 2024, Vol. 44
2024, Vol. 44文章信息
- 刘子琪, 姚良锦, 焦洁洁, 陆超, 王志高, 李婷婷, 袁位高, 江波, 吴初平
- LIU Ziqi, YAO Liangjin, JIAO Jiejie, LU Chao, WANG Zhigao, LI Tingting, YUAN Weigao, JIANG Bo, WU Chuping
- 亚热带次生常绿阔叶林植物功能性状及多样性对人工抚育的响应
- Response of functional traits and functional diversity of woody plants to tending in a secondary subtropical evergreen broad-leaved forest
- 生态学报. 2024, 44(11): 4733-4743
- Acta Ecologica Sinica. 2024, 44(11): 4733-4743
- http://dx.doi.org/10.20103/j.stxb.202307251589
-
文章历史
- 收稿日期: 2023-07-25
- 网络出版日期: 2024-03-18






2. 浙江省林业科学研究院, 杭州 310023
2. Zhejiang Forestry Academy, Hangzhou 310023, China
植物的功能性状是指那些对于植物的生存、生长和繁殖具有适应性和效应特征的特征[1—2], 这些特征可以根据环境因素的变化做出响应, 并且直接参与生态系统的过程, 从而对生态系统的服务产生影响[3—4]。其中的环境因素包括气候[5]、地形[6]和土壤[7]等自然环境和火灾[8]、人为干扰[9]和土地利用[10]等干扰环境。植物的不同功能性状之间也存在着紧密的内在联系与权衡关系, 显著影响着生物群落的功能特征[11—12]。研究群落功能性状之间的权衡关系, 可以揭示植物的生长策略和分配资源的模式, 探索植物生长生理生态过程的内在机制, 并为生态系统的能量与物质动态的研究提供依据, 有助于更加准确地预测植物群落对气候变化的响应[12—13]。植物功能性状多样性是生物多样性的重要组成, 是生态系统功能或过程的主要决定者[14—16]。Cadotte等[17]在美国明尼苏达州的长期实验研究显示, 植物的功能性状多样性对环境胁迫或扰动更具敏感性, 它相较于物种多样性可以更好地指示生态系统功能的变化。同时, 它们也能够更清楚地解释群落中物种多样性与生态系统功能之间的关系。因此, 对植物功能性状进行深入研究, 能够找寻植物共存与群落稳定发展的内在机制, 为提升森林生态系统功能的实践提供基础。
森林自然演替需要的时间十分漫长, 若能在更新过程中采取合理正确的人工抚育, 便能加快次生林向顶级森林群落演替的进程[18]。刘佳佳[19]在西双版纳的研究表明, 包括抚育在内的环境因素是驱动片段化生境内功能性状组成和功能多样性指数分布格局的主要因素;Carreño-Rocabado等[20]在玻利维亚热带低地的研究发现, 植物功能性状变化沿土地利用强度梯度呈驼峰状变化;而另一在热带雨林中的研究也得出了干扰强度会影响群落水平功能性状值的结论[21]。因此, 开展不同强度的抚育措施对天然次生林群落植被功能性状影响的研究, 能够有助于理解抚育后森林群落演替的机理和过程, 从而为制订合理的森林管理制度提供科学依据。路兴慧等[22]按照不同功能群的生态适应性及其演替的变化规律, 划分不同目标树种类别, 对海南岛热带天然林进行抚育后的结果显示, 抚育措施能够促使植物的生态策略向老龄林方向转化, 提高物种的资源利用效率, 同时增强生态系统服务功能。这种在植物功能群的基础上进行的抚育方式为森林抚育途径提供良好参考, 但其是否能够对具有复杂干扰历史、植被较复杂的亚热带次生常绿阔叶林产生同样的积极影响尚不明确。
亚热带常绿阔叶林是世界主要森林植被类型之一, 在涵养水源、维持生物多样性、固碳释氧等方面具有较大的优势[23—24], 但中国的绝大部分亚热带常绿阔叶林都由于人类长期的干扰而退化为次生林, 其在演替过程中稳定性差, 并且普遍存在生长滞缓、固碳功能和经济价值低下等问题[25—26]。因此, 为了探究人工抚育对亚热带次生常绿阔叶林质量提升的影响, 本实验前期对亚热带次生常绿阔叶林样地实施了不同强度的人工抚育, 通过群落植被复查, 分析群落植物功能性状以及功能多样性对不同强度人工抚育的响应, 旨在探讨不同抚育强度会如何影响亚热带次生常绿阔叶林群落的功能性状组成和功能多样性指数, 从而为亚热带次生常绿阔叶林的经营与管理提供依据与参考。
1 研究区概况午潮山国家森林公园位于杭州西郊(33°41′N, 120°00′E), 为天目山余脉, 平均海拔254m, 主峰海拔504m。属中亚热带季风气候, 年降雨量1400.7mm, 年平均相对湿度为76%。年均气温16.1℃, 极端最高气温39.0℃, 极端最低气温-9.4℃, 其中温度7月最高, 1月最低, 达到3.6℃, 年平均日照时数为1800—2100h, 年无霜期为240d。土壤类型为红壤[27—28]。
午潮山林区内的森林植被为亚热带常绿次生阔叶林, 其森林覆盖率达到98.2%。从近代开始, 在林区内的人工开发对区域内原始植被造成了严重破坏。自1960年起, 浙江省林业科学研究院将该地视为科研基地, 并采取措施进行保护。经过长期的封山育林, 人工种植了大量的经济树种以保护和恢复森林。至今该地的森林已逐渐演变为常绿次生阔叶林[29]。研究区森林群落乔木层的主要树种包括青冈(Cyclobalanopsis glauca)、木荷(Schima superba)和麻栎(Quercus acutissima)等, 灌木层的主要树种为马银花(Rhododendron ovatum)、毛花连蕊茶(Camellia fraterna)、窄基红褐柃(Eurya rubiginosa var. attenuata)和薄叶山矾(Symplocos anomala)等。
2 方法 2.1 样地设置及调查本研究按照热带森林科学研究中心的原则, 于2020年在浙江省林业科学研究院实验林场午潮山林区的次生常绿阔叶林内, 利用全站仪设置了海拔、坡度、坡向等生境特征和林分结构均较为相似的30个20m×20m的样地, 对样地内胸径≥1cm的存活木本树种进行标定并挂牌, 并对树种名称、胸径、树高和空间坐标等信息进行记录。同年8月对20个样地进行2种强度的抚育, 剩余10个样地作为对照。后于2022年8月对所有样地进行植物群落复查。
2.2 抚育方法本实验前期对午潮山次生常绿阔叶林内45种主要木本植物进行功能性状收集和测量, 将群落内主要物种按照演替进程划分为4个功能群:早期先锋树种、中期中性树种、中后期树种和后期耐阴树种[30]。2020年8月, 在已划分的植物功能群的基础上, 对树种的生态特性和木材价值进行考量, 最后将研究区树种划分为目的树种、辅助树种和清除树种[22](表 1)进行抚育实验。对目的树种原则上保留, 仅对萌芽丛伐除萌芽条保留1株最有生长前途的主干;对清除树种进行整株伐除;对辅助树种依据生长状况以及是否阻碍目的树种生长等酌情进行伐除或保留;为了避免造成过大的林窗, 仅对胸径20cm以下的个体进行抚育, 保留超过该径级的所有树木。根据抚育前后样地内单位面积的胸高断面积比变化计算得出抚育强度, 其中10个样地为10%—15%、10个样地为20%—25%、10个样地未抚育作为对照(表 2)。抚育前后样地内主要树种的重要值变化详见表 3。
| 类别 Categories |
树种 Species |
特征描述 Feature description |
| 目的树种 Target species |
木荷、麻栎、青冈、檫木、石栎、冬青、湿地松、苦槠、短梗冬青、朴树、山矾、黄檀、化香、光叶山矾、光叶石楠、盐肤木、红楠、小叶青冈、油桐、短柄枹栎、刺楸、小蜡等 | 处于演替中期的生态关键种经济价值、观赏价值和生态价值高的树种 |
| 辅助树种 Auxiliary species |
毛花连蕊茶、薄叶山矾、大青、马银花、窄基红褐柃、白栎、老鼠矢、宁波木樨、檵木、格药柃、江南越桔、宜昌荚迷、红淡比、四川山矾、微毛柃、铁冬青等 | 与目的树种成正联结的树种能帮助恢复森林林窗的树种 |
| 清除树种 Cleared species |
小叶石楠、柃木、老鸦糊、白檀、毛漆树、宜昌荚蒾、毛八角枫、豆腐柴、映山红、山胡椒、乌饭、乌药、木蜡树、红果山胡椒、荚蒾、垂丝海棠等 | 与目的树种成负联结的树种对目的树种生长有较大干扰的树种 |
| 组别 Groups |
处理强度 Tending intensity/% |
胸高断面积 Basal area/(m2/hm2) |
密度 Number of plants/(株/hm2) |
年均生长率 Average annual production rate/% |
|||||
| 2020年 | 2022年 | 2020年 | 2022年 | ||||||
| 抚育前 | 抚育后 | 抚育前 | 抚育后 | ||||||
| 对照Control group | 0 | 36.56 | — | 37.90 | 6465 | — | 5880 | 1.84 | |
| 处理1 Treatment group 1 | 10—15 | 37.07 | 32.03 | 35.50 | 6738 | 3896 | 3978 | 5.42 | |
| 处理2 Treatment group 2 | 20—25 | 35.81 | 28.22 | 30.46 | 7008 | 3713 | 3851 | 3.97 | |
| 树种 Species |
20%抚育强度重要值 Important value pre- and post- 20% intensity tending/% |
10%抚育强度重要值 Important value pre- and post- 10% intensity tending/% |
||
| 抚育前 Pre-tending |
抚育后 Post-tending |
抚育前 Pre-tending |
抚育后 Post-tending |
|
| 木荷Schima superba | 10.19 | 10.46 | 10.34 | 11.32 |
| 麻栎Quercus acutissima | 9.52 | 9.78 | 11.37 | 11.56 |
| 毛花连蕊茶Camellia fraterna | 8.26 | 9.26 | 8.41 | 8.53 |
| 湿地松Pinus elliottii | 8.08 | 9.91 | 8.25 | 10.13 |
| 杉木Cunninghamia lanceolata | 5.69 | 4.62 | 3.93 | 2.66 |
| 格药柃Eurya muricata | 3.00 | 3.56 | 3.14 | 3.27 |
| 马银花Rhododendron ovatum | 2.97 | 2.35 | 4.38 | 3.07 |
| 苦槠Castanopsis sclerophylla | 2.44 | 3.34 | 2.67 | 3.41 |
| 薄叶山矾Symplocos anomala | 2.32 | 1.57 | 3.96 | 2.94 |
| 山胡椒Lindera glauca | 0.90 | 0.37 | 0.82 | 0.69 |
植物功能性状主要针对样地内胸径超过1cm的全部木本植物个体进行取样, 主要包括枝条密度(g/cm3)、潜在最大树高(m)、比叶面积(cm2/g)、叶片干物质含量(g/g)、叶片叶绿素含量(SPAD)、叶片氮含量(mg/g)、叶片磷含量(mg/g)、叶片钾含量(mg/g)等。这些功能性状均直接反映了自然条件下的树木生长速度、竞争能力、资源利用策略等[31]。
在30个20m×20m样方内, 采集每株木本植物个体的5枚阳性成熟叶, 对所有叶片进行形态和养分含量分析。利用扫描仪测定植物叶片面积参数, 利用叶绿素仪测定叶片叶绿素含量, 利用电子天平测定叶片鲜重, 置于80℃烘箱内至恒重后利用电子天平测定叶片的干重, 并将干燥后的叶片带回实验室做叶片养分含量分析。采用凯氏定氮仪法测定全氮含量;采用钒钼黄吸光光度法测定全磷含量;采用火焰原子吸收分光光度法测定全钾含量[32]。其中比叶面积(单位:cm2/g)=叶片面积/叶片干重, 干物质含量(单位:g/g)=叶片干重/叶片鲜重。每株木本植物个体依据芽鳞痕数目及其木质化程度取非一年生枝条3—5cm, 去皮后利用电子天平及其密度组件通过排水法测定枝干样品密度及鲜重, 然后置于80℃烘箱烘干至少72h至恒重, 称取干重。
2.4 数据计算及分析表 4展示了功能性状及功能多样性各指数的计算公式, 其中数据分析及制图均在R 4.1.1中完成, 其中功能性状及功能多样性指数使用R语言FD包中的dbFD函数计算, 不同抚育强度之间功能性状及功能多样性的差异使用One-Way ANOVA检验,功能性状之间相关性利用简单线性回归模型进行拟合。
| 指数 Index |
计算公式 Calculation formula |
解释 Interpretation |
| 群落水平功能性状(CWM) Community-level weighted means[33] |  |
物种水平的功能性状数值以物种多度加权平均。n为物种数;traiti为物种i的特征值。 |
| 功能丰富度(FRic) Functional richness[33] | 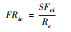 |
群落中物种占的以n维功能性状为基础的凹凸包量。SFci为群落i内物种所占据的生态位空间;Rc为特征c的绝对值范围。 |
| 功能均匀度(FEve) Functional evenness[34] |  |
群落中物种功能性状数值在凹凸包量中排列的规则性。S为物种丰富度;PEWi为物种i的局部加权均匀度。 |
| 功能分离度(FDiv) Functional divergence[33] | 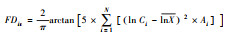 |
群落中物种功能性状数值在凹凸包量中排列的分散性。Ci为第i项功能特征值;Ai为第i项功能特征的相对丰富度;lnX为物种特征值自然对数的加权平均。 |
| 功能离散度(FDis) Functional dispersion[35] | 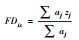 |
群落中每个物种的n维功能性状到所有功能性状空间重心的平均距离。aj为物种j的多度;zj为物种j到加权质心的距离。 |
两种强度的抚育对群落水平功能性状都有显著影响。抚育2年后, 枝条密度在20%强度显著下降, 10%强度有下降趋势;最大潜在树高与叶片的氮、磷、钾含量均显著提高;叶片干物质含量和叶片叶绿素含量均显著下降;比叶面积均有下降趋势。两种强度的抚育之间差异不显著, 但随着抚育强度的增加, 群落水平枝条密度指数逐渐降低, 而群落水平潜在最大树高逐渐升高(图 1)。

|
| 图 1 午潮山样地抚育2年后功能性状的变化(平均值±标准差) Fig. 1 Variations of community-level weight mean of plant functional traits (Mean±SD) after 2 years of tending in Wuchao Mountain sample plots 字母不同表示差异显著(P < 0.05) |
表 5展示了群落水平叶片功能性状间相关性的变化, 抚育2年后, 功能性状相关性有所变化, 叶片干物质含量、叶绿素含量与叶片氮含量间的相关性由不显著变为了显著负相关, 而叶片干物质含量与叶片叶绿素含量, 叶片磷、钾含量与叶片氮含量, 叶片磷含量与叶片钾含量间的相关性由不显著变为显著正相关;另外, 叶片叶绿素含量与枝条密度呈正相关, 而与叶片磷含量与最大潜在树高呈负相关, 并且之间相关性的斜率发生了变化;而在对照样地中比叶面积与叶片叶绿素含量和枝条密度呈正相关, 与叶片磷含量呈负相关, 但在经过抚育的样地中这些相关关系与前者相反(图 2)。
| 叶片功能性状 Leaf functional traits |
比叶面积 Specific leaf area |
叶片干物质含量 Leaf dry matter content |
叶片叶绿素含量 Leaf chlorophyll content |
叶片氮含量 Leaf nitrogen content |
叶片磷含量 Leaf phosphorus content |
| 叶片干物质含量 Leaf dry matter content |
-0.52/-0.82**/-0.53 | ||||
| 叶片叶绿素含量 Leaf chlorophyll content |
0.77**/-0.70*/-0.85** | -0.50/0.51/0.78** | |||
| 叶片氮含量 Leaf nitrogen content |
-0.31/0.93**/0.97** | 0.60/-0.66*/-0.51 | 0.09/-0.72*/-0.76* | ||
| 叶片磷含量 Leaf phosphorus content |
-0.68*/0.86**/0.95** | 0.84**/-0.60/-0.55 | -0.78**/-0.82**/-0.80** | 0.47/0.93**/0.98** | |
| 叶片钾含量 Leaf potassium content |
-0.58/0.20/0.66* | 0.39/-0.47/-0.10 | -0.25/0.32/-0.24 | 0.25/-0.01/0.80** | 0.28/-0.15/0.76* |
| “/”左、中、右侧分别是对照组、10%和20%强度抚育后的相关性指数; *表示相关性显著(P < 0.05); * *表示相关性极显著(P < 0.01) | |||||

|
| 图 2 午潮山样地抚育2年后功能性状相关性的变化 Fig. 2 Relationships between functional traits in stands after 2 years of tending in Wuchao Mountain sample plots |
抚育2年后, 群落的功能均匀度与功能分离度在20%强度显著提高, 10%强度有提高趋势;功能丰富度在20%强度显著下降, 10%强度有下降趋势;功能离散度均有下降趋势, 两种强度的抚育之间差异不显著。随着抚育强度的增加, 群落功能丰富度和功能离散度指数逐渐降低, 而群落功能均匀度和功能分离度指数逐渐升高(图 3)。

|
| 图 3 午潮山样地抚育2年后功能多样性的变化(平均值±标准差) Fig. 3 Variation of functional diversity (Mean±SD) after 2 years of tending in Wuchao Mountain sample plots 字母不同表示差异显著(P < 0.05) |
群落加权平均性状值可以很好地反映由于环境对某些功能性状的选择而导致的群落内平均性状值的变化[36]。植物叶片的功能性状与其利用资源的方式密切相关[31], 也是调节植物生态系统功能的关键特征之一[35]。在受到环境条件的长期影响下, 植物会逐渐形成适应自身资源获取方式的叶片特征构建机制[37]。Wright等[12]提出的叶片经济谱指出, 随着演替的进行, 叶片性状将从开放性的生长策略即比叶面积大、光合速率高但寿命短的策略, 而向保守性的生长策略即比叶面积小、光合速率低但寿命长的策略转变。本研究的结果显示, 经过抚育后的群落叶片的比叶面积、叶片干物质含量和叶片叶绿素含量都显著下降。这可能是由于抚育解放了目的树种的生存空间, 减小了种间竞争, 叶片中很大一部分物质可以用于构建保卫构造(防虫食、防止过度失水等)或者增加叶肉细胞密度所致, 同时群落水平比叶面积指数的降低意味着植物的生存策略在从开放性向保守性转变, 说明抚育后群落中常绿阔叶树种的优势度有所增加, 人工抚育加快了次生常绿阔叶林的恢复, 加速了阔叶林群落的演替进程。
植物养分含量(氮、磷、钾含量等)是反映植物光合能力和养分状况的生理功能指标[37]。抚育使森林群落及森林冠层结构得到改善, 为森林群落内树木的生长提供了充足的阳光及营养。本研究观察到抚育后的群落叶片氮含量有所增加, 有研究[38]表明叶片氮含量的增加可以通过调节光合酶的数量与活性, 提高CO2的同化率, 进而促进植物的光合作用效率。同时抚育后叶片养分含量的增加, 说明植物磷钾等养分供给充足, 这可能是由于抚育释放出了森林群落内的空间, 一定程度提高了保存下来的树木利用资源的能力和效率。叶经济谱的变化同样说明抚育后群落的生态策略正在从开放性向保守性策略转变。
植物的枝干功能性状反映了植株的稳定性、抗逆性和固碳能力等[39]。木质部密度较低的植物个体, 由于在导管运输组织上的低经济投入, 其生长速度相对较快[40]。反之, 密度较高的植物个体, 其生长速率和死亡率相对较低, 这是由于它们在单位体积的生物量投入大, 更能够抵抗环境变化[41], 因此木质部密度是反映植物在生长速率与耐受性之间权衡的重要指标。而我们的研究结果显示, 经过抚育后群落枝条密度有所降低, 这可能是抚育后样地采取的枝条样品包含大量两年生新枝所致。群落水平最大潜在高度能够反映群落垂直结构, 并与植物的竞争、恢复能力及生产力密切相关[39]。有研究表明在植被恢复过程中, 演替后期物种比先锋种的最大潜在高度高[42]。在本研究中, 演替后期种的数量在抚育后的群落中增加, 群落光竞争能力增强, 因此观察到群落水平最大潜在高度显著上升。
功能性状之间通常具有协同作用, 因此不能将其隔离开看待和分析[43]。植物受到环境的影响后, 经过长时间的适应, 会逐步形成其特有的功能性状组合, 如植株的潜在最大株高和叶片氮含量与植物的光合效率和资源竞争力密切相关[44];叶面积、比叶面积和叶片干物质含量等功能性状与同化效率有关[45]。功能性状之间的权衡关系反映了生物在适应环境变化过程中所采取策略的趋同性与趋异性[33, 42]。Rodríguez-Gallego等[46]的研究发现, 沿海沙丘植物的叶片氮、磷含量与比叶面积呈显著正相关, 也有研究发现草原群落中氮浓度与叶绿素浓度呈显著正相关[47], 本研究的结果也符合这些规律。实验结果还显示, 在两种强度的干扰下, 功能性状间的关系发生了变化, 其中在干扰下比叶面积与叶片磷含量、枝条密度、叶片叶绿素含量的相关关系与对照组相关性相反, 这可能是由于人工抚育砍伐了部分林分下层的耐阴树种所致, 另外本实验中观察到, 经过干扰的样地相较于对照组群落间的叶片氮含量与叶片磷含量和叶绿素含量、枝条密度、比叶面积之间, 以及叶片磷含量与枝条密度之间的相关关系都由不显著变为了显著, 叶片叶绿素含量与最大潜在树高之间的相关性系数也有所升高, 这说明植物的生存策略发生了变化。植物功能性状相互关系的影响因素是复杂的, 需要结合具体环境因子, 全面揭示植物功能性状之间的适应及响应机制。
综上所述, 对亚热带次生常绿阔叶林实施抚育后, 与对照组相比群落水平的植物功能性状发生了显著变化, 这些变化说明保留的树种能够更加充分地利用资源, 并且群落的生存策略在由开放性向保守性转变, 促进了群落的演替进程。
4.2 抚育措施与群落功能多样性植物的功能多样性能够很好地反映植物在群落中的空间分布特征和生态系统功能之间的相互关系, 从而为维持生态系统的稳定性提供重要的科学依据[33]。其中功能丰富度反映的是群落中存在的物种占据总性状空间的体积, 当环境过滤主导时, 部分性状被筛掉, 功能丰富度减小;反之相似性限制主导时, 功能丰富度增加[48]。有研究发现经过择伐后群落功能丰富度指数将呈现出先降低后升高的趋势[49]。本研究在抚育过程中砍伐了一些与目的树种呈负联结的树种及部分演替前期树种, 而2年内增加的更新树种相对较少, 导致功能丰富度在10%抚育强度下呈下降趋势, 在20%抚育强度下显著下降。
功能均匀度反映了植物的功能性状在群落中的分布一致性[50]。群落的功能均匀度高表明物种对资源的利用较为充分, 反之较低则表明不同的物种之间在一些资源上产生了竞争, 或者一些资源没有得到充分的利用[51]。在对高寒草甸的研究中发现, 更适宜生存的中海拔群落功能均匀度较高, 而高海拔地区的群落为了能够有效地保存资源, 功能均匀度较低[52]。本研究中, 群落的功能均匀度在10%抚育强度下有升高趋势, 在20%抚育强度下显著增高, 说明抚育措施解放了森林群落内部分功能性状空间, 使保留的树种对资源的利用更为全面、均一, 从而提高了各个种群对资源的利用效率与能力。
功能分离度反映了群落内各物种之间的生态位互补性[34]。功能分离程度越高, 则代表着不同物种之间的功能生态位重叠程度越小, 在同一资源上的竞争也越小[53]。已有研究发现, 功能分离度在不同大小的斑块中存在差异, 小斑块往往具有较高的功能分离度[54]。本实验的结果显示, 20%强度的人为干扰后, 群落的功能分离度显著增高, 这说明20%强度的抚育措施降低了群落内树种对资源的竞争, 增强了森林资源间的互补性, 进而提高了资源的使用效率, 改善生态系统的功能。
功能离散度指功能性状聚集在一起的物种组合的空间分布格局, 高的功能离散度表明森林群落受到相似影响的限制[35]。前人的研究表明竞争往往会导致性状的发散[55]。性状的发散模式说明物种间竞争的排除对群落构建过程有所贡献。赵家乐[56]的研究也证明环境过滤和相似性限制能够通过不同功能性状从而驱动本地群落构建过程。本实验中, 群落功能离散度变化并不显著, 但两种干扰强度后都呈降低趋势, 且20%干扰强度较10%干扰强度功能离散度降度更高, 说明干扰在一定程度上对群落中竞争强度起到降低作用, 群落的资源利用程度、群落生产力、稳定性及物种适应对策发生了变化。
综上所述, 人工抚育下的亚热带次生常绿阔叶林群落相比自然恢复群落的生态系统功能有显著提高, 并且20%强度较10%强度更能解放资源空间, 使树种可以更加充分且均匀地利用资源, 提高亚热带次生常绿阔叶林种群利用资源的效率和能力, 提高生态系统功能。
5 总结与展望本研究基于10%和20%抚育强度下的亚热带次生常绿阔叶林群落分析了群落植物功能性状和功能多样性对抚育强度的响应, 结果显示人工抚育一定程度改变了群落水平的植物功能性状与功能多样性, 提高了群落植物利用资源的能力和效率;而20%强度的抚育对群落产生的影响则更加明显。根据《浙江省公益林管理办法》规定, 公益林采伐强度不得超过伐前林分蓄积的15%。亚热带次生常绿阔叶林作为浙江省公益林的重要组成部分, 该研究结果对于进一步完善公益林的经营管理提供了科学的依据。同时, 鉴于本研究跨度时间和研究内容有限, 有必要持续跟踪, 进一步探讨抚育经营措施下地上和地下不同生物类群对亚热带次生常绿阔叶林碳储量及其组成稳定性的影响机制和驱动机理等。
| [1] |
Violle C, Navas M L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E. Let the concept of trait be functional!. Oikos, 2007, 116(5): 882-892. DOI:10.1111/j.0030-1299.2007.15559.x |
| [2] |
刘晓娟, 马克平. 植物功能性状研究进展. 中国科学: 生命科学, 2015, 45(4): 325-339. |
| [3] |
Eviner V T, Chapin Iii F S. Functional matrix: a conceptual framework for predicting multiple plant effects on ecosystem processes. Annual Review of Ecology Evolution & Systematics, 2003, 34(1): 455-485. |
| [4] |
Balvanera P, Pfisterer A B, Buchmann N, He J S, Nakashizuka T, Raffaelli D, Schmid B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters, 2006, 9(10): 1146-1156. DOI:10.1111/j.1461-0248.2006.00963.x |
| [5] |
Murray B R, Brown A H D, Dickman C R, Crowther M S. Geographical gradients in seed mass in relation to climate. Journal of Biogeography, 2004, 31(3): 379-388. DOI:10.1046/j.0305-0270.2003.00993.x |
| [6] |
胡耀升, 么旭阳, 刘艳红. 长白山森林不同演替阶段植物与土壤氮磷的化学计量特征. 应用生态学报, 2014, 25(3): 632-638. |
| [7] |
肖露梅, 张伟, 王彩艳, 胡培雷, 陈元凯, 王克林. 典型喀斯特区不同植被恢复方式苔藓功能性状及其对土壤因子的响应. 生态学报, 2022, 42(23): 9769-9779. |
| [8] |
孙兴悦, 张雨鉴, 秦倩倩, 白岩松, 刘艳红. 火干扰下白桦树干不同高度的树皮性状及其环境解释. 应用生态学报, 2023, 34(2): 315-323. |
| [9] |
符强, 王楠, 肖曼, 习靓靓, 邵毅贞, 贾宏汝, 陈云, 袁志良, 叶永忠. 白云山国家森林公园不同人为干扰强度的群落冠层结构和光照特征. 生态学报, 2021, 41(19): 7830-7837. |
| [10] |
文志, 赵赫, 刘磊, 王丽娟, 欧阳志云, 郑华, 李彦旻. 基于土地利用变化的热带植物群落功能性状与土壤质量的关系. 生态学报, 2019, 39(1): 371-380. |
| [11] |
Wright I J, Reich P B, Cornelissen J H C, Falster D S, Groom P K, Hikosaka K, Lee W, Lusk C H, Niinemets V, Oleksyn J, Osada N, Poorter H, Warton D I, Westoby M. Modulation of leaf economic traits and trait relationships by climate. Global Ecology and Biogeography, 2005, 14(5): 411-421. DOI:10.1111/j.1466-822x.2005.00172.x |
| [12] |
Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T L, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827. DOI:10.1038/nature02403 |
| [13] |
冯秋红, 史作民, 董莉莉. 植物功能性状对环境的响应及其应用. 林业科学, 2008, 44(4): 125-131. |
| [14] |
Diaz S, Cabido M. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology & Evolution, 2001, 16(11): 646-655. |
| [15] |
Díaz S, Lavorel S, de Bello F, Quétier F, Grigulis K, Robson T M. Incorporating plant functional diversity effects in ecosystem service assessments. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(52): 20684-20689. |
| [16] |
Roscher C, Schumacher J, Gubsch M, Lipowsky A, Weigelt A, Buchmann N, Schmid B, Schulze E D. Using plant functional traits to explain diversity-productivity relationships. PLoS One, 2012, 7(5): e36760. DOI:10.1371/journal.pone.0036760 |
| [17] |
Cadotte M W, Cavender-Bares J, Tilman D, Oakley T H. Using phylogenetic, functional and trait diversity to understand patterns of plant community productivity. PLoS One, 2009, 4(5): e5695. DOI:10.1371/journal.pone.0005695 |
| [18] |
李志明. 林龄对人工促进天然更新林林下层生源要素的影响. 亚热带资源与环境学报, 2020, 15(4): 47-51. |
| [19] |
刘佳佳. 非随机生境片段化过程对树种多样性的影响: 从物种到性状[D]. 北京: 中国科学院大学, 2014.
|
| [20] |
Carreño-Rocabado G, Peña-Claros M, Bongers F, Díaz S, Quetier F, Chuviña J, Poorter L. Land-use intensification effects on functional properties in tropical plant communities. Ecological Applications: a Publication of the Ecological Society of America, 2016, 26(1): 174-189. DOI:10.1890/14-0340 |
| [21] |
Carreño-Rocabado G, Peña-Claros M, Bongers F, Alarcón A, Licona J C, Poorter L. Effects of disturbance intensity on species and functional diversity in a tropical forest. Journal of Ecology, 2012, 100(6): 1453-1463. DOI:10.1111/j.1365-2745.2012.02015.x |
| [22] |
路兴慧, 臧润国, 丁易, 黄继红, 杨秀森, 周亚东. 抚育措施对热带次生林群落植物功能性状和功能多样性的影响. 生物多样性, 2015, 23(1): 79-88. |
| [23] |
张宏锦, 王娓. 生态系统多功能性对全球变化的响应: 进展、问题与展望. 植物生态学报, 2021, 45(10): 1112-1126. |
| [24] |
宋永昌, 陈小勇, 王希华. 中国常绿阔叶林研究的回顾与展望. 华东师范大学学报: 自然科学版, 2005(1): 1-8. |
| [25] |
丁圣彦, 宋永昌. 常绿阔叶林植被动态研究进展. 生态学报, 2004, 24(8): 1769-1779. |
| [26] |
田海静. 非气候因素引起的中国植被变化遥感诊断——以林业工程为例[D]. 北京: 中国科学院大学, 2017.
|
| [27] |
周重光, 许利群, 杭韵亚. 午潮山常绿阔叶林气候生态效应定位研究. 浙江林业科技, 1998, 18(2): 1-14. |
| [28] |
田立新, 吴初平, 杨少宗, 许玥, 黄继红, 丁易, 臧润国. 浙江杭州午潮山亚热带常绿阔叶林群落结构和物种组成. 应用生态学报, 2020, 31(6): 1909-1915. |
| [29] |
金超, 吴初平, 丁易, 袁位高, 朱锦茹, 黄玉洁, 焦洁洁, 臧润国, 江波. 午潮山常绿次生阔叶林主要木本植物功能群及其演替特征. 生态学报, 2021, 41(8): 3053-3066. |
| [30] |
金超. 午潮山次生常绿阔叶林植物功能群划分及其对干扰的响应[D]. 杭州: 浙江农林大学, 2021.
|
| [31] |
雷羚洁, 孔德良, 李晓明, 周振兴, 李国勇. 植物功能性状、功能多样性与生态系统功能: 进展与展望. 生物多样性, 2016, 24(8): 922-931. |
| [32] |
中华人民共和国农业部. 植物中氮、磷、钾的测定: NY/T 2017—2011[S]. 北京: 中国农业出版社, 2011: 1-7.
|
| [33] |
Mason N W H, Mouillot D, Lee W G, Wilson J B. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 2005, 111(1): 112-118. DOI:10.1111/j.0030-1299.2005.13886.x |
| [34] |
Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301. DOI:10.1890/07-1206.1 |
| [35] |
Laliberté E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305. DOI:10.1890/08-2244.1 |
| [36] |
Ricotta C, Moretti M. CWM and Rao's quadratic diversity: a unified framework for functional ecology. Oecologia, 2011, 167(1): 181-188. DOI:10.1007/s00442-011-1965-5 |
| [37] |
Lavorel S, Grigulis K, McIntyre S, Williams N S G, Garden D, Dorrough J, Berman S, Quétier F, Thébault A, Bonis A. Assessing functional diversity in the field-methodology matters!. Functional Ecology, 2008, 22(1): 134-147. DOI:10.1111/j.1365-2435.2007.01339.x |
| [38] |
赵平, 孙谷畴, 彭少麟. 植物氮素营养的生理生态学研究. 生态科学, 1998, 17(2): 37-42. |
| [39] |
Poorter L, McDonald I, Alarcón A, Fichtler E, Licona J C, Peña-Claros M, Sterck F, Villegas Z, Sass-Klaassen U. The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species. The New Phytologist, 2010, 185(2): 481-492. DOI:10.1111/j.1469-8137.2009.03092.x |
| [40] |
Navas M L, Roumet C, Bellmann A, Laurent G, Garnier E. Suites of plant traits in species from different stages of a Mediterranean secondary succession. Plant Biology, 2010, 12(1): 183-196. DOI:10.1111/j.1438-8677.2009.00208.x |
| [41] |
Slik J W, Bernard C S, Breman F C, VAN Beek M, Salim A, Sheil D. Wood density as a conservation tool: quantification of disturbance and identification of conservation-priority areas in tropical forests. Conservation Biology, 2008, 22(5): 1299-1308. DOI:10.1111/j.1523-1739.2008.00986.x |
| [42] |
Falster D S, Westoby M. Tradeoffs between height growth rate, stem persistence and maximum height among plant species in a post-fire succession. Oikos, 2005, 111(1): 57-66. DOI:10.1111/j.0030-1299.2005.13383.x |
| [43] |
Cheng X Q, Ping T, Li Z Z, Wang T, Han H R, Epstein H E. Effects of environmental factors on plant functional traits across different plant life forms in a temperate forest ecosystem. New Forests, 2022, 53(1): 125-142. DOI:10.1007/s11056-021-09847-0 |
| [44] |
Albert C H, de Bello F, Boulangeat I, Pellet G, Lavorel S, Thuiller W. On the importance of intraspecific variability for the quantification of functional diversity. Oikos, 2012, 121(1): 116-126. DOI:10.1111/j.1600-0706.2011.19672.x |
| [45] |
Poorter L, Markesteijn L. Seedling traits determine drought tolerance of tropical tree species. Biotropica, 2008, 40(3): 321-331. DOI:10.1111/j.1744-7429.2007.00380.x |
| [46] |
Rodríguez-Gallego C, Navarro T, Meerts P. A comparative study of leaf trait relationships in coastal dunes in southern Spain. Plant Ecology and Evolution, 2015, 148(1): 57-67. DOI:10.5091/plecevo.2015.951 |
| [47] |
He J S, Wang Z H, Wang X P, Schmid B, Zuo W Y, Zhou M, Zheng C Y, Wang M F, Fang J Y. A test of the generality of leaf trait relationships on the Tibetan Plateau. The New Phytologist, 2006, 170(4): 835-848. DOI:10.1111/j.1469-8137.2006.01704.x |
| [48] |
任一星. 北京山区典型森林群落功能多样性研究[D]. 北京: 北京林业大学, 2012.
|
| [49] |
张自浩. 中国东部典型森林功能多样性与植被生产力的关系[D]. 北京: 北京林业大学, 2020.
|
| [50] |
Zhang Z H, Hou J H, He N P. Predictability of functional diversity depends on the number of traits. Journal of Resources and Ecology, 2021, 12(3): 332-345. |
| [51] |
Mouchet M A, Villéger S, Mason N W H, Mouillot D. Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Functional Ecology, 2010, 24(4): 867-876. DOI:10.1111/j.1365-2435.2010.01695.x |
| [52] |
张国娟. 甘南高寒草甸海拔梯度上生物多样性与生态系统多功能的关系[D]. 兰州: 西北师范大学, 2022.
|
| [53] |
Magnago L F S, Edwards D P, Edwards F A, Magrach A, Martins S V, Laurance W F. Functional attributes change but functional richness is unchanged after fragmentation of Brazilian Atlantic forests. Journal of Ecology, 2014, 102(2): 475-485. DOI:10.1111/1365-2745.12206 |
| [54] |
姚兰. 湖北木林子保护区15hm2大样地森林群落结构及多样性[D]. 北京: 北京林业大学, 2016.
|
| [55] |
Grime J P. Trait convergence and trait divergence in herbaceous plant communities: mechanisms and consequences. Journal of Vegetation Science, 2006, 17(2): 255-260. DOI:10.1111/j.1654-1103.2006.tb02444.x |
| [56] |
赵家乐. 秦岭和黄土高原基于水力性状的辽东栎群落构建机制[D]. 西安: 西北大学, 2021.
|