 2024, Vol. 44
2024, Vol. 44文章信息
- 程鸿浩, 陈诗燕, 周夏芝, 邹运鼎, 毕守东
- CHENG Honghao, CHEN Shiyan, ZHOU Xiazhi, ZOU Yunding, BI Shoudong
- 害虫与天敌个体数量比值对天敌竞争作用指数的影响
- Impact of the population ratio between pest and predator on the competitive effect index among predators: taking Frankliniella intonsa and their major natural enemies as an example
- 生态学报. 2024, 44(11): 4914-4921
- Acta Ecologica Sinica. 2024, 44(11): 4914-4921
- http://dx.doi.org/10.20103/j.stxb.202311302613
-
文章历史
- 收稿日期: 2023-11-30
- 采用日期: 2024-05-15






2. 安徽农业大学 林学与园林学院, 合肥 230036
2. College of Forestry and Horticulture, Anhui Agricultural University, Hefei 230036, China
花蓟马Frankliniella intonsa是一种体型微小, 隐匿性强的茶树害虫, 在我国多地茶园均有分布, 主要危害茶花和茶树芽叶主脉两侧, 使叶质变差, 严重时可导致整叶变黄焦枯[1]。花蓟马不仅可以通过取食行为对茶叶进行为害, 更为严重的是可以传播病毒[2—3], 如不加以控制, 会对茶叶产量造成巨大损失。目前, 花蓟马的传统防治措施大多存在污染环境、方法单一和应用前景受限等弊端[4], 因此花蓟马的防治方式需要进一步创新。利用自然界天敌对害虫进行生物防治是一种安全、持久且经济的防治方法[1], 茶园花蓟马的常见天敌包括瓢虫、蝽类[5]和蜘蛛等, 张汉鹄等[1]报道蓟马被草间小黑蛛、瓢虫等捕食, 贺仲麟[6]指出蜘蛛是花蓟马的主要天敌, 胡金林[7]、陈樟福等[8]报道机敏漏斗蛛捕食蓟马、叶蝉等, 朱俊庆[9]报道茶园多种蜘蛛是蓟马的主要天敌。蜘蛛是花蓟马捕食类天敌中种类多、食性广, 食量大的一大类群, 是重要的生物防治资源。
竞争作用是物种为了生存, 对食物、空间等资源进行竞争的行为[10]。黄林等[11]研究轮虫与角枝类间的竞争作用, 总结得出温度和食物等均为竞争强度的影响因素;Park[12]在室内条件试验得出气候、寄生物和种群遗传性等均会影响拟谷盗不同品系间的竞争作用;郑全明[13]发现天敌斯氏钝绥螨对烟蓟马和二斑叶螨的种间竞争存在制约作用。由此可见, 外界环境、寄主资源、种群特性和天敌因素等均会影响昆虫种间竞争作用。竞争系数是生物间竞争作用的量化, 在昆虫生态学领域得到广泛应用[14—16]。
竞争作用在自然界普遍存在, 已有的文献多是研究环境及自身生物因素对昆虫种间竞争的影响, 以及一种天敌的竞争强度受目标物种群密度的影响[17—18], 多种天敌综合分析天敌-害虫比值与天敌竞争作用间相关性的研究少见报道, 在这种情况下, 对天敌捕食作用的研究就要涉及到天敌因取食产生竞争作用的复杂情况, Chen等[19]研究了天敌捕食小绿叶蝉时的竞争系数与竞争强度指数, 为方便简化研究竞争作用, 本文以茶园花蓟马及其数量较多的主要天敌(七种游猎蛛及两种瓢虫)为例, 并假设这9种天敌只捕食花蓟马, 研究天敌间的竞争作用指数(竞争系数和竞争强度指数), 并在此基础上, 分析花蓟马与天敌个体数量比值对竞争作用指数的影响, 明确了天敌在害虫-天敌个体数量比值不同时的竞争能力, 分析不同比值对每种天敌竞争作用的影响, 有利于合理评价和保护利用竞争力强的优势种天敌, 为花蓟马的生物防治提供科学依据。
1 材料与方法 1.1 调查地点与时间调查地点为安徽农业大学农业科技示范基地(31°94′N, 117°21′E), 选取“平阳特早”“农抗早”和“黄山大叶种”3个茶园进行调查, 茶园面积均为0.2 hm2, 每半个月调查一次。根据以往茶园害虫发生情况的研究, 主要采集花蓟马、卵形短须螨、小贯小绿叶蝉和茶尺蠖等主要害虫及鳞纹肖蛸、斑管巢蛛、三突花蟹蛛等主要天敌的种群数量, 本文主要研究9种天敌在捕食花蓟马时的竞争作用。花蓟马主要在秋冬季发生, 2020年自9月27日至11月20日共调查6次, 2021年自3月2日至11月19日共调查14次。花蓟马种群数量在各茶园的高峰日均在7月8日到11月20日之间, 茶园均按常规措施管理, 不施用化学农药。
1.2 茶树品种介绍“黄山大叶种”(“黄山早芽”), 由安徽省农业研究院茶叶研究所培育, 属灌木型大叶类, 叶形椭圆, 叶色黄绿[20];“平阳特早”, 属灌木型中叶类, 特早生种, 叶形椭圆, 叶色深绿[21];“农抗早”, 属灌木与小乔木型之间类型, 中叶类, 特早生种[22]。
1.3 调查取样方法茶园共5行, 为消除边缘群落影响, 采用平行跳跃法选取中间3行, 每行长50 m, 间隔1 m取2 m长的样方, 每行取10个样方, 共30个样方。用沾有洗衣粉溶液的搪瓷盘对样方中所有枝叶进行盘拍(搪瓷盘长40 cm, 宽30 cm), 调查和记载花蓟马及其天敌的种类和个体数。本研究以花蓟马数量高峰日时的种群数量及其9种天敌的种群数量作为数据资料。
1.4 分析方法 1.4.1 害虫-天敌比值分组依据选取3种茶园每年花蓟马种群数量最多的3次数据共18次数据作为研究内容, 并将该18次数据中数量最多的7种游猎蛛和2种瓢虫作为主要天敌, 对花蓟马与天敌个体数量比值按极差法进行三等份分组, 比值为0.00—2.00之间的属于低比值组(食饵相对不足), 2.01—4.00之间的属于中比值组(食饵相对充足), 大于4.00的属于高比值组(食饵相对宽裕)。
1.4.2 灰色系统分析灰色系统分析的基本思想是通过确定参考数据列和若干个比较数据列的几何形状相似程度来判断其联系是否紧密, 反映了曲线间的关联程度[23]。本研究将花蓟马数量与其9种天敌种群数量分别作为参考数据列与若干个比较数据列, 首先对数据进行无量纲化处理, 即采用均值化法将不同量纲的数据都统一归化到1这个量级附近, 其次利用公式计算关联系数ξi(k), 最后计算关联度ri。
关联系数公式为:
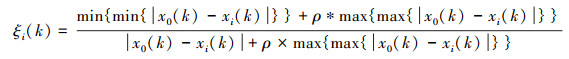
|
关联度公式为:
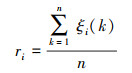
|
式中, x0(k)为k时害虫的种群数量;xi(k)为k时天敌i的种群数量;ρ为分辨系数, 一般取0到1之间, 本研究取0.8;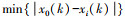
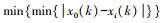
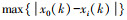
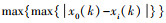
May将种间竞争系数公式定义为[24]:
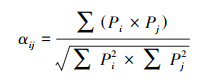
|
式中, αij表示物种i与物种j的种间竞争系数, Pi(Pj)表示物种i(j)在该资源状态下占物种i(j)种群数量之和的百分比。
1.4.4 竞争强度指数
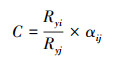
|
式中, 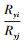
对于2个分类单位i与j之间的Pearson相关系数可定义为[27]:
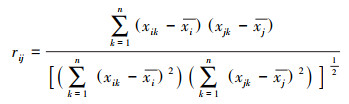
|
式中:xik和xjk分别为物种i和j在不同测量条件k下的种群数量;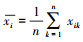
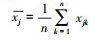
害虫数量多时的数据更能真实地反映害虫与天敌之间的关系[28], 选取2020年和2021年花蓟马种群数量均为峰值时的三次调查数据进行分析, 在20余种天敌中选择数量多的鞍型花蟹蛛Xysticus ephippiafus、三突花蟹蛛Misumenopstricuspidatu、粽管巢蛛ClubionaJaponicola、斑管巢蛛Clubiona reichlini、斜纹猫蛛Oxyopes sertatus、黑色跳蛛Blacksalticidae、条纹蝇虎Plexippus setipes等7种游猎蛛和龟纹瓢虫Propylaea Japonica、异色瓢虫Harmoniaaxyridis共9种天敌作为研究对象, 按极差法对18组数据进行高中低比值分组, 将结果列于表 1。
| 茶园 Tea garden |
日期 Date |
Y | X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | 害虫-天敌 种群数量比值 Pest-natural enemy population ratio |
组别 Groups |
| G1 | 2020-10-28 | 577 | 22 | 27 | 10 | 0 | 15 | 8 | 4 | 22 | 7 | 5.0174 | HR |
| 2020-11-10 | 213 | 34 | 29 | 7 | 1 | 10 | 5 | 24 | 6 | 0 | 1.8362 | LR | |
| 2020-11-20 | 865 | 15 | 32 | 7 | 0 | 10 | 3 | 6 | 3 | 1 | 11.2338 | HR | |
| 2021-07-08 | 116 | 56 | 30 | 10 | 23 | 26 | 29 | 0 | 32 | 32 | 0.4874 | LR | |
| 2021-11-01 | 749 | 16 | 14 | 6 | 3 | 20 | 5 | 10 | 5 | 1 | 9.3625 | HR | |
| 2021-11-20 | 87 | 1 | 2 | 3 | 5 | 26 | 2 | 4 | 0 | 1 | 3.9545 | MR | |
| G2 | 2020-10-28 | 177 | 5 | 14 | 13 | 1 | 42 | 6 | 2 | 2 | 0 | 2.0824 | MR |
| 2020-11-10 | 11 | 9 | 17 | 4 | 4 | 25 | 9 | 8 | 2 | 1 | 0.1392 | LR | |
| 2020-11-20 | 223 | 2 | 7 | 1 | 1 | 32 | 0 | 7 | 6 | 0 | 3.9821 | MR | |
| 2021-07-08 | 51 | 14 | 65 | 51 | 88 | 6 | 38 | 0 | 25 | 5 | 0.1747 | LR | |
| 2021-10-13 | 132 | 8 | 1 | 2 | 0 | 45 | 4 | 5 | 3 | 6 | 1.7838 | LR | |
| 2021-11-01 | 763 | 4 | 2 | 3 | 9 | 39 | 4 | 19 | 2 | 2 | 9.0833 | HR | |
| G3 | 2020-10-28 | 301 | 4 | 2 | 12 | 2 | 25 | 5 | 6 | 8 | 0 | 4.7031 | HR |
| 2020-11-10 | 144 | 6 | 3 | 3 | 0 | 23 | 2 | 10 | 2 | 1 | 2.8800 | MR | |
| 2020-11-20 | 214 | 1 | 8 | 2 | 0 | 32 | 0 | 9 | 0 | 1 | 4.0377 | HR | |
| 2021-07-08 | 312 | 26 | 46 | 24 | 24 | 22 | 7 | 0 | 61 | 41 | 1.2430 | LR | |
| 2021-11-01 | 254 | 5 | 2 | 8 | 3 | 2 | 2 | 3 | 0 | 1 | 9.7692 | HR | |
| 2021-11-20 | 87 | 7 | 2 | 0 | 1 | 4 | 3 | 4 | 0 | 1 | 1.9773 | LR | |
| Y: 花蓟马Frankliniella intonsa;X1:鞍型花蟹蛛Xysticus ephippiafus;X2:三突花蟹蛛Misumenopstricuspidatu;X3:粽管巢蛛ClubionaJaponicola;X4:斑管巢蛛Clubiona reichlini;X5:斜纹猫蛛Oxyopes sertatus;X6:黑色跳蛛Blacksalticidae;X7:条纹蝇虎Plexippus setipes;X8:龟纹瓢虫Propylaea Japonica;X9:异色瓢虫Harmoniaaxyridis;G1:黄山大叶种茶园Huangshan Dayezhong;G2:农抗早茶园Nongkangzao;G3:平阳特早茶园Pingyang Tezao;LR:低比值组Low-ratio group;MR:中比值组Median-ratio group;HR:高比值组High-ratio group | |||||||||||||
18组数据中, 花蓟马及其天敌比值属于高比值的占38%, 属于中比值的占24%, 属于低比值也的占38%, 高低比值组较中比值组占比较大, 表明花蓟马与天敌的种群数量始终保持差异悬殊, 两者种群数量存在显著负相关:天敌数量多时, 捕食作用强, 从而控制花蓟马种群数量;天敌数量少时, 对花蓟马的控制效应削弱, 从而花蓟马种群数量增加。
2.2 两年综合分析花蓟马与天敌数量比值对天敌竞争作用的影响对两年三个茶园的数据进行综合分析, 高比值组的比值均值为7.6010, 中比值组的比值均值为3.2248, 低比值组的比值均值为1.0917。将竞争系数和竞争强度指数的计算结果列于表 2。9种天敌竞争系数与花蓟马-天敌比值的Pearson相关系数分别为-0.4017、-0.9887、-0.0252、-0.9306、0.0945、-0.6003、0.4086、-0.3352和-0.5840, 除斜纹猫蛛和条纹蝇虎外, 竞争系数均随比值增加而降低, 9种天敌竞争系数均值与花蓟马-天敌比值的Pearson相关系数R为-0.7858, df=7时, R0.05=0.666, R>R0.05, 即9种天敌的竞争系数与花蓟马-天敌的比值呈负显著负相关。
| 项目 Item |
组别 Groups |
X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 |
| 竞争系数 | LR | 0.8562 | 0.8266 | 0.8009 | 0.8295 | 0.7018 | 0.8321 | 0.3963 | 0.7936 | 0.7621 |
| Competition coefficient | MR | 0.7386 | 0.7803 | 0.6102 | 0.6141 | 0.7981 | 0.6961 | 0.5991 | 0.5876 | 0.4245 |
| HR | 0.7886 | 0.5793 | 0.7570 | 0.5012 | 0.7302 | 0.7255 | 0.5142 | 0.6880 | 0.5040 | |
| 竞争强度指数 | LR | 0.9077 | 0.8447 | 0.8050 | 0.7969 | 0.7174 | 0.7570 | 0.388 | 0.8029 | 0.7915 |
| Competitive intensity | MR | 0.7782 | 0.8167 | 0.5327 | 0.6359 | 0.9528 | 0.6515 | 0.6402 | 0.6112 | 0.3099 |
| index | HR | 0.8691 | 0.5768 | 0.7573 | 0.4658 | 0.7205 | 0.7731 | 0.5383 | 0.6150 | 0.4924 |
竞争强度指数与比值的Pearson相关系数分别为-0.0980、-0.9732、0.0316、-0.9838、-0.1839、0.3132、0.4237、-0.7403和-0.4494, 除粽管巢蛛、黑色跳蛛和条纹蝇虎外, 竞争强度指数也随比值增加而降低, 9种天敌竞争强度指数均值与花蓟马-天敌比值的Pearson相关系数R为-0.8199, df=7时, R0.05=0.666, R>R0.05即9种天敌的竞争强度指数也与花蓟马-天敌的比值呈显著负相关, 与竞争系数的分析结果一致。
2.3 2020年花蓟马与天敌数量比值对天敌间竞争作用的影响分别计算2020年各天敌在不同比值组的竞争系数, 竞争系数随花蓟马-天敌比值的变化趋势如图 1所示, 竞争系数越大, 表示该天敌在9种天敌中的竞争能力越强。三突花蟹蛛、斑管巢蛛和斜纹猫蛛的竞争系数均在高比值组达到最小值, 其余六种天敌的竞争系数均在中比值组达到最小值, 9种天敌的竞争系数均在低比值组达到最大值。

|
| 图 1 2020年天敌竞争系数随花蓟马-天敌比值变化趋势 Fig. 1 Trend of natural enemy competition coefficient with the ratio of F. intonsa to natural enemies in 2020 X1:鞍型花蟹蛛Xysticus ephippiafus;X2:三突花蟹蛛Misumenopstricuspidatu;X3:粽管巢蛛ClubionaJaponicola;X4:斑管巢蛛Clubiona reichlini;X5:斜纹猫蛛Oxyopes sertatus;X6:黑色跳蛛Blacksalticidae;X7:条纹蝇虎Plexippus setipes;X8:龟纹瓢虫Propylaea Japonica;X9:异色瓢虫Harmoniaaxyridis |
2020年高比值组的比值均值为6.2480, 中比值组的比值均值为2.9815, 低比值组的比值均值为0.9877。通过相关性分析, X1、X2、X3、X4、X5、X6、X7、X8和X9的竞争系数与分组比值的Pearson相关系数分别为-0.5626、-0.9985、0.3277、-0.9190、-0.8707、-0.6467、-0.7751、-0.5308和-0.4374, 除粽管巢蛛外, 各天敌的竞争系数均与花蓟马-天敌比值呈负相关。再将9种天敌的竞争系数均值与分组比值进行相关性分析, 得出两者间Pearson相关系数R为-0.7391, df=7时, R0.05=0.666, R>R0.05, 即2020年天敌的竞争系数与花蓟马-天敌比值整体呈显著负相关。
计算2020年各天敌与花蓟马的灰色关联度, 由公式计算得到各天敌的竞争强度指数, 竞争强度指数随花蓟马-天敌比值的变化趋势结果如图 2所示。

|
| 图 2 2020年天敌竞争强度指数随花蓟马-天敌比值变化趋势 Fig. 2 Trend of natural enemy Competitive intensity index with the ratio of F. intonsa to natural enemies in 2020 |
2020年9种天敌竞争强度指数随花蓟马-天敌比值的变化趋势与竞争系数变化趋势相似, 斑管巢蛛、斜纹猫蛛的竞争强度指数均在高比值组最低。将竞争强度指数与各分组比值进行相关性分析, 得出9种天敌竞争强度指数与比值的Pearson相关系数R分别为-0.5660、-0.9269、-0.3256、-0.9172、-0.8737、-0.6401、-0.7819、-0.5359和-0.4486, 均为负相关。竞争强度指数均值与比值的Pearson相关系数R为-0.7819, df=7时, R0.05=0.666, R>R0.05, 即2020年天敌的竞争强度指数与花蓟马-天敌比值整体也呈显著负相关, 与竞争系数分析结果一致。
2.4 2021年花蓟马与天敌数量比值对天敌竞争作用的影响2021年对高比值组和低比值组的9种天敌进行竞争作用分析, 竞争系数、竞争强度指数在不同比值组的分布如图 3所示。

|
| 图 3 2021年9种天敌在各比值组的竞争强度指数和竞争系数值 Fig. 3 Competition intensity index and competition coefficient values of nine natural enemies in each ratio group in 2021 |
三突花蟹蛛、粽管巢蛛、龟纹瓢虫和异色瓢虫的竞争系数和竞争强度指数均在低比值组达到最大值, 即这四种天敌的竞争作用指数随花蓟马-天敌比值增加而减弱。鞍型花蟹蛛、斑管巢蛛、斜纹猫蛛和条纹蝇虎的竞争系数和竞争强度指数在高比值组达到最大值, 即这四种天敌的竞争作用指数随花蓟马-天敌比值的增加而加强。黑色跳蛛的竞争系数随比值增加而增加, 竞争强度指数随比值增加而减少, 说明考虑天敌与目标害虫密切程度会使黑色跳蛛竞争作用的量化产生差异。低比值组的X7(条纹蝇虎)种群数量较少是导致其竞争作用明显低于其他天敌的主要原因。综合9种天敌竞争系数和竞争强度指数均值分析, 2021年天敌的竞争作用与花蓟马-天敌比值呈正相关。2021年9种天敌竞争作用指数与比值的跟随关系不明显, 与平阳特早茶园中除9种主要天敌外其他花蓟马天敌的种群数量较多有关。
3 讨论关于天敌取食时的竞争作用, Chen等[19]研究了多种天敌取食小绿叶蝉的竞争系数与竞争强度指数, 本文在此基础上深入研究害虫与天敌个体数量比值对天敌竞争作用指数的影响, 明确了不同比值情况下天敌的捕食竞争情况。害虫(食饵)与天敌的数量比值可反映出天敌的食饵充足与否。食饵不足时, 天敌为获取有限的食物资源, 势必增加天敌间(包括种内和种间)的竞争压力, 产生扩大觅食范围或改变觅食策略的行为变化, 只有竞争力强的天敌才能获得较多的食饵, 从防治角度考虑, 应合理保护与利用这些竞争力强的天敌, 以便提高天敌对害虫的控制能力;食饵充足时, 生态系统趋于平衡, 天敌数量相应增加, 对高质量食饵的需求及挑剔等选择性取食也会增强部分天敌的竞争作用;食饵宽裕时, 由HollingⅡ模型可知当食饵数量增加到一定数量时, 捕食者数量会趋于平缓或饱和, 此时的食物资源丰富, 天敌更多专注于觅食和繁殖, 种间及种内竞争的强度也随之削弱。
关于物种间竞争作用, 应将注意力放在物种对其他资源的相对可利用性上[10]。本研究中提出的竞争强度指数与May的竞争系数的含义有很大的不同, 前者包含了天敌与害虫间数量关系的密切程度。May的竞争系数是考虑两种天敌竞争一种目标害虫的竞争力, 与两种天敌的数量、时空分布的占比有关, 但没有考虑天敌与目标害虫的在数量或时空上关系密切与否, 而本文在研究竞争系数时, 加入了两种竞争关系的天敌与目标害虫在数量上密切程度的影响因素。某天敌在与其他天敌竞争同一种害虫食饵时, 考虑该天敌与目标害虫数量关系密切程度(灰色关联度)以及其他参与竞争天敌与目标害虫数量关系密切程度, 若该天敌与目标害虫数量上关系密切, 其竞争力理应比其它参与竞争的天敌竞争力强。
为了合理利用和保护自然界中的天敌, 对目标害虫的天敌控害能力必须要进行充分的考虑, 天敌参与害虫防治时存在各种竞争作用差异, 从而对害虫的控害效果也存在显著不同。张安盛等研究了东亚小花蝽成虫[29]和中华草蛉幼虫[30]对西花蓟马的捕食反应, 发现两者的捕食作用率均随自身种群数量密度的增加而降低, 即食饵不足时竞争作用增强, 导致其摄食速率降低;吕文彦等[31]系统调查了麦田主要害虫及天敌的种群动态, 得出龟纹瓢虫在天敌间的竞争作用较为激烈, 从而对害虫的跟随与控制作用也较强;范江龙等[32]报道土壤中的拮抗菌由于较强的竞争能力, 对病原菌形成物理屏障或生态位排斥, 使病原菌最终不能感染寄主。Yu等[33]研究发现龟纹瓢虫及异色瓢虫在不同温度条件下的竞争作用不同, 且在食饵密度不同时的捕食速率也不同, 因此在施放天敌进行防治时应研究食饵密度, 以达到最佳控害效能。天敌竞争能力强弱直接影响其对害虫的控害能力, 研究害虫和天敌个体数量比值大小对天敌竞争能力的影响, 一是可以明确食饵多寡对竞争作用的影响, 二是可以评出竞争力强的优势种天敌, 种间竞争力强的天敌在害虫防治中可以有效地控制害虫增长, 这是害虫生物防治考虑的重要内容。
4 结论综合两年三种茶园的结果综合分析, 得出花蓟马主要天敌的竞争作用指数与花蓟马-天敌比值呈显著负相关, 比值越低, 竞争作用越强, 天敌由于有限的资源加强了竞争强度。害虫-天敌比值较高即害虫数量较多时, 接近田间为害的实际情况, 此时竞争能力强的天敌应受到合理保护与利用, 如本研究中的鞍型花蟹蛛。研究不同食饵密度下的天敌竞争作用, 有利于保护竞争能力强的天敌, 在实际生物防治中, 可根据花蓟马不同密度选择相应情况下竞争作用强的天敌, 从而达到更好的防治效能, 科学高效的开展花蓟马的生物防治工作。为研究方便, 本研究把7种游猎蛛及2种瓢虫均作为只取食花蓟马的单食性天敌进行研究, 实际上这9种天敌都是广食性天敌, 应进一步研究花蓟马与其他食饵共存时的天敌竞争作用, 这是一个新的研究课题, 有待于深入研究。
| [1] |
张汉鹄, 谭济才. 中国茶树害虫及其无公害治理. 合肥: 安徽科学技术出版社, 2004: 284-286.
|
| [2] |
Whitfield A E, Ullman D E, German T L. Tospovirus-thrips interactions. Annual Review of Phytopathology, 2005, 43: 459-489.
|
| [3] |
Gilbertson RL, Batuman O, Webster C G, Adkins S. Role of the insect supervectors Bemisia tabaci and Frankliniella occidentalis in the emergence and global spread of plant viruses. Annual Review of Virology, 2015, 2(1): 67-93.
|
| [4] |
杨磊, 邵雨, 李芬, 陈德鑫, 李方友, 吴少英. 缨翅目害虫蓟马生物防治的研究进展. 中国生物防治学报, 2021, 37(3): 393-405.
|
| [5] |
程娴, 余燕, 王建盼, 毕守东, 周夏芝, 邹运鼎, 王振兴, 李尚. 茶园天敌对花蓟马和茶短须螨的跟随效应研究. 植物保护, 2018, 44(6): 99-106.
|
| [6] |
贺仲麟. 烟草昆虫学. 合肥: 中国科学技术大学出版社, 2002: 117.
|
| [7] |
胡金林. 中国农林蜘蛛. 天津: 天津科学技术出版社, 1984.
|
| [8] |
陈樟福, 宋大祥. 机敏漏斗蛛的生活习性. 动物学杂志, 1980, 15(3): 14-15.
|
| [9] |
朱俊庆. 茶树害虫. 北京: 中国农业科技出版社, 1999: 156.
|
| [10] |
Southwood. Ecological research methods——applicable to the study of insect populations. Beijing: Science Press, 1984.
|
| [11] |
黄林, 席贻龙. 轮虫和枝角类的种间竞争及其影响因子研究进展. 生态学报, 2020, 40(19): 6720-6728.
|
| [12] |
Park T. Experimental studies on interspecies competition: Temperature, humidity and competition in two species of Tribolium. Physiol zool, 1954, 27: 177-238.
|
| [13] |
郑全明. 斯氏钝绥螨对烟蓟马与二斑叶螨共存时种间互作的影响[D]. 重庆: 西南大学, 2021.
|
| [14] |
马祖飞, 李典谟. 竞争指数及其在小红珠绢蝶保护生物学研究中的应用. 生态学报, 2002, 22(10): 1695-1698.
|
| [15] |
罗梅浩, 郭线茹, 郑晓军, 陈智, 马继盛. 烟青虫和棉铃虫在烟草上的生态位及其种间竞争. 中国烟草学报, 2002, 8(4): 35-38.
|
| [16] |
Ren Q L, Haseeb M, Fan J Y, Wu P X, Tian T Q, Zhang R Z. Functional response and intraspecific competition in the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects, 2020, 11(11): 806.
|
| [17] |
Papanikolaou N E, Dervisoglou S, Fantinou A, Kypraios T, Giakoumaki V, Perdikis D. Predator size affects the intensity of mutual interference in a predatory mirid. Ecology and Evolution, 2021, 11(3): 1342-1351.
|
| [18] |
赵英杰, 符成悦, 李维薇, 张立敏, 杜广祖, 陈斌, 李正跃. 异色瓢虫幼虫对草地贪夜蛾卵和低龄幼虫的捕食作用. 植物保护, 2020, 46(1): 51-54, 86.
|
| [19] |
Chen S Y, Cheng H H, Cai J J, Zou Y D, Cheng X, Bi S D. Interspecific competitive effects and annual differences in predation of Empoasca onukii (Hemiptera: Cicadellidae) by wandering spiders. Environmental Entomology, 2023, 52(6): 1048-1056.
|
| [20] |
江济和. 茶树新品种"黄山早芽" 选育报告. 茶叶, 1994, 20(4): 10-13.
|
| [21] |
林平, 赵东. 平阳特早茶研究. 茶叶, 1998, 24(4): 200-204.
|
| [22] |
杨维时, 程徽儿, 胡绍德. 茶树新品种"农抗早" 选育的理论与实践. 茶业通报, 2000, 22(4): 20-22.
|
| [23] |
邓聚龙. 灰色系统理论教程. 武汉: 华中理工大学出版社, 1990: 33-84.
|
| [24] |
May R M. Some notes on estimating the competition matrix, A. Ecology, 1975, 56(3): 737-741.
|
| [25] |
吴筱萌, 徐悦, 陈诗燕, 程鸿浩, 周夏芝, 邹运鼎, 毕守东. 游猎蛛在茶尺蠖高峰日的种间竞争作用. 环境昆虫学报, 2023, 45(5): 1385-1398.
|
| [26] |
陈诗燕, 程鸿浩, 吴筱萌, 徐悦, 邹运鼎, 毕守东. 捕食小贯小绿叶蝉的茶园游猎蛛的种间竞争作用研究. 植物保护, 2023, 49(4): 195-203, 210.
|
| [27] |
徐克学. 生物数学. 北京: 科学出版社, 1999: 89.
|
| [28] |
邹运鼎. 害虫管理中的天敌评价理论与应用. 北京: 中国林业出版社, 1997: 45-49.
|
| [29] |
张安盛, 于毅, 李丽莉, 张思聪, 门兴元. 东亚小花蝽成虫对西花蓟马若虫的捕食功能反应与搜寻效应. 生态学杂志, 2007, 26(8): 1233-1237.
|
| [30] |
张安盛, 李丽莉, 于毅, 门兴元. 中华草蛉幼虫对西花蓟马若虫的捕食功能反应与搜寻效应. 植物保护学报, 2007, 34(3): 247-251.
|
| [31] |
吕文彦, 秦雪峰, 杜开书. 麦田害虫与天敌群落动态变化研究. 中国生态农业学报, 2010, 18(1): 111-116.
|
| [32] |
范江龙, 李欣蕊, 席雪冬. 小麦赤霉病生物防治研究进展. 生物加工过程, 2021, 19(4): 420-431.
|
| [33] |
Yu X L, Zhang Y J, Zuo J F, Luo X, Zhang L, Danzeng Z M, Wang B, Xia P L, Zhang S Z, Liu T X, Feng Y. Rising temperatures affect the interspecific interference competition between Harmonia axyridis and Propylea japonica, and their predation rate on Myzus persicae. Journal of Pest Science, 2023, 96(2): 695-709.
|