 2024, Vol. 44
2024, Vol. 44文章信息
- 骆沁宇, 张梦园, 李晓璐, 范舒欣, 董丽
- LUO Qinyu, ZHANG Mengyuan, LI Xiaolu, FAN Shuxin, DONG Li
- 北京城市绿地不同生境自生植物多样性特征及其功能性状组成
- Diversity characteristics and functional trait composition of spontaneous plants in different habitats of urban green space, Beijing
- 生态学报. 2024, 44(11): 4744-4757
- Acta Ecologica Sinica. 2024, 44(11): 4744-4757
- http://dx.doi.org/10.20103/j.stxb.202311022387
-
文章历史
- 收稿日期: 2023-11-02
- 网络出版日期: 2024-03-16






气候变化与城市化对城市生物多样性带来了前所未有的挑战, 这引发人们对生物多样性保护的思考。城市绿地为城市生物提供了庇护所和栖息环境[1—2], 通过科学、合理地开展绿地与植物景观营建能够有效保护和提升城市生物多样性。然而, 在过去几十年, 由于城市绿地建设中过度使用栽培植物和人工景观, 大多数城市的植物种类和绿地景观呈现同质化[3—4], 亟需探索一条新的城市绿地建设路径, 使绿地发挥应有的生物多样性支持和保护功能。
城市绿地中分布着大量未经人工栽培而自发生长的植被, 即自生植物。这类植物种类丰富且能够在绿地中自发繁衍、生长, 不需要过多养护即可适应环境并形成自然野趣的景观, 受到越来越多学者的关注[1, 5—6]。作为城市植物多样性的重要组成部分, 自生植物可以为绿地中昆虫、鸟类和其他小型哺乳动物提供食源和遮蔽场所, 在城市生物多样性保护、支持和提升中扮演着重要角色[7—9]。随着这类植物的生态与景观价值逐渐被证实, 开展自生植物多样性的评估和保护工作成为城市生物多样性保护的重要环节。绿地中丰富的生境类型为不同种类植物的定居、生长提供了可能。人们根据景观审美偏好和功能需求在绿地中营造出不同的植物群落结构和生境条件, 多样化的生境类型能够有效提高绿地的物种丰富度[10—11]。前人研究已经证实, 一个生境类型丰富的绿地中支撑的自生植物物种数与一个城区相当[1]。然而, 并不是所有自生植物都能在各类环境中生存下来, 由于自生植物对外界环境适应性的不同, 不同的环境中形成了差异化的多样性特征[12—14]。目前已有研究从物种的角度阐述了不同生境之间自生植物多样性的差异[15—16], 我们需要一个基于个体、群落与环境的整体认知, 以更好地理解这些多样性特征的形成过程和维持机制。
功能性状是植物在长期进化过程中形成的, 它通过影响植物的营养生长、繁殖扩散进而影响植物的发生, 能够将植物个体、群落和外部环境紧密联系起来[5, 17]。具有特定功能性状的物种能够通过环境因子的过滤得以定居、生长。已有研究发现, 物种在城市环境中的分布与自身功能性状相关。高干扰环境中的植物物种以种子重量重、生活周期短的一年生植物为主[18—19];陆地与水域交界处分布有更多依靠风力授粉和扩散的物种[5]。植物的功能性状反映了不同物种的生态适应性差异, 直观体现在群落水平的多样性特征上[20]。然而, 目前较少研究关注功能性状在城市绿地的异质化生境中的分布情况。基于此, 本文提出假设, 城市绿地不同生境中自生植物的功能性状组成存在差异, 这些差异与自生植物群落水平的多样性相关, 反映了群落水平的物种聚集状况。
本研究从植物起源、营养生长、繁殖扩散等方面分析了城市绿地的8类生境中自生植物的物种和功能性状的特征, 主要探索以下3个问题:(1)城市绿地不同生境类型中自生植物的多样性特征有何差异?(2)这些差异在功能性状组成上是如何体现的?(3)在特定生境类型中, 即存在相似环境过滤器的条件下, 自生植物在群落水平的多样性与功能性状组成是否存在特定的关系?本文尝试通过在特定生境中建立群落水平自生植物多样性与功能性状组成之间的关系, 初步探索植物个体、群落、生境间的相互作用, 更好地理解不同生境类型中自生植物多样性的形成过程和维持机制, 以期切实指导城市绿地植物景观设计中自生植物生境的营建与管理。
1 研究方法 1.1 研究区域与样地设置北京地处华北平原的北部(北纬39°28′至41°5′, 东经115°25′至117°30′), 属暖温带半湿润半干旱季风气候, 四季分明, 夏季高温多雨, 冬季寒冷干燥, 最低温可达-20℃以下, 年均降雨量630mm左右, 且主要集中在夏季。行政辖区总面积为16410km2, 其中, 中心城区总面积约1378km2。快速城市化导致北京市建设用地扩张趋势明显, 沿环线由中心向四周扩散, 整体上呈现出“摊大饼”式的空间格局[21]。至2022年, 北京城六区绿化覆盖率达45.00%, 绿地面积达4.4×104hm2。仅2023年以来, 全市已完成“揭网见绿”927.09hm2, 绿化建设成效显著。
本研究选取北京市主城区20个覆盖丰富生境类型城市绿地作为研究样地, 排除郊野公园、生态公园、自然(类)公园等自然植被较多、养护管理强度低的绿地。为排除土地利用历史对自生植物的影响, 本研究选择建于2000年之后的城市绿地。在各样地内部采用网格取样结合典型取样法进行植物样方设置。在全面踏查样地的基础上, 根据主要植被类型、下垫面类型以及与水体、园路的位置关系, 将样地生境划分为8类:常绿落叶林地、常绿林地、落叶林地、灌丛、草地、水体、行道树种植带、硬质铺装区域。在自生植物丰富的区域设置样点, 保证样点覆盖全部生境类型(表 1)。以样点为中心设置10m×10m的木本样方, 在木本样方中心及四个角点处设置1m×1m的草本样方。对于道路边缘等狭长形区域, 按照100m2设置条形样方, 小样方按照1m2进行设置。共调查木本样方230个, 草本样方1150个。
| 生境类型 Habitat types |
描述 Description |
样点数/个 Points |
| 常绿落叶林地(ED) Evergreen and deciduous woodland |
由常绿树种和落叶树种组成的混交林区域, 以乔木为主;不与水体相邻。 | 52 |
| 常绿林地(EW) Evergreen woodland |
由常绿树种组成的林地区域, 以乔木为主;不与水体相邻。 | 23 |
| 落叶林地(DW) Deciduous woodland |
由落叶树种组成的林地区域, 以乔木为主;不与水体相邻。 | 71 |
| 灌丛(SH) Shrubland |
以灌木为优势植被类型的区域, 灌木覆盖度≥50%;不与水体相邻。 | 24 |
| 草地(GR) Grassland |
树木郁闭度<0.1, 生长草本植物为主的区域;不与水体相邻。 | 33 |
| 水边(WA) Waterside |
与绿地内水体相邻的种植区域。 | 10 |
| 行道树种植带(TP) Tree pool |
与园路相邻, 种植行道树的带状区域, 垂直路缘的宽度<3m。 | 11 |
| 硬质铺装区域(IP) Impervious pavement |
下垫面为硬质铺装, 供人群活动的非种植区域。 | 6 |
于2022年6月至8月开展自生植物调查。记录木本样方内自生木本的物种名, 各物种株数、盖度。记录草本样方内自生草本植物的种类, 各物种株(丛)数、盖度、平均株高。本次调查记录到的木本植物高度低于40cm, 均为自发生长而非人工栽植的木本小苗, 定义为自生植物纳入自生植物群落进行分析。
1.2.2 功能性状组成与功能多样性从物种发生和植物群落建立的角度, 选取与物种来源、营养生长和繁殖扩散相关的16类性状进行研究。性状数据通过实地调查记录以及查阅工具书[22]、文献[23—27]与开放数据库获取[28—29], 包括《北京植物志》、全球植物性状数据库(Plant Trait Database, TRY v6.0)等(表 2)。其中, 本地植物指自然分布于北京地区的物种;外来植物指自然分布于北京范围以外, 由人类有意或无意引入北京地区的物种, 根据自然分布区域分为国内外来植物和国外外来植物。对于在数据库和文献中无法获取且难以通过野外调查得到的物种数据, 本研究使用同属中近缘种的数据表示。
| 性状 Traits |
性状类型 Trait type |
性状状态 Trait state |
数据来源 Data source |
| 生活型 Life form |
名义型 | 一年生草本;二年生草本;多年生草本;藤本;木本; | 《北京植物志》;《中国植物志》 |
| 物种来源 Origin |
名义型 | 本地植物;国内外来植物;国外外来植物;入侵植物 | 《北京植物志》;《中国植物志》;肖翠等[23];崔夏等[24] |
| 地理成分 Floristic zone |
名义型 | 世界分布;温带分布;热带分布 | 吴征镒[25] |
| 植株高度 Plant height |
连续型 | 以厘米(cm)为单位的同一物种植株主茎顶部到基部的平均距离 | 实地调查记录 |
| 比叶面积 Specific leaf area |
连续型 | 以平方毫米每毫克(mm2/mg)为单位的叶片单面面积与其干重之比 | TRY v.6.0 |
| 光合作用途径 Photosynthetic pathway |
名义型 | C3光合途径;C4光合途径 | TRY v.6.0;翟增康[26] |
| 花色 Flower color |
名义型 | 花被不明显;绿色系;白色系;蓝紫色系;黄色系;红色系;两种及以上花色 | 《北京植物志》;《中国植物志》;实地调查记录 |
| 始花期 Flowering start month |
数值型 | 植物开始开花的月份, 1—12月 | 《北京植物志》;《中国植物志》 |
| 开花持续期 Florescence |
数值型 | 从始花期至末花期之间的时期, 按月份计算, 1—12月 | 《北京植物志》;《中国植物志》 |
| 授粉方式 Pollination mode |
名义型 | 虫媒授粉;风媒授粉;自体授粉; | TRY v.6.0 |
| 果实开裂类型 Fruit dehiscence type |
名义型 | 闭果;裂果 | 《北京植物志》;《中国植物志》;黎维平[27] |
| 果皮类型 Fruit pericarp type |
名义型 | 干果;肉果 | 《北京植物志》;《中国植物志》;黎维平[27] |
| 始果期 Fruiting start month |
数值型 | 植物开始结果的月份, 1—12月 | 《北京植物志》;《中国植物志》 |
| 果实持续期 Fruiting stage |
数值型 | 从始果期至末果期之间的时期, 按月份计算, 1—12月 | 《北京植物志》;《中国植物志》;实地调查记录 |
| 种子干重 Seed dry mass |
连续型 | 以毫克(mg)为单位的种子干燥状态下的重量 | TRY v.6.0;英国邱园皇家植物园种子数据库 |
| 种子扩散方式 Dispersal mode |
名义型 | 自体扩散;风扩散;动物扩散;水扩散; | TRY v.6.0;英国邱园皇家植物园种子数据库 |
| TRY v.6.0: 全球植物性状数据库 | |||
为评估物种多样性, 选取Patrick丰富度指数(R)、Simpson多样性指数(D)、Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)[30], 使用R v.4.2.2中的‘vegan’包进行计算。
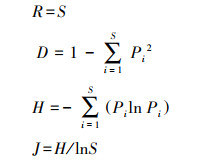
|
式中, S为样方中记录到的物种数, Pi为物种i的相对多度。
1.3.2 功能多样性计算为评估功能多样性, 选取功能丰富度指数(Functional richness, FRic)、功能均匀度指数(Functional evenness, FEve)、功能分歧度指数(Functional divergence, FDiv)以及Rao二次熵指数(Rao′s quadratic entropy, FDQ)[31], 使用R v.4.2.2中的‘FD’包进行计算。
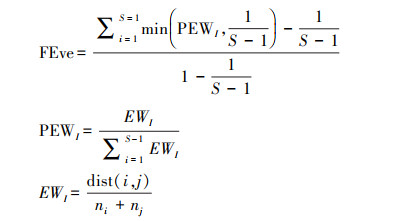
|
式中, S为物种数, EWI表示均匀度权重, PEWI为分支长权重, I为分支长, dist(i, j)表示物种i和j的欧氏距离, ni表示物种i的数目。

|
式中, dij为物种i与物种j的功能性状距离, Pi和Pj分别是一个样方中物种i和物种j的相对丰度;Xik和Xjk分别代表物种i和物种j在一个样方中的功能性状k, n代表功能性状的数量。
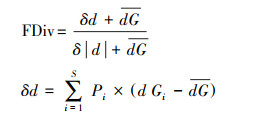
|
式中, dGi是欧氏距离, 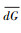
选取群落加权平均值(Community weighted mean, CWM)作为群落水平功能性状组成的量化指标[31], 将名义型性状按性状状态转化为二元数据进行计算。使用R v.4.2.2中的‘FD’包进行计算。
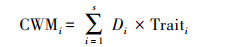
|
式中, CWMi表示所测物种i的功能性状的群落加权平均值, Di代表物种i的相对多度, Traiti代表物种i的平均功能性状值。
1.3.4 统计分析使用Excel2016对数据进行整理, 并在SPSS和R v.4.2.2软件中进行数据的统计分析。采用Shapiro-Wilk检验和Levene检验功能性状的群落加权平均值(CWM)方差的正态性和同质性。由于各变量均呈非正态分布, 且部分变量的方差不齐, 采用Kruskal-Wallis检验确定不同生境类型之间自生植物功能性状组成的差异性水平。选取P=0.05为具有显著性差异的阈值。在此基础上, 对16类功能性状在群落上的加权平均值(CWM)与物种多样性指数、功能多样性指数进行标准化处理, 采用spearman相关性分析检验8类生境中多样性指数与群落加权平均值(CWM)之间的关系。选取P=0.05为具有显著相关性的阈值。使用R中的“ggplot2”、“ggpubr”软件包进行统计图绘制。
2 研究结果 2.1 城市绿地中自生植物的种类和功能性状组成研究共调查到夏季自生植物176种, 隶属48科130属。生活型方面, 以一年生草本植物种类最多(68种, 不含草质藤本), 占总物种数的38.64%;其次为多年生草本植物(54种, 不含草质藤本), 藤本植物最少(14种)。物种来源方面, 本地植物112种, 占物种总数的63.64%;入侵植物21种, 占物种总数的11.93%。
共统计到130种自生植物能够通过昆虫进行授粉, 占总物种的73.86%, 其中有80种为自生本地植物;其次是风媒授粉型(49种)和自体授粉型(38种)自生植物;共有34种自生植物能够通过两种及以上的方式进行授粉。入侵植物中, 共统计到17种能够通过昆虫进行授粉。自体扩散(68种)是自生植物种子扩散的主要方式, 其次为动物扩散型(60种)、风扩散型(58种)和水扩散型(14种)。其中, 本地植物以自体扩散型为主(49种), 10种入侵植物通过风力进行扩散。
2.2 不同生境类型中自生植物的多样性特征各类生境所调查到的自生植物总物种数从多至少依次是落叶林地(127种)>常绿落叶林地(103种)>草地(88种)>灌丛(87种)>常绿林地(68种)>水边(63种)>行道树种植带(37种)>硬质铺装区域(20种)。Kruskal-Wallis检验结果显示, 群落水平自生植物的Patrick丰富度指数、Pielou均匀度指数在不同生境类型之间存在显著性差异(P<0.05)(图 1)。三种林地、灌丛和水边自生植物的Patrick丰富度指数显著高于硬质铺装区域。硬质铺装区域自生植物的Pielou均匀度指数显著高于落叶林地、草地、水边和行道树种植带。自生植物的Shannon-Wiener多样性指数与Simpson多样性指数在不同生境类型之间不存在显著差异(P>0.05), 两者的最高值均出现在常绿林地, 最低值均出现在硬质铺装区域。

|
| 图 1 不同生境下自生植物的物种多样性 Fig. 1 Species diversity of spontaneous plants between habitats 不同小写字母表示不同生境之间差异显著(P<0.05); ED:常绿落叶林地Evergreen and deciduous woodland;EW:常绿林地Evergreen woodland;DW:落叶林地Deciduous woodland;SH:灌丛Shrubland;GR:草地Grassland;WA:水边Waterside;TP:行道树种植带Tree pool;IP:硬质铺装区域Impervious pavement |
Kruskal-Wallis检验结果显示, 群落水平自生植物的功能丰富度指数、功能均匀度指数和Rao二次熵指数在生境水平存在显著差异(P<0.05), 在事后多重比较中无显著差异(P>0.05)(图 2)。自生植物的功能丰富度指数均值在行道树种植带生境较高, 在草地与硬质铺装区域较低。功能均匀度指数均值在硬质铺装区域、常绿落叶林地较高, 在水边最低。自生植物群落的Rao二次熵指数均值在常绿落叶林地最高, 在行道树种植带最低。自生植物的功能分歧度指数在不同生境类型之间均无显著差异(P>0.05), 最高均值出现在常绿落叶林地, 最低均值出现在行道树种植带。

|
| 图 2 不同生境下自生植物的功能多样性 Fig. 2 Functional diversity of spontaneous plants between habitats 不同小写字母表示不同生境之间差异显著(P<0.05) |
共有16项功能性状的群落加权平均值(CWM)在不同生境之间存在显著差异(P<0.05)(图 3)。草地中自生二年生草本的CWM值显著高于常绿落叶林地和落叶林地;常绿落叶林地和常绿林地中自生藤本植物的CWM值显著高于硬质铺装区域;草地和硬质铺装区域的自生乔、灌木植物CWM值显著低于林地和灌丛。物种来源方面, 行道树种植带和水边自生本地植物的CWM均值较高, 灌丛中自生本地植物的CWM均值最低;常绿落叶林地、常绿林地和落叶林地的自生国内外来植物CWM值显著高于草地、行道树种植带和硬质铺装区域;世界分布型自生植物的CWM最高均值出现在水边, 最低均值出现在灌丛。叶片性状方面, 硬质铺装区域自生植物的比叶面积CWM值显著低于行道树种植带。花部性状与果实性状方面, 三类林地中肉果型自生植物的CWM值显著高于草地和硬质铺装区域;行道树种植带中蓝紫色系开花植物的CWM值显著高于草地;草地自生植物群落始花期的CWM值显著高于水边、落叶林地和常绿落叶林地。种子性状方面, 草地中动物扩散型自生植物的CWM值显著高于落叶林地;三种林地和灌丛中自生植物种子干重显著高于草地。

|
| 图 3 不同生境下自生植物功能性状的群落加权平均值 Fig. 3 Community weighted mean of functional traits between habitats 不同小写字母表示不同生境之间差异显著(P<0.05) |
常绿林地中, 自生植物群落的Rao二次熵指数与国外外来植物CWM值(r=0.70, P<0.001)、种子干重CWM值(r=0.65, P<0.01)呈显著正相关, 与多年生草本植物CWM值(r=-0.73, P<0.001)、群落开花持续时间(r=-0.76, P<0.001)呈显著负相关。灌丛中, 自生植物群落的Simpson多样性指数与裂果型植物CWM值呈显著正相关(r=0.57, P<0.01), Pielou均匀度指数与风扩散型植物CWM值呈显著正相关(r=0.71, P<0.001), 功能分歧度指数与热带分布型植物CWM值(r=0.72, P<0.001)、C4光合途径型植物CWM值(r=0.65, P<0.01)呈显著正相关。草地中, 自生植物群落的Pielou均匀度指数与虫媒授粉型植物CWM值呈显著正相关(r=0.51, P<0.01), Rao二次熵指数与C4光合途径型植物CWM值呈显著正相关(r=0.60, P<0.001)。水边自生植物群落的Simpson多样性指数与一年生草本植物CWM值呈显著正相关(r=0.75, P<0.05), Rao二次熵指数与C4光合途径型植物CWM呈显著正相关(r=0.83, P<0.01)。行道树种植带中, 自生植物群落的Patrick丰富度指数与一年生草本植物CWM值(r=-0.71, P<0.05)、C4光合途径型植物CWM值(r=-0.73, P<0.05)呈显著负相关, 与温带分布型植物CWM值(r=0.84, P<0.01)、虫媒授粉型植物CWM值(r=0.83, P<0.01)呈显著正相关(图 4)。常绿落叶林地与落叶林地中自生植物群落的多样性水平与功能性状组成之间仅存在部分显著弱相关(-0.50<r<0.50)。

|
| 图 4 不同生境中自生植物多样性与功能性状组成的关系 Fig. 4 Relationship between spontaneous plant diversity and functional trait composition in habitats r为spearman相关系数;选取P<0.05;阴影区域表示95%的置信区间 |
城市绿地支撑了丰富的本地自生植物种类, 极大地丰富了城市植物的多样性[10]。但由于频繁受到养护管理的干扰, 绿地中的自生植物呈现出一年生植物为主的群落构成特征。相比于以往对北京地区城市河流廊道和单一城市公园的研究[32—33], 本研究调查到更为丰富的自生乔、灌木植物, 可能是绿地中丰富的栽培木本种类为自生木本苗提供种子来源。但研究也发现, 丰富的生境与频繁人类活动在一定程度上也为国内、国外外来植物与入侵植物的定植、生长提供了机会。
超过70%的自生植物能够通过昆虫进行授粉, 证实了城市绿地中自生植物在支持授粉昆虫多样性中发挥的重要作用。调查到的以昆虫为授粉媒介的自生植物中, 本地植物占到了一半以上, 这一结论证实了自生本地植物作为重要蜜粉来源, 在城市绿地支持传粉昆虫多样性方面做出的贡献[34—35]。绿地中调查到的自生植物中, 接近40%的物种通过种子自身重力或产生的机械力量进行扩散, 略多于借助风力扩散的植物, 这可能由于绿地中复杂的微地形与植物群落在一定程度上阻挡了空气的流通, 阻碍了风力扩散型自生植物的扩散[17]。值得注意的是, 超过40%的自生入侵植物借助风力进行种子扩散, 这类入侵植物可以在绿地中进行远距离的扩散, 进而增加对非入侵植物的威胁[36]。
3.2 不同生境类型中自生植物多样性与功能性状组成的差异本文的结果验证了假设——城市绿地中丰富的生境类型为自生植物种类和功能性状的多样化提供了可能, 不同生境类型中自生植物的多样性和功能性状组成存在差异。并且相比于物种多样性, 不同生境类型中自生植物的功能性状组成差异更为显著。与前人研究结果一致, 林地和水边往往支持了丰富多样的自生植物种类, 而草地和硬质铺装区域包含的物种数量较低[1, 37]。本文研究还发现, 具有较高物种丰富度水平的林地和水边生境, 同样支持了丰富的自生植物功能性状;而草地和硬质铺装区域自生植物的功能丰富度水平较低;相比于种植了落叶树种的林地, 常绿林地中自生植物功能性状的离散程度更低。值得注意的是, 行道树种植带分布的少量自生植物种类覆盖了丰富的功能性状。
大量研究证实, 环境过滤显著影响了植物定植的可能性, 从而影响环境中植物的多样性, 这种过滤作用能够通过群落的功能性状组成得到直观体现[5]。本文结果表明, 林地和水边支持了更丰富的自生多年生的草本、藤本和乔、灌木植物, 如大车前(Plantago major)、萝藦(Cynanchum rostellatum)等, 这些植物可以多年稳定生长, 为形成物种与功能丰富的群落提供可能。相比之下, 草地往往受到较高的人为干扰, 这使得对环境敏感的多年生植物难以生存下来而一、二年生植物占比高, 如荠(Capsella bursa-pastoris)、牛筋草(Eleusine indica)等, 这与前人研究结果相似[38]。这些植物生长速度快、竞争力强, 易在群落中形成优势种进而影响群落物种的丰富水平[39]。硬质铺装区域高干扰、高温、低湿、低养分的环境形成了严格的过滤器, 导致其中分布的自生植物物种丰富度低且功能趋于同质化, 以生活周期短、种子重量轻、比叶面积小的植物为主, 如马齿苋(Portulaca oleracea)、朝天委陵菜(Potentilla supina)等, 这些植物耐高温、干旱且对养分利用率高, 能够适应高温、干旱、资源贫瘠的硬质铺装区域[40—41]。在行道树种植带调查到了丰富的自生本地植物, 如地黄(Rehmannia glutinosa)、酢浆草(Oxalis corniculata)等。这些本地植物被证实具有更强的生态弹性与抗干扰能力, 能够在频繁受到人类活动影响的行道树种植带中生存下来[42]。值得注意的是, 尽管行道树种植带分布的自生植物种类不及林地和水边, 但这些少量的本地植物覆盖了丰富的功能性状, 这一结果证实了本地植物在功能多样性保护与生态功能发挥中的重要地位[43—44]。除了受到环境的过滤, 自生植物多样性还受到绿地的土壤、自然残留植被及栽培植物等潜在种子库的影响, 这些往往与绿地的土地利用历史、建成年限、建设理念等相关[45—46]。在林地和灌丛中调查到了大量的自生乔、灌木植物, 如栾树(Koelreuteria paniculata)、紫丁香(Syringa oblata)等,这类生境中人工栽植的乔木与灌木为地被层自生木本苗提供了重要来源, 极大丰富了自生植物物种与功能性状。
3.3 群落中相对稀有的功能性状在自生植物多样性形成与维持中的重要性本研究进一步探索了特定生境类型中, 即环境条件相对一致的情况下, 不同自生植物群落之间多样性与功能性状组成之间的关系, 以更好地理解特定生境中自生植物多样性的形成过程与机制。研究结果表明, 一些在群落水平相对稀有的功能性状与多样性之间存在显著相关性。根据环境过滤理论, 环境作为一个“过滤器”只允许具有特定特征的物种在其中定居生长[47]。大量研究证实了这种机制在城市化进程中产生的负面影响, 城市环境对当地物种过滤而表现出物种和功能性状同质化, 这意味着区域内优势物种可能存在功能冗余的现象[48—49]。而那些相对稀有的物种往往具有独特的功能性状, 对城市生物多样性保护和提升同样具有重要贡献[50—51]。在多样性水平较低的草地生境, 尽管C4光合途径型植物(如马齿苋、地锦草(Euphorbia humifusa))在群落中的占比明显低于C3光合途径型植物, 但随着群落中C4光合途径型植物的增加群落物种多样性呈现升高趋势。灌丛中自生植物的物种和功能多样性均随着少量分布的裂果型植物(如繁缕(Stellaria media)、少花米口袋(Gueldenstaedtia verna))和C4光合途径型植物的增加而得到提高。水边的自生植物群落中, 少量分布的一年生草本植物的增加往往意味着群落多样性水平的提高, 如萹蓄(Polygonum aviculare)、酸模叶蓼等(Persicaria lapathifolia)。灌丛中的风扩散型植物和草地中的虫媒授粉型植物能够提高群落的物种均匀度, 在一定程度上避免了单一优势种的聚集。
常绿林地中自生植物功能多样性与国外外来植物占比存在正相关趋势, 即自生植物多样性水平的增加可能是由这些占比较小的非本地植物引起的。这些由人为无意或有意引入的非本地植物往往具有与本地物种不同的功能性状, 能够提高群落对环境资源的利用效率[52]。此外, 特定生境中占比较小的地理成分同样能够提供独特的种类与功能, 如灌丛中的热带分布型植物和行道树种植池的温带分布型植物, 对自生植物群落多样性水平的提升具有贡献。
3.4 在生境类型丰富的城市绿地中保护和利用自生植物的策略自生植物经过了环境的筛选与过滤, 在绿地中自播繁衍、顽强生长, 不需要过多养护管理而减少了养护成本和资源消耗。同时, 具有不同功能性状的植物自发形成群落, 按照各自的节律完成生活史, 随生境和季节动态变化, 在绿地中形成了近自然、可持续的植物景观[33, 53]。水边自生植物群落中分布有更多的蓝紫色系开花植物, 如鸭跖草(Commelina communis)、乳苣(Lactuca tatarica), 而黄色系开花植物在常绿林地中的占比最大, 如酢浆草、尖裂假还阳参(Crepidiastrum sonchifolium)等。此外, 各类生境中分布了许多三至四月开花的植物, 如蒲公英(Taraxacum mongolicum)、早开堇菜(Viola prionantha)等, 能够丰富绿地的早春植物景观。此外, 研究结果显示, 具有较高多样性水平的自生植物群落, 往往具有更多的虫媒授粉型植物, 如旋覆花(Inula japonica)、长萼鸡眼草(Kummerowia stipulacea)。这类植物规模的增加能够吸引更多的授粉媒介, 进而提高植物群落与传粉昆虫之间的互动[54], 这大大提高了绿地对传粉昆虫的支持功能从而改善生物多样性水平。这些丰富多样的自生植物能够有效提升绿地的景观效果与生态服务功能, 因此, 将自生植物群落纳入景观规划与设计的范畴是十分必要的。
然而, 一味追求高水平的植物多样性是不可取的。研究结果显示, 多样性更高的群落在花期和果期的持续时间上可能表现出更低的趋势。这可能是环境过滤强于生物过滤的结果, 前者通过环境因子筛选出相似的功能性状, 使得群落中的物种具有相似的开花物候;后者则筛选出开花物候相异的物种, 避免对授粉媒介的竞争[55—56]。这种过滤机制有可能造成一些功能冗余或具有入侵性的物种在群落中聚集, 如反枝苋(Amaranthus retroflexus)、小蓬草(Erigeron canadensis)。这些植物虽然能在短时间内提高群落的物种多样性水平, 但它们在性状空间分布上往往高度重合[57], 增加同类资源竞争强度的同时降低对环境资源的利用效率, 对群落的可持续发展和生态功能发挥造成威胁。因此, 在城市绿地自生植物的管理实践中, 要注意对这类植物的干预, 尤其是在落叶林地、草地等容易滋生入侵植物的生境中。
自生植物的多样性特征及其形成与维持机制因生境类型而异, 这意味着城市绿地自生植物的生境营建与管理策略需要细化到特定的生境类型。研究结果表明, 环境中分布的相对稀有的物种与功能性状可能是提升植物多样性的关键要素, 且这些要素因特定生境类型而存在差异。在城市绿地营建与管理中, 需要加强对灌丛、草地和水边C4光合途径型自生植物的评估与保护, 如马齿苋、朝天委陵菜、白鳞莎草(Cyperus nipponicus), 因为这些植物占比的增加可能带来自生植物多样性水平的提升。适当保留水边非入侵性的自生一年生草本植物, 如萹蓄、长芒稗(Echinochloa caudata)、鸭跖草, 对提升群落功能多样性具有积极作用;但在行道树种植带, 适度控制群落中自生一年生草本植物的生长更利于形成物种多样的自生植物群落。此外, 城市绿地中的栽培植物景观引入了大量外来植物, 为城市绿地自生植物提供了潜在种子库。本研究结果表明, 这类外来植物易在林地、灌丛中自发生长从而影响自生植物群落的多样性特征。常绿林地中自生外来植物的增加能够给自生植物群落带来丰富的功能性状, 但灌丛中自生外来植物对群落的多样性水平可能带来积极或消极的影响。后续研究需要加强对自生外来植物的持续调查、监测与评估。
4 结论目前, 我国城市绿地建设正在全力推进, 这些大大小小的绿地有效缓解了自然栖息地与原生植被退化对城市生物多样性与景观风貌造成的威胁。研究结果显示, 在20个总面积为190hm2的城市绿地中支持的夏季自生植物物种数可达176种, 其中包含的本地植物和虫媒型开花植物资源十分可观。作为绿地的主要生境类型, 三种林地均支持了丰富的自生植物物种和功能性状;草地和硬质铺装区域自生植物群落的物种与功能性状多样性水平低且呈现均质化。结果表明, 林地分布有更多的自生乔、灌木植物与外来植物;草地中则以自生一年生植物与本地植物为主。环境中分布相对稀有的自生植物物种和功能性状与多样性之间存在显著相关性, 对自生植物多样性的提升具有重要意义。高光效与高养分利用率的C4植物对草地、灌丛和水边自生植物多样性提升具有积极作用;外来植物能够为林地中自生植物群落提供独特的功能性状。这些结果从功能性状组成角度解释了不同生境中自生植物群落的成分差异, 更好地理解特定生境中自生植物多样性形成和维持过程。在未来的城市绿地植物景观营建与管理中, 需要针对特定的生境类型, 采取科学、合理的植物群落设计策略和管理措施, 以优化近自然、可持续的植物景观营造与生物多样性保护、提升的实施路径。
| [1] |
李晓鹏, 董丽, 关军洪, 赵凡, 吴思佳. 北京城市公园环境下自生植物物种组成及多样性时空特征. 生态学报, 2018, 38(2): 581-594. |
| [2] |
Müller A, Bøcher P K, Fischer C, Svenning J C. 'Wild' in the city context: do relative wild areas offer opportunities for urban biodiversity?. Landscape and Urban Planning, 2018, 170: 256-265. DOI:10.1016/j.landurbplan.2017.09.027 |
| [3] |
侯冰飞, 贾宝全, 冷平生, 王文和. 北京市城乡交错区绿地和植物种类的构成与分布. 生态学报, 2016, 36(19): 6256-6265. |
| [4] |
Zhu Z X, Roeder M, Xie J, Nizamani M M, Friedman C R, Wang H F. Plant taxonomic richness and phylogenetic diversity across different cities in China. Urban Forestry & Urban Greening, 2019, 39: 55-66. |
| [5] |
Zhang M Y, Fan S X, Li X L, Li K, Xing X Y, Hao P Y, Dong L. How urban riparian corridors affect the diversity of spontaneous herbaceous plants as pollination and dispersal routes——a case of the Wenyu River- North Canal in Beijing, China. Ecological Indicators, 2023, 146: 109869. DOI:10.1016/j.ecolind.2023.109869 |
| [6] |
徐幼榕, 张梦园, 范舒欣, 李晓璐, 董丽, 曾赞青. 城市社区公园自生草本生态位及群落特征——以北京市西城区为例. 安徽农业大学学报, 2023, 50(5): 784-791. |
| [7] |
Li C S, Liang H Y, Gao D Y, Wang Y B, Jin K D, Liu J N, Xue D G, Chen Y Y, Li Y P, Gao T, Qiu L. Comparative study on the effects of soil quality improvement between urban spontaneous groundcover and lawn. Ecological Indicators, 2023, 148: 110056. DOI:10.1016/j.ecolind.2023.110056 |
| [8] |
Villaseñor N R, Chiang L A, Hernández H J, Escobar M A H. Vacant lands as refuges for native birds: an opportunity for biodiversity conservation in cities. Urban Forestry & Urban Greening, 2020, 49: 126632. |
| [9] |
Yin L T, Zheng W, Shi H H, Ding D W. Ecosystem services assessment and sensitivity analysis based on ANN model and spatial data: a case study in Miaodao Archipelago. Ecological Indicators, 2022, 135: 108511. DOI:10.1016/j.ecolind.2021.108511 |
| [10] |
Chang C R, Chen M C, Su M H. Natural versus human drivers of plant diversity in urban parks and the anthropogenic species-area hypotheses. Landscape and Urban Planning, 2021, 208: 104023. DOI:10.1016/j.landurbplan.2020.104023 |
| [11] |
Kümmerling M, Müller N. The relationship between landscape design style and the conservation value of parks: a case study of a historical park in Weimar, Germany. Landscape and Urban Planning, 2012, 107(2): 111-117. DOI:10.1016/j.landurbplan.2012.05.006 |
| [12] |
Cervelli E W, Lundholm J T, Du X. Spontaneous urban vegetation and habitat heterogeneity in Xi'an, China. Landscape and Urban Planning, 2013, 120: 25-33. DOI:10.1016/j.landurbplan.2013.08.001 |
| [13] |
陈春谛. 被遗忘的城市"生境": 重庆市墙体自生植物调查分析. 生态学报, 2020, 40(2): 473-483. |
| [14] |
Zhang H L, Muhammad Nizamani M, Padullés Cubino J, Harris A J, Guo L Y, Zhou J J, Wang H F. Habitat heterogeneity explains cultivated and spontaneous plant richness in Haikou City, China. Ecological Indicators, 2023, 154: 110713. DOI:10.1016/j.ecolind.2023.110713 |
| [15] |
Li X P, Fan S X, Guan J H, Zhao F, Dong L. Diversity and influencing factors on spontaneous plant distribution in Beijing Olympic Forest Park. Landscape and Urban Planning, 2019, 181: 157-168. DOI:10.1016/j.landurbplan.2018.09.018 |
| [16] |
李晓鹏, 张思凝, 冯黎, 吴然. 成都城区河流廊道自生植物的生境及物种多样性. 风景园林, 2022, 29(1): 64-70. |
| [17] |
Marini L, Bruun H H, Heikkinen R K, Helm A, Honnay O, Krauss J, Kühn I, Lindborg R, Pärtel M, Bommarco R. Traits related to species persistence and dispersal explain changes in plant communities subjected to habitat loss. Diversity and Distributions, 2012, 18(9): 898-908. DOI:10.1111/j.1472-4642.2012.00893.x |
| [18] |
Palma E, Catford J A, Corlett R T, Duncan R P, Hahs A K, McCarthy M A, McDonnell M J, Thompson K, Williams N S G, Vesk P A. Functional trait changes in the floras of 11 cities across the globe in response to urbanization. Ecography, 2017, 40(7): 875-886. DOI:10.1111/ecog.02516 |
| [19] |
吴雪, 黄力, 靳程, 钱深华, 杨永川. 重庆主城区杂草物种组成特征及多样性格局. 华东师范大学学报: 自然科学版, 2020, 210(2): 98-109. |
| [20] |
E-Vojtkó A, de Bello F, Lososová Z, Götzenberger L. Phylogenetic diversity is a weak proxy for functional diversity but they are complementary in explaining community assembly patterns in temperate vegetation. Journal of Ecology, 2023, 111(10): 2218-2230. DOI:10.1111/1365-2745.14171 |
| [21] |
韩会然, 杨成凤, 宋金平. 北京市土地利用空间格局演化模拟及预测. 地理科学进展, 2015, 34(8): 976-986. |
| [22] |
贺士元, 邢其华, 尹祖棠. 北京植物志 (2版). 北京: 北京出版社, 1993.
|
| [23] |
肖翠, 刘冰, 吴超然, 马金双, 叶建飞, 夏晓飞, 林秦文. 北京维管植物编目和分布数据集. 生物多样性, 2022, 30(6): 5-13. |
| [24] |
崔夏, 刘全儒, 吴超然, 何宇飞, 马金双. 京津冀外来入侵植物. 生物多样性, 2022, 30(8): 151-160. |
| [25] |
吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1991, 13(增4): 1-139. |
| [26] |
翟增康, 赵珉, 魏永胜, 刘虎岐, 白瑶. 中国C4植物名录. 陕西林业科技, 2020, 48(5): 71-89, 93. |
| [27] |
黎维平. 果实分类系统的修订和教学——植物学教材质疑(十). 生命科学研究, 2022, 26(5): 460-470. |
| [28] |
Kattge J, Bönisch G, Díaz S, Lavorel S, Prentice I C, …, Wirth C (729 authors). Try plant trait database-enhanced coverage and open access. Global Change Biology, 2020, 26. |
| [29] |
Society for Ecological Restoration, International Network for Seed Based Restoration and Royal Botanic Gardens Kew. Seed Information Database. [2024-01-23]. https://ser-sid.org/.
|
| [30] |
张金屯. 数量生态学 (3版). 北京: 科学出版社, 2018.
|
| [31] |
Laliberté E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305. |
| [32] |
张梦园, 李坤, 邢小艺, 范舒欣, 徐一丁, 郝培尧, 董丽. 北京温榆河-北运河生态廊道自生植物多样性对城市化的响应. 生态学报, 2022, 42(7): 2582-2592. |
| [33] |
Li X P, Fan S X, Hao P Y, Dong L. Temporal variations of spontaneous plants colonizing in different type of planted vegetation-a case of Beijing Olympic Forest Park. Urban Forestry & Urban Greening, 2019, 46: 126459. |
| [34] |
Martínez-Núñez C, Kleijn D, Ganuza C, Heupink D, Raemakers I, Vertommen W, Fijen T P M. Temporal and spatial heterogeneity of semi-natural habitat, but not crop diversity, is correlated with landscape pollinator richness. Journal of Applied Ecology, 2022, 59(5): 1258-1267. |
| [35] |
Balfour N J, Ratnieks F L W. The disproportionate value of 'weeds' to pollinators and biodiversity. Journal of Applied Ecology, 2022, 59(5): 1209-1218. |
| [36] |
诸葛晓龙, 朱敏, 季璐, 崔旭红. 入侵杂草小飞蓬和钻形紫菀种子风传扩散生物学特性研究. 农业环境科学学报, 2011, 30(10): 1978-1984. |
| [37] |
陈晓双, 梁红, 宋坤, 达良俊. 哈尔滨中心城区杂草物种多样性及其在异质生境中的分布特征. 生态学杂志, 2014, 33(4): 946-952. |
| [38] |
Li X P, Li Y L, Zhang S N, Lin R, Chen M K, Feng L. Driving effects of land use and landscape pattern on different spontaneous plant life forms along urban river corridors in a fast-growing city. Science of the Total Environment, 2023, 876: 162775. |
| [39] |
李建, 卢世君, 邱礼杉, 庄春晓, 杨芳芳, 赵娟娟. 深圳市建成区入侵植物对草本植物种类及功能多样性的影响. 生态学报, 2023, 43(18): 7507-7522. |
| [40] |
Havrilla C A, Bradford J B, Yackulic C B, Munson S M. Divergent climate impacts on C3 versus C4 grasses imply widespread 21st century shifts in grassland functional composition. Diversity and Distributions, 2023, 29(3): 379-394. |
| [41] |
Hu L Y, Qin D Y, Lu H Y, Li W, Shang K K, Lin D M, Zhao L, Yang Y C, Qian S H. Urban growth drives trait composition of urban spontaneous plant communities in a mountainous city in China. Journal of Environmental Management, 2021, 293: 112869. |
| [42] |
Celesti-Grapow L, Pyšek P, Jarošík V, Blasi C. Determinants of native and alien species richness in the urban flora of Rome. Diversity and Distributions, 2006, 12(5): 490-501. |
| [43] |
Wang C Y, Yu Y L, Cheng H Y, Du D L. Which factor contributes most to the invasion resistance of native plant communities under the co-invasion of two invasive plant species?. Science of the Total Environment, 2022, 813: 152628. |
| [44] |
Sun X, Iwachido Y, Hayashi S, Horiuchi S, Kirishita M, Mori A S, Sasaki T. Plant functional rarity across different land use types in the megacity of Tokyo. Urban Forestry & Urban Greening, 2022, 78: 127779. |
| [45] |
王月, 赵亮. 重庆市不同年龄综合公园绿化乔木树种多样性和组成的比较研究. 广西植物, 2020, 40(4): 546-556. |
| [46] |
Li X P, Li X L, Zhang M Y, Luo Q Y, Li Y L, Dong L. Urban park attributes as predictors for the diversity and composition of spontaneous plants: A case in Beijing, China. Urban Forestry & Urban Greening, 2024, 91: 128185. |
| [47] |
Kraft N J B, Adler P B, Godoy O, James E C, Fuller S, Levine J M. Community assembly, coexistence and the environmental filtering metaphor. Functional Ecology, 2015, 29(5): 592-599. |
| [48] |
Kondratyeva A, Knapp S, Durka W, Kühn I, Vallet J, Machon N, Martin G, Motard E, Grandcolas P, Pavoine S. Urbanization effects on biodiversity revealed by a two-scale analysis of species functional uniqueness vs. Redundancy. Frontiers in Ecology and Evolution, 2020, 8: 73. |
| [49] |
Bruno D, Gutiérrez-Cánovas C, Sánchez-Fernández D, Velasco J, Nilsson C. Impacts of environmental filters on functional redundancy in riparian vegetation. Journal of Applied Ecology, 2016, 53(3): 846-855. |
| [50] |
Mouillot D, Bellwood D R, Baraloto C, Chave J, Galzin R, Harmelin-Vivien M, Kulbicki M, Lavergne S, Lavorel S, Mouquet N, Paine C E T, Renaud J, Thuiller W. Rare species support vulnerable functions in high-diversity ecosystems. PLoS Biology, 2013, 11(5): e1001569. |
| [51] |
Enquist B J, Feng X, Boyle B, Maitner B, Newman E A, Jørgensen P M, Roehrdanz P R, Thiers B M, Burger J R, Corlett R T, Couvreur T L P, Dauby G, Donoghue J C, Foden W, Lovett J C, Marquet P A, Merow C, Midgley G, Morueta-Holme N, Neves D M, Oliveira-Filho A T, Kraft N J B, Park D S, Peet R K, Pillet M, Serra-Diaz J M, Sandel B, Schildhauer M, Šímová I, Violle C, Wieringa J J, Wiser S K, Hannah L, Svenning J C, McGill B J. The commonness of rarity: global and future distribution of rarity across land plants. Science Advances, 2019, 5(11): eaaz0414. |
| [52] |
Bonthoux S, Voisin L, Bouché-Pillon S, Chollet S. More than weeds: spontaneous vegetation in streets as a neglected element of urban biodiversity. Landscape and Urban Planning, 2019, 185: 163-172. |
| [53] |
Brun M, Di Pietro F, Bonthoux S. Residents' perceptions and valuations of urban wastelands are influenced by vegetation structure. Urban Forestry & Urban Greening, 2018, 29: 393-403. |
| [54] |
E-Vojtkó A, de Bello F, Durka W, Kühn I, Götzenberger L. The neglected importance of floral traits in trait-based plant community assembly. Journal of Vegetation Science, 2020, 31(4): 529-539. |
| [55] |
Fantinato E, Del Vecchio S, Giovanetti M, Acosta A T R, Buffa G. New insights into plants co-existence in species-rich communities: the pollination interaction perspective. Journal of Vegetation Science, 2018, 29(1): 6-14. |
| [56] |
Camargo M G G, Arista M, Lunau K, Ortiz P L, Le Stradic S, Rocha N M W B, Morellato L P C. Flowering phenology of species with similar flower colours in species-rich communities. Flora, 2023, 304: 152312. |
| [57] |
Mologni F, Bellingham P J, Cameron E K, Dinh K, Wright A E, Burns K C. Functional traits explain non-native plant species richness and occupancy on northern New Zealand Islands. Biological Invasions, 2022, 24(7): 2135-2154. |