 2024, Vol. 44
2024, Vol. 44文章信息
- 於嘉禾, 王卫光, 陈泽峰
- YU Jiahe, WANG Weiguang, CHEN Zefeng
- 全球旱地饱和水汽压差和根区土壤水分变化对植被生产力的影响及其成因
- Influences of vapor pressure deficit and root-zone soil moisture changes on vegetation productivity and its causes across global drylands
- 生态学报. 2024, 44(11): 4808-4819
- Acta Ecologica Sinica. 2024, 44(11): 4808-4819
- http://dx.doi.org/10.20103/j.stxb.202309262093
-
文章历史
- 收稿日期: 2023-09-26
- 网络出版日期: 2024-03-18






2. 长江保护与绿色发展研究院, 南京 210098;
3. 河海大学水文水资源学院, 南京 210098
2. Yangtze Institute for Conservation and Development, Nanjing 210098, China;
3. College of Hydrology and Water Resources, Hohai University, Nanjing 210098, China
旱地生态系统在全球生态系统功能中发挥着举足轻重的作用。近期一项研究表明, 旱地植被主导着全球植被生产力的增加趋势[1], 然而旱地生态系统是对气候变化最为敏感的地区之一[2], 目前面临退化的风险[3]。因此在全球气候变化背景下, 外部因素对旱地植被生长的影响亟需得到量化和分析。
得益于气候变暖及二氧化碳施肥效应对植被生长季和最大光合速率的提升, 全球旱地的植被变绿已经得到了广泛证实[4]。然而气候变化也对生态系统产生了威胁, 这主要源于大气水分亏缺量的上升和土壤含水率下降以及这两个事件引发的复合效应。一方面, 土壤水是控制生态系统水碳通量的关键非生物因素[5], 土壤水可以调节植物根系的水分吸收和气孔导度以决定植物的水分状态[6]。然而, 干旱胁迫提升了木质部的水分张力, 诱发植物栓塞, 从而引起植被广泛的水力破坏[7]。另一方面, 由于气候变暖大气饱和水汽压差(VPD)不断上升, 较高的VPD促使植物倾向于降低气孔导度以便在给定的碳增益下实现水分损失的最小化[8], 这限制了植物的实际光合速率, 引发了植被生态系统碳储量的负平衡[9]。与此同时, 在低土壤湿度条件下, 潜热通量的减少和感热通量的上升往往会诱发低土壤湿度和高VPD的复合事件, 这强烈限制了生态系统的碳吸收过程, 并可能引发植被死亡[10]。
尽管先前的研究已经基于多种数据统计和模型模拟方法捕捉了植被生长对土壤水分敏感性的上升以及VPD上升对植被生长的负效应[11—12], 然而土壤水分和VPD对全球旱地区域不同种类植被生长的影响以及它们与当地气候条件的联系仍未得到较为全面的认识, 且由于植被生产力对气候变化的响应机制较为复杂, 局部尺度的研究很容易忽略由于环境要素、植被覆盖等因素的差异对研究结果的影响。为了解决上述问题, 本研究利用多年降水和潜在蒸散发数据划分了全球旱地区域, 并基于1982至2014年的多源气象、根区土壤含水率和植被总初级生产力(GPP)数据, 参考两种不同的植被生长季定义将所有变量汇总至生长季尺度, 基于仿真实验思想和随机森林算法量化了植被生产力对根区土壤含水率以及饱和水汽压差的敏感性, 分析了不同类型植被对根区土壤含水率和饱和水汽压差的敏感性特征, 并结合分档平均方法探究了敏感性与当地气候条件的潜在关系。
1 材料与方法 1.1 研究区划分本文依据多年尺度的干旱指数(AI)划分全球旱地区域, 该指数被定义为年降水量与年潜在蒸散发量多年平均值的比值[13]。其中, 干旱指数介于0.05至0.65的区域被定义为旱地区域, 其中:干旱指数小于至0.20的地区为干旱区;干旱指数介于0.2至0.5的地区为半干旱区, 干旱指数介于0.5至0.65的地区为半湿润区[14]。本研究基于CRU TS V4.06数据集的月尺度降水和潜在蒸散发数据计算得出干旱指数的全球分布, 并划分出旱地区域, 结果如下图所示:

|
| 图 1 全球干旱指数(AI)分布及旱地区域 Fig. 1 Distribution of global aridity index (AI) and drylands |
GPP产品是在对新型遥感植被指数近红外反射率指数(Nirv)作为日光诱导叶绿素荧光(SIF)和GPP探针的可行性论证基础上, 基于先进高分辨率辐射计(AVHRR)遥测数据集和全球数百个通量站数据生成。该产品时间分辨率为月尺度, 空间分辨率为0.05°×0.05°, 相较于其他GPP产品, 该产品更依赖于实测而非模型模拟, 且以较高精度实现了对全球GPP的估计, 其季节变化也与通量站点和模式的结果一致, 因此该产品可用于陆地碳通量和未来气候的预测[15]。此外本文选用了植被物候遥感产品VIP, 该产品的生成基于logistic模型的地表物候(LSP)检测算法, 结合了AVHRR LTDR和MODIS CMG记录生成的逐日植被增强指数(EVI)数据[16], 提供了1981—2016年全球植被生长季节开始和结束的日期。
1.2.2 气象数据集本文选取的遥感气象数据来源于ERA5-Land再分析数据集, 该数据集由欧洲中期天气预报中心(ECMWF)开发。该数据集的集成基于ECMWF全球高分辨率陆地表面数值模型, 涵盖了1950年至今的许多陆面要素, 以一致的方式描述了陆地水和能量循环的演变过程。与ERA5和ERA-Interim数据集相比, 该数据集可以在高时间分辨率(小时)下提供更高空间分辨率的数据, 尤其是提升了土壤水分、湖泊等水文变量的精度[17]。本文选取了该数据集中的月尺度气温(T, ℃)、露点温度(Td, ℃)、下行短波辐射(Srad, J/m2)及ERA5数据集中的近地表 10m风速数据(WS, m/s), 具体如下表所示:
| 变量名称 Name of variables |
数据集名称 Name of datasets |
范围与分辨率 Range and resolution |
时间范围 Time span |
数据来源 Data sources |
| P, PET | CRU TS V4.06 | 全球, 0.5°×0.5°, 月尺度 | 1901—2020年 | https://www.uea.ac.uk/ |
| T, Td, Srad, WS | ERA5-Land, ERA5 | 全球, 0.1°×0.1°, 月尺度 | 1950年至今 | https://cds.climate.copernicus.eu/ |
| CO2 | Mauna Loa站点数据 | 站点尺度, 月尺度 | 1958年3月至今 | https://gml.noaa.gov/ |
| P:降水量Precipitation;PET:潜在蒸散发量Potential Evapotranspiration;T:气温Air Temperature;Td:露点温度Dew point temperature;Srad:入射太阳辐射Incoming Solar Radiation;WS:风速Wind Speed | ||||
由于ERA5-Land数据集未提供月尺度VPD数据, 因此本文采用联合国粮食及农业组织(FAO)所通用的VPD计算方法, 基于ERA5-Land数据集的月尺度气温和露点温度数据计算出研究区每个格点的VPD, 计算公式如下:
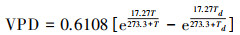
|
(1) |
式中:T表示月尺度气温;Td表示月尺度露点温度, VPD表示月尺度饱和水汽压差。
1.2.3 根区土壤含水率数据集本文选用全球地表蒸散发阿姆斯特丹模型产品(GLEAM V3.6a)的根区土壤含水率数据。该数据集空间分辨率为0.25°×0.25°, 单位为体积含水率(m3/m3), 该数据集结合多源降水、遥感观测地表土壤水分等数据, 基于水量平衡原理估计地表和根区含水量[18], 其根区土壤湿度估算值与全球多个土壤水分观测值集合实现了较高的精度验证结果[19]。
1.2.4 地表覆盖数据GLASS-GLC数据集以1982—2015年的全球陆表特征参量数据集(GLASS CDR)为数据源, 基于有限样本稳定分类理论, 它有效确保了数据产品的可靠性以及地物判别的有效性。经全球独立样本库检验, 该产品的年平均总体精度为82.81%, 并且首次以5公里空间分辨率记录了1982年至2015年全球土地覆盖的时空变化格局。与现有土地覆盖数据产品相比, GLASS-GLC数据集具有高精度、高一致性、高可比性的特点[20]。该数据集将地表覆盖分为耕地、森林、草地、灌丛、苔原、荒地、冰雪七大类别。本研究以1982年的土地覆盖分类结果作为植被的分类依据, 具体植被空间分布及其在全球旱地面积占比如图 2所示:

|
| 图 2 全球旱地植被类型空间分布及面积占比 Fig. 2 Spatial distribution and area proportion of vegetation types in global drylands |
本研究中所有原始数据被统一重采样至0.5°×0.5°空间分辨率(对于植被物候数据、土地覆盖数据采用最近邻插值算法、其余数据采用双线性插值算法), 时间分辨率统一为月尺度。由于陆地生态系统的生长季是影响全球植物生产力的主要决定因素之一, 且植被生长季节的定义具有多样性, 因此本文采用了两种常用的生长季定义:(1)基于物候数据集VIP的生长季开始和结束日期[22]。(2)基于研究时期月均温多年均值高于零摄氏度的月份[21]。因此可以依据物候数据集VIP、ERA5-Land的温度数据将所有变量汇总为两组数据集, 以ERA5-Land(VIP), ERA5-Land(TMP)命名。对于第一种方式定义的植被生长季, 为了在每个栅格上得到一个可以保留与物候周期年际变化相关的且固定的植被生长季, 本研究参考了Chen等[22]在过去研究中采用的方法:首先在重采样后的物候数据网格单元上提取多年平均的生长季节开始日期和结束日期;随后, 根据多年平均生长季开始日期和结束日期确定每个植被网格单元的生长季所包含的月份, 并凭此判断每个格点在某一月份是否处于生长季;最后, 对GPP、Srad数据采取生长季各月份求和的方式, 而对其余变量采取求平均的方式。
1.3.2 趋势分析与趋势估计Mann-Kendall趋势检验法是一种可用于判断趋势显著性的非参数统计检验方法, 且当Mann-Kendall趋势检验统计量超过1.96, 则表示该序列通过了95%置信水平下的显著性检验(P < 0.05)。为了量化饱和水汽压差、根区土壤含水率以及植被生产力的变化速率, 本文采用Theil-Sen趋势估计法对植被生产力、根区土壤含水率以及饱和水汽压差进行趋势估计。这两种方法的共同优势在于:不要求样本遵循一定的分布, 且不受少数异常值的干扰[23]。
1.3.3 模拟实验设计与敏感性计算鉴于生态系统各要素之间往往呈现非线性耦合关系, 且机器学习算法在解决复杂非线性问题具备较强优势, 并已经在植被生长对土壤水分的敏感性的研究中得到了应用[24], 因此本文根据上述数据, 将植被生产力设置为因变量, 其余因子设置为自变量, 基于随机森林算法构建黑箱模型:
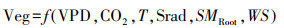
|
(2) |
式中:Veg代表生长季尺度植被生产力; CO2, VPD, T, WS分别表示生长季二氧化碳浓度均值(‰)、饱和水汽压差均值(kPa)、气温和10m风速(m/s)均值, Srad表征生长季下行短波辐射通量总和(J/m2), SMRoot表示生长季根区土壤水分均值, 三组模拟实验用于分离生长季根区土壤水分、VPD在整个研究时期对植被生产力的影响, 具体如表 2所示:
| 实验名称 Name of experiment |
情境描述 Description of situation |
| 仿真实验(S0) Simulation experiment | 所有因子均为时变因子 |
| 控制实验1(S1) Control experiment 1 | VPD为时不变因子, 其余因子均保持时变 |
| 控制实验2(S2) Control experiment 2 | 根区土壤水分为时不变因子, 其余因子均保持时变 |
| VPD:饱和水汽压差Vapor pressure deficit | |
在随机森林算法中, 所有模拟实验均设置了50棵决策树, 为了避免叶子节点过多或过少对拟合效果的不利影响, 本研究统计了两个数据集每年生长季尺度的气象要素、SMRoot和GPP的面积加权平均值, 以袋外分位数误差的最小化为优化目标, 基于贝叶斯全局优化算法对最小叶子节点数进行优化(参数优化结果如表 3所示)。
| 数据集生成方式 Dataset generation method |
最小叶子节点数 The minimum number of leaf nodes |
袋外分位数误差最优解 Optimal solution for out-of-pocket quantile error |
| 基于VIP数据集汇总Summarized based on dataset VIP | 2 | 0.0012 |
| 基于气温数据汇总Summarized based on air temperature data | 3 | 0.0011 |
| VIP:植被指数与物候产品Vegetation index and phenology | ||
对于每个栅格, 所有的输入、输出变量都经过了归一化处理, 并进行了33轮模拟, 每一轮选取某一年作为验证集, 其余32年作为测试集, 因此在33轮模拟后可汇总出一组完整的验证集。为了消除实际值对精度评估的影响, 我们计算了每个栅格上验证集GPP序列与该数据集生长季GPP序列的均方根偏差率作为评价指标, 以评估算法的整体性能:

|
(3) |
式中:RMSE Ratio表示均方根偏差率(无量纲), RMSE表示验证集GPP序列与生长季GPP的均方根误差(kgC/m2), Mean表示生长季GPP序列均值(kgC/m2)。我们对全球旱地范围内每个栅格计算所得的均方根偏差率进行平均汇总, 实现了较为良好的模拟精度, 结果如表 4所示。
| 数据集生成方式 Dataset generation method |
均方根偏差率 Root mean square deviation rate |
| 基于VIP数据集汇总Summarized based on dataset VIP | 0.1719 |
| 基于气温数据汇总Summarized based on air temperature data | 0.1631 |
基于模拟实验的思想, 植被生产力对各个因子的敏感性是通过计算正常模型运行情境模拟的植被生产力与对比试验情境模拟结果的差值与二者控制因子差异的最小二乘拟合斜率得以量化[25]。具体地, βVPD, βSMRoot的计算方法基于以下公式:

|
(4) |

|
(5) |
式中: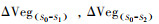
本研究采用分档平均分析方法以最大限度地减少由于空间异质性(地形、土壤性质等因素)对研究结果带来的不确定性[26], 本研究将干旱指数以一定的间隔划分子区间, 对每一子区间内所有栅格对应的敏感度大小进行汇总, 计算出各个分箱内敏感度的平均值。在后续的结果展示中, 分档平均结果将用于表征全球旱地植被生产力对根区土壤含水率和VPD的敏感性的一般关系。
2 结果与分析 2.1 全球旱地植被总初级生产力、饱和水汽压差及根区土壤含水率变化的空间分布本文依据不同生长季定义, 汇总所得全球旱地植被生长季GPP, 饱和水汽压差(VPD)以及根区土壤含水率的变化趋势如图 1所示。图 1第一行分别表征了全球旱地VPD生长季均值变化趋势的空间分布及其变化显著区域, 结果表明, 1982至2014年全球旱地约82.5%的植被覆盖区域生长季平均VPD呈显著上升趋势, 平均总体增幅为3Pa/a (P < 0.01), 其中在北美洲西南部、南美洲西南部、非洲东北部均捕捉到VPD的明显上升。此外, 全球旱地约62.9%的植被覆盖区域生长季平均GPP呈现显著上升趋势, 平均总体增幅为8.8 gC m-2 10a-1 (P < 0.01), 其中在北美洲旱地东北部、南亚地区、澳大利亚东北部增幅较大。然而, 全球旱地生长季平均根区土壤含水率总体无显著变化趋势(P>0.5), 尽管在非洲南部地区根区土壤含水率呈现显著上升趋势, 但在美国西部、中国东北地区以及中亚部分地区仍呈现较强的下降趋势。上述分析结果表明, 在全球气候变暖背景下, 全球旱地植被生产力的上升往往伴随着大气水分亏缺量的增加, 且生长季根区土壤水分的下降在北半球干旱地区更为普遍。

|
| 图 3 全球旱地植被总初级生产力、饱和水汽压差及根区土壤含水率变化的空间分布 Fig. 3 Spatial distribution of changes in gross primary productivity, vapor pressure deficit and root-zone soil moisture across global drylands |

|
| 图 4 全球旱区总初级生产力对饱和水汽压差及根区土壤含水率的敏感性 Fig. 4 Sensitivity of gross primary productivity to vapor pressure deficit and root-zone soil moisture in global drylands SMroot:根区土壤湿度Root-zone soil moisture |
为了探究全球旱地不同类型植被生长对根区土壤水分和饱和水汽压差的敏感性是否存在异质性特征, 本文基于GLASS-GLC土地覆盖分类产品统计出农田、森林、草地、灌丛以及苔原五种植被类型的βSMRoot以及βVPD数值, 汇总结果如图 5所示。图 5表明, 相较于农田、草地, 森林、灌丛植被种群对根区土壤水分的敏感性较弱, 这说明单位根区土壤含水率的上升对森林、灌丛植被生长的正效应弱于农田、草地和苔原, 然而饱和水汽压差在农田、草地、苔原植被系统会引发更加强烈的负效应。通过上述分析可以得出:在全球旱地区域, 植被生产力对根区土壤水分和饱和水汽压差的敏感性在不同类型植被之间存在异质性, 且相比于森林和灌丛群落, 根区土壤水分和饱和水汽压差的单位变化会在农田、草地的响应更为强烈。这表明全球旱地区域的森林和灌丛种群抵御饱和水汽压差升高和土壤水分短缺对植被生长不利影响的能力可能更优。

|
| 图 5 全球旱地不同种类植被总初级生产力对根区土壤水含水率和饱和水汽压差敏感性箱线图 Fig. 5 Boxplot of the sensitivity of gross primary productivity to root-zone soil moisture and vapor pressure deficit in different vegetation species across global drylands ERAS-Land:欧洲第五代陆面组分再分析数据集; VIP:植被指数与物候产品Vegetation index and phenology; TMP:气温Temperature |
为了探究植被生产力对根区土壤含水率和饱和水汽压差的敏感性随气象干旱条件变化的规律, 本文将干旱指数以0.15为间隔分为四个区间, 并统计出不同所有栅格对应的βSMRoot、βVPD数值, 如图 6所示。由图 6可知, 植被生产力对根区土壤水分以及饱和水汽压差的敏感性随干旱指数变化呈现非线性关系, 且ERA5-Land(VIP), ERA5-Land(TMP)两个数据集一致表征了植被生产力对根区土壤水分、饱和水汽压差的敏感性随干旱指数变化趋势的相反性, 即随着气候类型由干旱气候转变为半干旱气候, 根区土壤水分对植被生长的正效应以及饱和水汽压差对植被生长的负效应将不断增强, 但随着气候类型转变为半湿润气候, 根区土壤水分对植被生长的正效应以及饱和水汽压差对植被生长的负效应呈减弱态势。

|
| 图 6 植被生产力对饱和水汽压差及根区土壤水分敏感性随干旱指数关系箱线图 Fig. 6 Boxplot of the relationship between the sensitivity of gross primary productivity to vapor pressure deficit and root-zone soil moisture with aridity index |
在此基础上, 本研究将全球旱地范围内的所有栅格对应的βSMRoot、βVPD数值以0.03为间隔划分为20个分箱, 并对各个分箱的平均值序列进行线性拟合, 拟合结果如图 7所示。由图 7可知:ERA5-Land(VIP), ERA5-Land(TMP)两个数据集一致表征了植被生产力对根区土壤水和饱和水汽压差敏感性的分箱均值存在显著的线性负相关关系。因此, 从整体上而言, 全球旱地区域内植被生产力对根区土壤水分和饱和水汽压差的敏感性在不同干旱分区存在着明显的异质性, 且根区土壤水分和饱和水汽压差对植被生长的影响大小存在着较强的相关性, 而植被所处地区的气候条件的差异是形成这一关系的重要影响因素。

|
| 图 7 不同气象干旱条件下植被总初级生产力对根区土壤含水率及饱和水汽压差敏感性分箱均值的数量关系 Fig. 7 Quantitative relationship between the binned average sensitivity of gross primary productivity to root-zone soil moisture and vapor pressure deficit under different meteorological drought conditions |
本研究表征了不同植被类型对大气水分亏缺和根区土壤含水率变化的响应存在异质性, 这与以往植被对干旱的响应研究的结论具有一定的相似性。Flach等[27]指出, 在全球范围内, 相较于其他植被类型, 森林对于干旱事件具有较低的敏感性。张更喜等[28]对基于1982—2015年中国地区的NDVI及改进帕尔默干旱指数(scPDSI)表明草地对干旱的敏感性强于林地。Sun等[29]基于MODIS、FLUXCOM数据集以及多款陆面模式生成的GPP产品计算出GPP与标准化降水蒸散发指数(SPEI)之间的最大相关系数, 结果表明农田和草地的GPP与干旱的相关性最强, 其次是灌丛和森林。这一现象与不同植被群落的结构、根区分布及其所处气候条件等方面的差异关系密切[30]。
本研究发现, 基于不同生长季定义汇总所得数据集计算所得全球旱地区域植被生产力对大气饱和水汽压差和根区土壤含水率的敏感性随气象干旱指数变化均表现出一致的变化规律。尽管不同干旱分区存在水分可利用性的差异, 然而半干旱地区植被与干旱、半湿润地区植被生态群落对于水分缺失的生理响应机制存在差异, 由于干旱地区的植被受到更强的水分胁迫, 这促使这些地区的植被生态系统在生理和功能策略上采取主动进化以快速应对水分的短缺[31], 而半湿润地区的植被群落相较于干旱地区水分可利用性较强, 这有助于减轻水分亏缺对植被的影响, 相较于干旱区和半干旱半湿润地区, 半干旱区的植被会经历较长时间尺度的水分亏缺[32], 这可能对植被生长带来更强的的负效应。由于在全球变暖背景下饱和水汽压差-土壤水分的耦合将进一步增强, 这一正反馈过程使得大气和土壤水分亏缺对植被生长的负效应的单一影响会进一步放大[33], 同时这一过程对植被生长的负效应将以低土壤水分和高VPD相关的水分胁迫和固碳量减小过程来体现, 且该趋势将进一步加剧[34], 因此未来的研究需要进一步完善有关复合极端事件对植被固碳影响的评估。
4 结论本文量化了全球旱地植被生产力对大气饱和水汽压差和根区土壤水分的敏感性, 并结合土地覆盖分类数据及分档平均统计分析方法探究了敏感性在不同植被类型和气候条件下的异质性特征, 得出如下结论:
(1) 全球旱地生长季大气饱和水汽压差及植被生产力呈普遍增长态势, 而根区土壤含水率呈现较强的空间异质性, 且总体无显著变化趋势。
(2) 全球旱地区域根区土壤含水率对植被总初级生产力总体呈正敏感性, 饱和水汽压差对植被总初级生产力总体呈负敏感性。
(3) 相较于灌丛和森林, 农田、草地以及苔原类型植被的生长对根区土壤水分的敏感性更强, 且饱和水汽压差对这些种类植被生长的负效应更为强烈。
(4) 根区土壤水分对植被生长的正效应随干旱指数增加总体呈先增后减态势, 而饱和水汽压差对植被生长的负效应随干旱指数上升呈先减后增态势, 且二者在数量上呈现显著的线性负相关关系。因此, 相较于地处半干旱区的植被, 地处干旱区以及半湿润地区的植被能够更好应对气候变化背景下大气水分亏缺量上升对植被生长带来的威胁。
| [1] |
Wang S, Fu B J, Wei F L, Piao S L, Maestre F T, Wang L X, Jiao W Z, Liu Y X, Li Y, Li C J, Zhao W W. Drylands contribute disproportionately to observed global productivity increases. Science Bulletin, 2023, 68(2): 224-232. DOI:10.1016/j.scib.2023.01.014 |
| [2] |
Han D L, Huang J P, Ding L, Liu X Y, Li C Y, Yang F. Oxygen footprint: An indicator of the anthropogenic ecosystem changes. Catena, 2021, 206. |
| [3] |
Hoover D L, Bestelmeyer B, Grimm N B, Huxman T E, Reed S C, Sala O, Seastedt T R, Wilmer H, Ferrenberg S. Traversing the Wasteland: A Framework for Assessing Ecological Threats to Drylands. Bioscience, 2020, 70: 35-47. DOI:10.1093/biosci/biz126 |
| [4] |
Zhu Z C, Piao S L, Myneni R B, Huang M T, Zeng Z Z, Canadell J G, Ciais P, Stich S, Friedlingstein P, Arneth A, Cao C X, Cheng L, Kato E, Koven C, Li Y, Lian X, Liu Y W, Liu R G, Mao J F, Pan Y Z, Peng S S, Penuelas J, Poulter B, Pugh T A M, Stocker B D, Viovy N, Wang X H, Wang Y P, Xiao Z Q, Yang H, Zaehle S, Zeng N. Greening of the Earth and its drivers. Nature Climate Change, 2016, 6: 791. DOI:10.1038/nclimate3004 |
| [5] |
Green J K, Seneviratne S I, Berg A M, Findell K L, Hagemann S, Lawrence D M, Gentine P. Large influence of soil moisture on long-term terrestrial carbon uptake. Nature, 2019, 565: 476. DOI:10.1038/s41586-018-0848-x |
| [6] |
Sperry J S, Love D M. What plant hydraulics can tell us about responses to climate-change droughts. New Phytologist, 2015, 207: 14-27. DOI:10.1111/nph.13354 |
| [7] |
Gentine P, Guerin M, Uriarte M, McDowell N G, Pockman W T. An allometry-based model of the survival strategies of hydraulic failure and carbon starvation. Ecohydrology, 2016, 9: 529-546. DOI:10.1002/eco.1654 |
| [8] |
Medlyn B E, Duursma R A, Eamus D, Ellsworth D S, Prentice I C, Barton C V M, Crous K Y, Angelis P De, Freeman M, Wingate L. Reconciling the optimal and empirical approaches to modelling stomatal conductance. Global Change Biology, 2012, 18: 3476. DOI:10.1111/j.1365-2486.2012.02790.x |
| [9] |
Fletcher A L, Sinclair T R, Allen L H. Transpiration responses to vapor pressure deficit in well watered 'slow-wilting' and commercial soybean. Environmental and Experimental Botany, 2007, 61: 145-151. DOI:10.1016/j.envexpbot.2007.05.004 |
| [10] |
Zhou S, Zhang Y, Williams A P, Gentine P. Projected increases in intensity, frequency, and terrestrial carbon costs of compound drought and aridity events. Science Advances, 2019, 5. |
| [11] |
Zhou S, Williams A P, Berg A M, Cook B I, Zhang Y, Hagemann S, Lorenz R, Seneviratne S I, Gentine P. Land-atmosphere feedbacks exacerbate concurrent soil drought and atmospheric aridity. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116: 18848-18853. |
| [12] |
Li W T, Migliavacca M, Forkel M, Denissen J M C, Reichstein M, Yang H, Duveiller G, Weber U, Orth R. Widespread increasing vegetation sensitivity to soil moisture. Nature Communications, 2022, 13. |
| [13] |
Berg A, Mccoll K A. No projected global drylands expansion under greenhouse warming. Nature Climate Change, 2021, 11: 331-U371. DOI:10.1038/s41558-021-01007-8 |
| [14] |
Zhong Z Q, He B, Wang Y P, Chen H W, Chen D L, Fu Y H, Chen Y N, Guo L L, Deng Y, Huang L, Yuan W P, Hao X M, Tang R, Liu H M, Sun L Y, Xie X M, Zhang Y F. Disentangling the effects of vapor pressure deficit on northern terrestrial vegetation productivity. 2023, Science Advances, 9.
|
| [15] |
Wang S H, Zhang Y G, Ju W M, Qiu B, Zhang Z Y. Tracking the seasonal and inter-annual variations of global gross primary production during last four decades using satellite near-infrared reflectance data. Science of the Total Environment, 2021, 755. |
| [16] |
Zhang X Y, Tan B, Yu Y Y. Interannual variations and trends in global land surface phenology derived from enhanced vegetation index during 1982-2010. International Journal of Biometeorology, 2014, 58: 547-564. DOI:10.1007/s00484-014-0802-z |
| [17] |
Munoz-Sabater J, Dutra E, Agusti-Panareda A, Albergel A, Arduini G, Balsamo G, Boussetta S, Choulga M, Harrigan S, Hersbach H, Martens B, Miralles D G, Piles M, Rodriguez-Fernandez N J, Zsoter E, Buontempo C, Thepaut J N. ERA5-Land: a state-of-the-art global reanalysis dataset for land applications. Earth System Science Data, 2021, 13(9): 4349-4383. DOI:10.5194/essd-13-4349-2021 |
| [18] |
Miralles D G, Holmes T R H, De Jeu R A M, Gash J H, Meesters A G C A, Dolman A J. Global land-surface evaporation estimated from satellite-based observations. Hydrology and Earth System Sciences, 2011, 15(2): 453-469. DOI:10.5194/hess-15-453-2011 |
| [19] |
Martens B, Miralles D G, Lievens H, Van Der Schalie R, De Jeu R A M, Fernandez-Prieto D, Beck H E, Dorigo W A, Verhoest N E C. GLEAM v3: satellite-based land evaporation and root-zone soil moisture. Geoscientific Model Development, 2017, 10: 1903-1925. DOI:10.5194/gmd-10-1903-2017 |
| [20] |
Liu H, Gong P, Wang J, Clinton N, Bai Y Q, Liang S L. Annual dynamics of global land cover and its long-term changes from 1982 to 2015. Earth System Science Data, 2020, 12: 1217-1243. DOI:10.5194/essd-12-1217-2020 |
| [21] |
Wang S H, Zhang Y G, Ju W M, Chen J M, Ciais P, Cescatti A, Sardans J, Janssens I A, Wu M S, Berry J A, Campbell E, Fernandez-Martinez M, Alkama R, Sftch S, Friedlingstein P, Smith W K, Yuan W P, He W, Lombardozzi D, Kautz M, Zhu D, Lienert S, Kato E, Poulter B, Sanders T G M, Kruger I, Wang R, Zeng N, Tian H Q, Vuichard N, Jain A K, Wiltshire A, Goll D S, Penuelas J. Recent global decline of CO2 fertilization effects on vegetation photosynthesis. Science, 2020, 370: 1295. DOI:10.1126/science.abb7772 |
| [22] |
Chen Z F, Wang W G, Cescatti A, Forzieri G. 2023. Climate-driven vegetation greening further reduces water availability in drylands. Global Change Biology, 29: 1628-1647.
|
| [23] |
宋进喜, 齐贵增, 佘敦先, 蒋晓辉, 毛瑞晨. 中国植被生产力对干湿变化的响应. 地理学报, 2023, 78(7): 1764-1778. |
| [24] |
Liu X F, Sun G P, Fu Z, Ciais P, Feng X M, Li J, Fu B J. Compound droughts slow down the greening of the Earth. Global Change Biology, 2023, 29: 3072-3084. DOI:10.1111/gcb.16657 |
| [25] |
Yuan W P, Zheng Y, Piao S L, Ciais P, Lombardozzi D, Wang Y P, Ryu Y, Chen G X, Dong W J, Hu Z M, Jain A K, Jiang C Y, Kato E, Li S H, Lienert S, Liu S G, Nabel J E M S, Qin Z C, Quine T, Sitch S, Smith W K, Wang F, Wu C Y, Xiao Z Q, Song Y. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Science Advances, 2019, 5. |
| [26] |
Chen Z F, Wang W G, Yu Z B, Xia J, Schwartz F W. The collapse points of increasing trend of vegetation rain-use efficiency under droughts. Environmental Research Letters, 2020, 15. |
| [27] |
Flach M, Brenning A F, Gans F, Reichstein M, Sippel S, Mahecha M D. Vegetation modulates the impact of climate extremes on gross primary production. Biogeosciences, 2021, 18: 39-53. DOI:10.5194/bg-18-39-2021 |
| [28] |
张更喜, 粟晓玲, 郝丽娜, 吴海江. 基于NDVI和scPDSI研究1982-2015年中国植被对干旱的响应. 农业工程学报, 2019, 35(20): 145-151. |
| [29] |
Sun S B, Du W L, Song Z L, Zhang D B, Wu X C, Chen B Z, Wu Y T. Response of Gross Primary Productivity to Drought Time-Scales Across China. Journal of Geophysical Research-Biogeosciences, 2021, 126. |
| [30] |
刘静, 温仲明, 刚成诚. 黄土高原不同植被覆被类型NDVI对气候变化的响应. 生态学报, 2020, 40(2): 678-691. DOI:10.5846/stxb201901090082 |
| [31] |
Vicente-Serrano S M, Gouveia C, Camarero J J, Begueria S, Trigo R, Lopez-Moreno J I, Azorin-Molina C, Pasho E, Lorenzo-Lacruz J, Revuelto J, Moran-Tejeda E, Sanchez-Lorenzo A. Response of vegetation to drought time-scales across global land biomes. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110: 52-57. |
| [32] |
Chaves M M, Maroco J P, Pereira J S. Understanding plant responses to drought-from genes to the whole plant. Functional Plant Biology, 2003, 30: 239-264. DOI:10.1071/FP02076 |
| [33] |
Liu L B, Gudmundsson L, Hauser M, Qin D H, Li S C, Seneviratne S I. Soil moisture dominates dryness stress on ecosystem production globally. Nature Communications, 2020, 11. |
| [34] |
Lian X, Piao S L, Chen A P, Wang K, Li X Y, Buermann W, Huntingford C, Penuelas J, Xu H, Myneni R B. Seasonal biological carryover dominates northern vegetation growth. Nature Communications, 2021, 12. |