10.1111/nph.14041 |
| [82] |
Weremijewicz J, Janos D P. Common mycorrhizal networks amplify size inequality in Andropogon gerardii monocultures. New Phytologist, 2013, 198(1): 203-213. DOI:10.1111/nph.12125 |
| [83] |
Fellbaum C R, Mensah J A, Cloos A J, Strahan G E, Pfeffer P E, Kiers E T, Bücking H. Fungal nutrient allocation in common mycorrhizal networks is regulated by the carbon source strength of individual host plants. New Phytologist, 2014, 203(2): 646-656. DOI:10.1111/nph.12827 |
| [84] |
Walder F, Niemann H, Natarajan M, Lehmann M F, Boller T, Wiemken A. Mycorrhizal networks: common goods of plants shared under unequal terms of trade. Plant Physiology, 2012, 159(2): 789-797. DOI:10.1104/pp.112.195727 |
| [85] |
Achatz M, Morris E K, Müller F, Hilker M, Rillig M C. Soil hypha-mediated movement of allelochemicals: arbuscular mycorrhizae extend the bioactive zone of juglone. Functional Ecology, 2014, 28(4): 1020-1029. DOI:10.1111/1365-2435.12208 |
| [86] |
Ni Y Y, Jian Z J, Zeng L X, Liu J F, Lei L, Zhu J H, Xu J, Xiao W F. Climate, soil nutrients, and stand characteristics jointly determine large-scale patterns of biomass growth rates and allocation in Pinus massoniana plantations. Forest Ecology and Management, 2022, 504: 119839. DOI:10.1016/j.foreco.2021.119839 |
| [87] |
Bai Y F, Cotrufo M F. Grassland soil carbon sequestration: current understanding, challenges, and solutions. Science, 2022, 377(6606): 603-608. DOI:10.1126/science.abo2380 |
| [88] |
Feng X J, Wang S M. Plant influences on soil microbial carbon pump efficiency. Global Change Biology, 2023, 29(14): 3854-3856. DOI:10.1111/gcb.16728 |
| [89] |
Jeske E, Tian H, Hanford K, Walters D, Drijber R. Long-term nitrogen fertilization reduces extraradical biomass of arbuscular mycorrhizae in a maize (Zea mays L.) cropping system. Agriculture Ecosystems & Environment, 2017, 255: 111-118. |
| [90] |
Singh A K, Jiang X J, Yang B, Li H M, Liu W J, Singh N. Effect of root-glomalin on soil carbon storage in trees' rhizosphere and interspace of a tropical dry forest. Land Degradation & Development, 2021, 32(18): 5281-5291. |
| [91] |
Rosendahl S, Matzen H B. Genetic structure of arbuscular mycorrhizal populations in fallow and cultivated soils. New Phytologist, 2008, 179(4): 1154-1161. DOI:10.1111/j.1469-8137.2008.02535.x |
| [92] |
Säle V, Aguilera P, Laczko E, Mäder P, Berner A, Zihlmann U, van der Heijden M G A, Oehl F. Impact of conservation tillage and organic farming on the diversity of arbuscular mycorrhizal fungi. Soil Biology and Biochemistry, 2015, 84: 38-52. DOI:10.1016/j.soilbio.2015.02.005 |
| [93] |
Sochorová L, Jansa J, Verbruggen E, Hejcman M, Schellberg J, Kiers E T, Johnson N C. Long-term agricultural management maximizing hay production can significantly reduce belowground C storage. Agriculture, Ecosystems & Environment, 2016, 220: 104-114. |