 2024, Vol. 44
2024, Vol. 44文章信息
- 王云霞, 刘莹, 付雨辰, 李欣诺, 刘青, 马辉珍, 何俐蓉, 王国梁
- WANG Yunxia, LIU Ying, FU Yuchen, LI Xinnuo, LIU Qing, MA Huizhen, HE Lirong, WANG Guoliang
- 干旱胁迫对连翘幼苗非结构性碳分配和水力特性的影响
- Effects of drought on non-structural carbon allocation and hydraulic characteristics of Forsythia suspense seedlings
- 生态学报. 2024, 44(11): 4698-4707
- Acta Ecologica Sinica. 2024, 44(11): 4698-4707
- http://dx.doi.org/10.20103/j.stxb.202307081476
-
文章历史
- 收稿日期: 2023-07-08
- 网络出版日期: 2024-03-18






2. 中国科学院水利部水土保持研究所, 杨陵 712100;
3. 西北农林科技大学资源环境学院, 杨陵 712100;
4. 西北农林科技大学生命科学学院, 杨陵 712100
2. Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources, Yangling 712100, China;
3. College of Resources and Environment, Northwest Agriculture and Forestry University, Yangling 712100, China;
4. College of Life Sciences, Northwest Agriculture and Forestry University, Yangling 712100, China
干旱频率和持续时间的增加导致干旱和半干旱地区植物生产力下降, 甚至死亡[1]。干旱胁迫时, 植物木质部导管内聚力增大, 气体通过纹孔膜进入功能导管产生栓塞, 破坏导管输水的连续性[2]。植物通过调控气孔关闭以降低水分损失, 但同时减弱了光合作用, 限制了非结构性碳水化合物的产生和运输[3]。因此, 在干旱胁迫下维持碳平衡和水力运输是植物生长、存活以及适应环境的基础, 但目前关于植物如何调控NSC和水力特性的关系以应对长期干旱胁迫的机制尚不清楚。
非结构性碳水化合物(NSC)是植物初级和次级代谢的主要底物, 可溶性糖是碳水化合物运输和利用的主要形式, 淀粉是主要的能量储存物质[4]。干旱促进植物体淀粉转化为可溶性糖, 维持细胞膨压、抵御并适应干旱环境。植物在遭受干旱胁迫时, 调动储备的NSC资源可能成为生存的必要条件, 包括增加光合产物向地下组织的分配[5]、降低蒸腾速率[6]、提高渗透调节物质浓度进行栓塞修复进而提高水分运输效率[7]等。此外, NSC在器官间的分配主要受碳源、碳汇关系和同化物利用“就近原则”[8]的影响, 导致其浓度、转化比例在不同器官间存在差异。已有研究认为干旱期间树木NSC浓度表现为增加[9]、减少[10]或保持不变[11], 造成这些结果差异的原因可能是NSC浓度的变化规律在树种和器官间显著不同。因此, 迫切需要在长期干旱条件下探究不同器官NSC及组分的积累和分配。
树木的水力特性与NSC之间密切相关。一方面, 栓塞会导致气孔关闭, 同时限制光合作用, 减少碳同化产物[12];另一方面, 植物在响应干旱胁迫时, 栓塞的产生会改变淀粉和可溶性糖的代谢和分配[13]。可溶性糖作为渗透调节物质被运输到栓塞导管用于改变其两侧的渗透势, 影响着木质部在旱后的水力恢复[14]。可见, 栓塞程度与可溶性糖含量密切相关[15]。已有研究认为, 干旱胁迫后植物的栓塞程度与可溶性糖的关系为正相关[16]、负相关[17]或者不相关[18]。造成这种差异的原因可能是淀粉降解为多种可溶性糖组分, 如蔗糖、葡萄糖、麦芽糖、果糖等。但这些糖分对水分输运的调节作用不同, 如蔗糖可以作为栓塞信号在木质部导管的保护和再填充机制中起到关键作用[19], 葡萄糖和果糖是主要的渗透调节物质[20]。由此可见, 可溶性糖各组分含量可能受到木质部水分状态和植物体内多种生理过程的影响。因此, 了解可溶性糖组分的变化对探究NSC与水力特性的关系至关重要。
连翘(Forsythia suspensa)是黄土高原水土保持和绿化的重要树种, 根系发达、生长迅速[21]。本文以2年生连翘幼苗为研究对象, 经过连续3年不同水分条件处理(适宜供水CK、中度干旱胁迫MS、重度干旱胁迫SS), 探究植物不同器官的非结构碳水化合物组分含量, 水力特性的变化及其两者之间的相关关系, 为半干旱区连翘培育和经营提供理论依据。
1 研究地区与研究方法 1.1 研究区概况研究区位于陕西杨陵西北农林科技大学水土保持研究所, 地理位置为34°16′N, 108°04′E、该区海拔为530 m, 属于暖温带半干旱区, 典型大陆性季风气候, 昼夜温差较大, 四季分明。最热为7月份, 平均气温为26.2℃, 最冷为1月份, 平均气温为-1.4℃。年总日照为2163.8 h, 年降雨量大约650 mm, 雨量分布不均匀。
1.2 实验设计2018年2月, 在杨陵苗圃地选取两年生的连翘幼苗作为实验材料, 将生长状况良好且根系完整的幼苗移植至花盆中, 花盆上口径、下口径和高分别为35、32、27cm, 每盆装有15 kg的黄绵土。经测定土壤全碳、全氮和全磷分别是6.746、0.493、0.48g/kg, pH为8.79。移栽后进行正常灌溉, 缓苗3个月, 保证幼苗正常生长。
本实验设置三个土壤水分梯度:适宜供水CK、中度干旱胁迫MS、重度干旱胁迫SS, 土壤含水率分别为13.8%、10.1%和6.4%, 即田间持水量的75%±5%, 55%±5%和35%±5%。每个水分梯度选取6盆, 共计18盆。于2018年5月份开始长期控水实验, 使用旱棚遮雨的方式来防止雨水和其他水分干扰, 每3—4 d补充水分, 控水时间统一为下午19:00, 通过称重法进行干旱控水处理, 控水周期3年为2018年5月—2021年5月。在干旱控水期间对连翘幼苗进行健康监测, 确保幼苗正常生长。
1.3 测定项目与方法 1.3.1 连翘光合特性和瞬时水分利用效率的测定于2021年6月1日进行取样, 采用Li-6800光合仪在晴朗无云的9:00—11:00测定自然条件下光合特性。每个处理随机选取3盆, 每个叶片重复测定3次, 等待叶片稳定2分钟后测定净光合速率、蒸腾速率和气孔导度, 并计算瞬时水分利用效率(WUE):

|
其中, Pn为净光合速率(umol m-2 s-1); Tr为蒸腾速率(umol m-2 s-1)。
1.3.2 连翘枝条水力特性的测定导水率、栓塞程度(PLC)的测定:于光合测定当日6:00, 选取3盆幼苗装在黑色的塑料袋中立即送回实验室, 将融化的石蜡通过注射器注入连翘髓心内并确保髓心处于密封状态。随后将枝条浸没在蒸馏水中, 在水下切割直径为6 mm, 长度为5 cm的5个连续的小段。将切好小段连接到低压液流计(LPFM)上, 使用冲洗液为10 mmol/L的KCL溶液在5 KPa水压下获取枝条初始导水率Kh, 100 KPa水压冲洗后测量枝条的最大导水率Kmax。枝条的栓塞程度用导水率损失百分数来表示[22]:

|
将剩余幼苗地上组织刈割为叶、枝、茎, 根系按根序法分为1级根、2级根、3级根、4级根、5级根和6级根, 将前3级根作为细根, 4、5、6级根作为粗根, 逐根手动挑出, 防止根系损失, 称取鲜重。随后将所有样本在烘箱内杀青、烘干至恒重后取出, 称量各器官的生物量以及计算组织含水量和生物量比∶叶茎比、根叶比、根冠比(地下部分总和/地上部分总和)和细根粗根比。烘干样品后使用球磨仪粉碎, 过0.25 mm筛后用于测定NSC组分含量。
1.3.4 非结构性碳水化合物及其组分的测定可溶性总糖:将约0.1 g样品置于10 mL离心管中, 加入5 mL的80%乙醇溶液。混合后, 将离心管置于80 ℃的水浴中30 min, 冷却至室温, 并以3500 r/min离心10 min, 将上清液收集在25 mL容量瓶中。再重复上述步骤两次, 将上清液收集在容量瓶后定容, 即为可溶性糖提取液。取2 mL后, 使用蒽酮-硫酸法在620 nm处测量吸光度, 计算可溶性总糖的浓度。
淀粉:向沉淀物中加入2 mL蒸馏水, 在沸水中糊化15 min。冷却后, 加入2 mL9.2 mol/L的HClO4, 震荡15 min, 加入4mL蒸馏水, 将样品在3500 r/min离心10 min, 提取上清液。再将2 mL 4.6 mol/L的HClO4加入到沉淀物, 震荡15 min, 加入5 mL蒸馏水, 离心提取上清液, 将两次上清液收集于50 mL容量瓶后定容后。取2 mL, 使用蒽酮-硫酸法在620 nm处测量吸光度, 计算可溶性糖和淀粉删除,加上计算如下公式[23]:
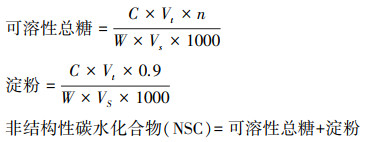
|
其中, C为标准曲线对应的读取吸光度的葡萄糖含量(μg);1000为转化因子, 1 mg=1000 μg;n为稀释比;Vt为样品提取物的总体积(mL);Vs为测定取样体积(mL);W为样品的干重(g);0.9为葡萄糖转化为淀粉的转化系数。
可溶性糖组分:取1 mL可溶性糖提取液, 使用高效液相色谱仪(HPLC)测定, 其配有RID-10示差检测器, 流速为0.4 mL/min, 温度为70 ℃, 流动相为纯水, 通过比较色谱图的峰高与标准溶液获得曲线来计算出可溶性糖组分各浓度, 主要有葡萄糖、蔗糖、麦芽糖[24]。
1.4 数据处理利用SPSS 26.0、Orgin 2018进行统计分析数据。用单因素方差分析(One-way ANOVE)检验干旱程度对不同器官的光合作用、生物量、淀粉和可溶性总糖及组分的差异。用线性回归和Pearson相关分析确定栓塞程度与可溶性总糖、淀粉、蔗糖和葡萄糖含量的相关性。分析的置信度标准为95%(P<0.05)。
2 结果与分析 2.1 连翘幼苗光合特性对干旱的响应连翘叶片的净光合速率、蒸腾速率、气孔导度、水分利用效率均随着干旱程度的增加而显著降低。其中, 在重度胁迫SS处理下, 净光合速率、蒸腾速率、气孔导度和水分利用效率值比CK处理降低了81.2%、54.7%、59.7%和18.3%(表 1)。
| 水分处理 Level of water stress |
净光合速率 Net photosynthetic rate/ (μmol m-2 s-1) |
蒸腾速率 Transpiration rate/ (mol m-2 s-1) |
气孔导度 Stomatal conductance/ (mol m-2 s-1) |
水分利用效率 Water use efficiency/ (mmol/mol) |
| 适应供水Appropriate water supply | 3.918±0.204 a | 1.270±0.083 a | 0.033±0.002 a | 3.109±0.108 a |
| 中度胁迫Moderate stress | 3.066±0.373 b | 0.992±0.152 ab | 0.026±0.004 ab | 3.160±0.093 a |
| 重度胁迫Severe stress | 2.161±0.100 c | 0.821±0.009 b | 0.021±0.000 b | 2.628±0.094 b |
| 不同小写字母表示不同水分处理间差异显著(P<0.05) | ||||
随着干旱程度的增加, 连翘地上部分和粗根生物量显著降低, 但细根生物量显著增加。SS处理下连翘叶、枝和茎生物量分别比CK处理降低了64.5%、50.9%和46.7%, 而SS处理下细根生物量比CK处理增加了32.3%。随着干旱程度的增加, 连翘的根冠比、细根粗根比和根叶比显著增加。SS处理下的根叶比、细根粗根比分别比CK处理增加了70.4%、127.3%, 而根冠比分别是MS和CK处理的1.2和1.5倍(图 1)。

|
| 图 1 不同水分条件下连翘不同器官的生物量及其比值 Fig. 1 The biomass and its ratio of different organs of Forsythia suspensa under different water stress CK:对照(适宜供水);MS:中度干旱胁迫;SS:重度干旱胁迫;小写字母表示不同水分处理间差异显著(P<0.05) |
随着干旱程度的增加, 连翘枝条栓塞程度显著增加, 而枝条导水率显著降低。CK、MS、SS处理下枝条栓塞程度分别为30.7%、41.8%和42.3%, 枝条导水率分别为0.95、0.71、0.65 kg m-1 s-1 MPa-1。随着干旱程度的增加, 连翘幼苗各器官的组织含水量显著降低。SS处理下幼苗叶片、枝条和粗根的组织含水量分别比CK处理降低了7.7%、24.2%和42.3%(图 2)。

|
| 图 2 不同水分条件下连翘幼苗枝条的栓塞程度和导水率以及不同器官组织含水量的影响 Fig. 2 The degree of embolism and hydraulic conductivity of Forsythia suspensa seeding branches, as well as the influence of water content in various organs under different water stress 不同小写字母表示导水率在不同水分处理间和组织含水量在器官间的显著差异, 不同大写字母表示栓塞程度在不同水分处理间的显著差异(P<0.05) |
随着干旱程度的增加, 连翘除叶片外, 各器官的NSC含量显著降低。SS处理下连翘枝条、茎、细根和粗根的NSC含量分别比CK处理降低了6.7%、10.7%、12.5%和16.1%。随着干旱程度的增加, 连翘叶片和枝条的可溶性总糖显著增加, 淀粉含量显著降低;而根系的可溶性总糖显著降低, 淀粉含量先增加后降低。SS处理下叶片、枝条的可溶性总糖比CK处理增加了21.2%、12.9%, 淀粉比CK处理降低了24.5%、40.7%;细根和粗根的可溶性总糖比CK处理降低了18.8%和15.7%(图 3)。此外, 枝条栓塞程度与可溶性总糖显著正相关, 而栓塞程度与淀粉含量为显著负相关(P<0.01, 图 4)。

|
| 图 3 不同水分条件下连翘不同器官非结构碳水化合物的影响 Fig. 3 The contents of non-structural carbohydrate in various organs of Forsythia suspensa under different water stress 不同小写字母表示非结构性碳水化合物(NSC)、可溶性总糖、淀粉、葡萄糖、蔗糖和麦芽糖在器官间显著差异, 不同大写字母表示NSC、可溶性总糖、淀粉、葡萄糖、蔗糖和麦芽糖在不同水分处理间显著差异(P<0.05) |

|
| 图 4 连翘幼苗枝条可溶性糖组分含量与栓塞程度和导水率线性相关分析 Fig. 4 Linear correlation analysis of soluble sugar component content and hydraulic characteristics in branches of Forsythia suspensa seedlings 图中灰色阴影表示95%的置信区间 |
随着干旱程度的增加, 叶片和枝条的葡萄糖、蔗糖含量显著增加;粗根的葡萄糖、蔗糖含量整体上显著降低。SS处理下, 连翘枝条的葡萄糖、蔗糖含量分别比CK处理增加了31.1%、45.7%, 粗根的葡萄糖、蔗糖含量分别比CK处理降低了28.8%、25.9%(图 3)。此外, 枝条的水力特性与糖组分显著相关。其中, 枝条栓塞程度与葡萄糖、蔗糖显著正相关, 而导水率与葡萄糖、蔗糖显著负相关(P<0.01, 图 4)。
3 讨论 3.1 长期干旱胁迫对连翘水分状况和光合作用的影响植物通过调整水力特性适应干旱环境[25]。本研究中, 长期干旱胁迫导致枝条组织含水量和导水率显著下降, 而栓塞程度显著增加(图 2)。有大量研究已经发现, 植物在干旱环境下水力功能受到抑制, 木质部的导水能力降低[26—27]。造成这种变化的原因可能是木质部解剖结构的变化, 如导管直径[28]、导管壁厚度[29]等。以往研究已经证实, 木质部导管数量和大小决定着植物的输水效率[30], 导管壁的厚度影响水力安全[31]。因此, 本研究推测, 导管结构的变化可能是造成水分运输效率和抗栓塞能力变化的主要因素。然而, 本研究只测定了枝条的水力特性, 未涉及解剖结构的实验, 不能为这一推测提供直接的证据, 后续还需补充关于连翘枝条解剖结构的研究来证实本研究的推测。
长期干旱不仅影响植物的水力特性, 还限制了光合作用。本研究中, 重度干旱胁迫导致叶片的光合速率、蒸腾速率和水分利用效率显著降低(表 1)。与已有研究结果[32—33]一致, 造成光合作用下降原因可能是气孔调控[34]。因此, 长期干旱胁迫改变了连翘的水力特性, 增加了枝条栓塞程度, 降低了木质部水分运输效率(图 2), 同时植物为了避免水分散失调控气孔关闭, 也降低植物的光合速率和蒸腾速率(表 1), 从而防止植物蒸腾失水, 维持水分平衡。
3.2 长期干旱胁迫对连翘生长和NSC含量的影响长期干旱胁迫导致连翘地上生物量和NSC含量显著下降。本研究中, 长期重度干旱胁迫后, 地上部分和粗根的生物量显著降低, 但细根生物量和根冠比显著增加(图 1)。造成这种变化的原因是植物通过增加地下生物量来提高根系的吸水能力, 减少叶片、茎的生物量来降低蒸腾损耗[35]。此外, 本研究发现长期干旱胁迫后除叶片外, 各器官NSC含量显著降低(图 3)。与已有的研究结果[36—37]一致, 可能是由于长期干旱胁迫导致光合获取能力受到抑制, 光合产物降低。其次, 干旱环境中植物为了维持代谢活动会大量消耗各器官的碳水化合物, 导致NSC储备显著降低。本文结果发现在SS处理下根系NSC消耗量最高, 根系的可溶性总糖和淀粉含量显著降低, 导致根系发生碳亏损的可能性更高(图 3)。关于刺槐和山毛榉的研究也发现在干旱环境下根系NSC消耗量较高[38—39], 可能是由于重度干旱胁迫降低了有机物从叶片通过韧皮部向根系的转运, 导致根系更依赖木质部储存的NSC[40]。因此, 本研究认为, 连翘通过调节不同器官的碳分配模式以应对长期干旱胁迫。
3.3 枝条栓塞程度与可溶性糖及组分含量的关系枝条水力功能降低对可溶性总糖及组分含量具有显著影响。本研究中, 枝条栓塞程度与可溶性总糖、淀粉、蔗糖和葡萄糖含量存在显著相关性(图 4)。可溶性糖和淀粉是NSC的主要组成部分, 对水分输运具有重要的调节作用[41]。当植物在干旱期间遭受的生理损伤较为严重时, 复水后就需要消耗更多的可溶性糖储备用于生理功能和水力功能的恢复[17]。其次, 淀粉作为主要的储存物质, 当栓塞发生后能够转化为可溶性糖以满足植物对碳的需求[42]。此外, 已有研究表明, 可溶性糖、葡萄糖和蔗糖是重要的渗透调节物质[43]。当木质部导管发生空穴化, 淀粉水解速率增加和蔗糖转运基因的表达促进了二糖(麦芽糖和蔗糖)外排到质外体, 导致木质部pH值下降[44], pH通过调控酸性转化酶活性诱导质外体蔗糖分解成葡萄糖和果糖, 导致单糖的积累[45]。结合本研究结果认为长期干旱胁迫下可溶性总糖、淀粉、葡萄糖和蔗糖含量的变化可能受到木质部水分状态和植物体内多种生理过程的调控。因此, 本研究认为在长期干旱胁迫下连翘通过协调NSC在不同器官间的分配及其各组分的含量以提高植物在干旱条件下的存活机会及旱后水分恢复能力。
4 结论本研究表明, 长期干旱胁迫增加了枝条栓塞程度, 降低了木质部的导水能力, 同时也降低了植物的光合作用和水分运输效率。其次, 长期干旱胁迫导致地上生物量和各器官的NSC含量均显著降低。此外, 栓塞程度与可溶性总糖、淀粉、蔗糖和葡萄糖含量显著相关。因此, 本研究认为连翘通过调节器官间碳分配模式及NSC各组分含量来应对长期重度干旱胁迫。本研究通过探究长期干旱胁迫下连翘幼苗不同器官的非结构性碳水化合物组分含量、水力特性的变化及其两者之间的相关关系, 为半干旱区连翘培育和经营提供理论依据。
| [1] |
Park Williams A, Allen C D, Macalady A K, Griffin D, Woodhouse C A, Meko D M, Swetnam T W, Rauscher S A, Seager R, Grissino-Mayer H D, Dean J S, Cook E R, Gangodagamage C, Cai M, McDowell N G. Temperature as a potent driver of regional forest drought stress and tree mortality. Nature Climate Change, 2013, 3(3): 292-297. DOI:10.1038/nclimate1693 |
| [2] |
金鹰, 王传宽, 周正虎. 木本植物木质部栓塞修复机制: 研究进展与问题. 植物生态学报, 2016, 40(8): 834-846. |
| [3] |
Anderegg W R L, Anderegg L D L, Berry J A, Field C B. Loss of whole-tree hydraulic conductance during severe drought and multi-year forest die-off. Oecologia, 2014, 175(1): 11-23. DOI:10.1007/s00442-013-2875-5 |
| [4] |
王雲霞, 单立山, 解婷婷, 马静, 师亚婷. 干旱-复水对红砂幼苗各器官非结构性碳水化合物的影响. 生态学杂志, 2023, 43(1): 1-14. |
| [5] |
Liu Y, Li P, Xiao L, Wang W, Yu K X, Shi P. Heterogeneity in short-term allocation of carbon to roots of Pinus tabuliformis seedlings and root respiration under drought stress. Plant and Soil, 2020, 452(1): 359-378. |
| [6] |
邵畅畅, 罗仙英, 丁贵杰, 段洪浪, 赵熙州, 娄清. 干旱对马尾松茎叶水力特征及解剖特性的影响. 植物生理学报, 2022, 58(5): 937-945. |
| [7] |
Yu Y C, Liu J X, Sun Z Y. Transcriptome profiling in Salix matsudana during refilling xylem vessels after embolism. Biologia Plantarum, 2019, 63: 425-431. |
| [8] |
Dietze M C, Sala A, Carbone M S, Czimczik C I, Mantooth J A, Richardson A D, Vargas R. Nonstructural Carbon in Woody Plants. Annual Review of Plant Biology, 2014, 65(1): 667-687. DOI:10.1146/annurev-arplant-050213-040054 |
| [9] |
O'Brien M J, Burslem D F R P, Caduff A, Tay J, Hector A. Contrasting nonstructural carbohydrate dynamics of tropical tree seedlings under water deficit and variability. The New Phytologist, 2015, 205(3): 1083-1094. DOI:10.1111/nph.13134 |
| [10] |
Woodruff D R. The impacts of water stress on phloem transport in Douglas-fir trees. Tree Physiology, 2014, 34(1): 5-14. DOI:10.1093/treephys/tpt106 |
| [11] |
Anderegg W R L, Berry J A, Smith D D, Sperry J S, Anderegg L D L, Field C B. The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(1): 233-237. |
| [12] |
Martorell S, Diaz-Espejo A, Medrano H, Ball M C, Choat B. Rapid hydraulic recovery in Eucalyptus pauciflora after drought: linkages between stem hydraulics and leaf gas exchange. Plant, Cell & Environment, 2014, 37(3): 617-626. |
| [13] |
Fang L D, Ning Q R, Guo J J, Gong X W, Zhu J J, Hao G Y. Hydraulic limitation underlies the dieback of Populus pseudo-simonii trees in water-limited areas of Northern China. Forest Ecology and Management, 2021, 483: 118764. DOI:10.1016/j.foreco.2020.118764 |
| [14] |
Choat B, Brodribb T J, Brodersen C R, Duursma R A, López R, Medlyn B E. Triggers of tree mortality under drought. Nature, 2018, 558(7711): 531-539. DOI:10.1038/s41586-018-0240-x |
| [15] |
Salleo S, Trifilò P, Esposito S, Nardini A, Lo Gullo M A. Starch-to-sugar conversion in wood parenchyma of field-growing Laurus nobilis plants: a component of the signal pathway for embolism repair?. Functional Plant Biology: FPB, 2009, 36(9): 815-825. DOI:10.1071/FP09103 |
| [16] |
Wang A Y, Han S J, Zhang J H, Wang M, Yin X H, Fang L D, Yang D, Hao G Y. The interaction between nonstructural carbohydrate reserves and xylem hydraulics in Korean pine trees across an altitudinal gradient. Tree Physiology, 2018, 38(12): 1792-1804. DOI:10.1093/treephys/tpy119 |
| [17] |
Tomasella M, Petrussa E, Petruzzellis F, Nardini A, Casolo V. The possible role of non-structural carbohydrates in the regulation of tree hydraulics. International Journal of Molecular Sciences, 2019, 21(1): 144. DOI:10.3390/ijms21010144 |
| [18] |
Tomasella M, Häberle K H, Nardini A, Hesse B, Machlet A, Matyssek R. Post-drought hydraulic recovery is accompanied by non-structural carbohydrate depletion in the stem wood of Norway spruce saplings. Scientific Reports, 2017, 7(1): 14308. DOI:10.1038/s41598-017-14645-w |
| [19] |
Ennajeh M, Ehwald R, Kühn C. Role of sucrose and phloem-xylem interaction in recovery of water status and hydraulic dehydration impacts in tobacco plants (Nicotiana tabacum). Acta Physiologiae Plantarum, 2022, 44(5): 56. DOI:10.1007/s11738-022-03390-x |
| [20] |
Zhong Y L, Xie J Y, Wen S Z, Wu W W, Tan L, Lei M G, Shi H Z, Zhu J K. TPST is involved in fructose regulation of primary root growth in Arabidopsis thaliana. Plant Molecular Biology, 2020, 103(4): 511-525. |
| [21] |
王阿丽, 任子蓓, 金蒙蒙, 任士福. 不同连翘属植物抗旱性比较. 林业科技开发, 2015, 29(3): 52-55. |
| [22] |
王林, 代永欣, 樊兴路, 张芸香, 黄平, 万贤崇. 风对黄花蒿水力学性状和生长的影响. 生态学报, 2015, 35(13): 4454-4461. DOI:10.5846/stxb201410071963 |
| [23] |
Hoch G, Richter A, Körner C. Non-structural carbon compounds in temperate forest trees. Plant, Cell & Environment, 2003, 26(7): 1067-1081. |
| [24] |
胡晓健, 喻方圆, 刘建兵, 万劲. 干旱胁迫对不同种源马尾松苗木针叶内可溶性糖含量的影响. 南京林业大学学报: 自然科学版, 2009, 33(5): 55-59. |
| [25] |
Słupianek A, Dolzblasz A, Sokołowska K. Xylem parenchyma-role and relevance in wood functioning in trees. Plants, 2021, 10(6): 1247. DOI:10.3390/plants10061247 |
| [26] |
龚容, 徐霞, 田晓宇, 江红蕾, 李霞, 关梦茜. 三种锦鸡儿属植物水力结构特征及其干旱适应策略. 生态学报, 2018, 38(14): 4984-4993. DOI:10.5846/stxb201706201123 |
| [27] |
陈志成, 陆海波, 刘世荣, 刘晓静, 刘畅, 万贤崇. 锐齿栎水力结构和生长对降雨减少的响应. 生态学报, 2018, 38(7): 2405-2413. |
| [28] |
Hacke U G, Spicer R, Schreiber S G, Plavcová L. An ecophysiological and developmental perspective on variation in vessel diameter. Plant, Cell & Environment, 2017, 40(6): 831-845. |
| [29] |
Pittermann J, Sperry J S, Wheeler J K, Hacke U G, Sikkema E H. Mechanical reinforcement of tracheids compromises the hydraulic efficiency of conifer xylem. Plant, Cell & Environment, 2006, 29(8): 1618-1628. |
| [30] |
Islam M, Rahman M, Bräuning A. Long-term hydraulic adjustment of three tropical moist forest tree species to changing climate. Frontiers in Plant Science, 2018, 9: 1761. DOI:10.3389/fpls.2018.01761 |
| [31] |
Guérin M, von Arx G, Martin-Benito D, Andreu-Hayles L, Griffin K L, McDowell N G, Pockman W, Gentine P. Distinct xylem responses to acute vs prolonged drought in pine trees. Tree Physiology, 2020, 40(5): 605-620. DOI:10.1093/treephys/tpz144 |
| [32] |
魏清江, 冯芳芳, 马张正, 苏受婷, 宁少君, 辜青青. 干旱复水对柑橘幼苗叶片光合、叶绿素荧光和根系构型的影响. 应用生态学报, 2018, 29(8): 2485-2492. |
| [33] |
施梦娇, 李斌, 伊力塔, 刘美华. 美洲黑杨幼苗生长和生理生态指标对干旱-复水响应的性别差异. 植物生态学报, 2023, 47(8): 1159-1170. |
| [34] |
高冠龙, 冯起, 张小由, 司建华, 鱼腾飞. 植物叶片光合作用的气孔与非气孔限制研究综述. 干旱区研究, 2018, 35(4): 929-937. |
| [35] |
王鑫, 余新晓, 贾国栋, 邱云霄, 史佳美, 孜尔蝶. 不同土壤水分条件下侧柏幼苗的生理活动及氮素分配策略. 水土保持学报, 2020, 34(3): 311-317. |
| [36] |
翟培凤, 关家欣, 何鹏, 刘贺永, 满良, 姜勇, 马成仓. 沿干旱梯度樟子松人工林针叶和枝条非结构性碳水化合物及氮含量的变化. 应用生态学报, 2022, 33(6): 1518-1524. |
| [37] |
Yan W M, Zhong Y Q W, Shangguan Z P. Rapid response of the carbon balance strategy in Robinia pseudoacacia and Amorpha fruticosa to recurrent drought. Environmental and Experimental Botany, 2017, 138: 46-56. DOI:10.1016/j.envexpbot.2017.03.009 |
| [38] |
Yan W M, Zhong Y Q W, Shangguan Z P. Elevated temperature exacerbates the effects of drought on the carbon and hydraulic characteristics of Robinia pseudoacacia seedlings. Agricultural and Forest Meteorology, 2020, 280(C): 107794. |
| [39] |
Ruehr N K, Offermann C A, Gessler A, Winkler J B, Ferrio J P, Buchmann N, Barnard R L. Drought effects on allocation of recent carbon: from beech leaves to soil CO2 efflux. The New Phytologist, 2009, 184(4): 950-961. DOI:10.1111/j.1469-8137.2009.03044.x |
| [40] |
Hartmann H, Ziegler W, Trumbore S. Lethal drought leads to reduction in nonstructural carbohydrates in Norway spruce tree roots but not in the canopy. Functional Ecology, 2013, 27(2): 413-427. DOI:10.1111/1365-2435.12046 |
| [41] |
郑悦, 王爱英, 苏立新, 郭晶晶, 段春旸, 殷笑寒, 龚雪伟, 郝广友. 沈阳市区不同环境下银杏水力特征和非结构性碳水化合物含量. 应用生态学报, 2022, 33(3): 711-719. |
| [42] |
Tomasella M, Nardini A, Hesse B D, Machlet A, Matyssek R, Häberle K H. Close to the edge: effects of repeated severe drought on stem hydraulics and non-structural carbohydrates in European beech saplings. Tree Physiology, 2019, 39(5): 717-728. |
| [43] |
Huang X Y, Guo W J, Yang L, Zou Z G, Zhang X Y, Addo-Danso S D, Zhou L L, Li S B. Effects of drought stress on non-structural carbohydrates in different organs of Cunninghamia lanceolata. Plants, 2023, 12(13): 2477. DOI:10.3390/plants12132477 |
| [44] |
Pagliarani C, Casolo V, Ashofteh Beiragi M, Cavalletto S, Siciliano I, Schubert A, Gullino M L, Zwieniecki M A, Secchi F. Priming xylem for stress recovery depends on coordinated activity of sugar metabolic pathways and changes in xylem sap pH. Plant, Cell & Environment, 2019, 42(6): 1775-1787. |
| [45] |
Secchi F, Zwieniecki M A. Accumulation of sugars in the xylem apoplast observed under water stress conditions is controlled by xylem pH. Plant, Cell & Environment, 2016, 39(11): 2350-2360. |