 2024, Vol. 44
2024, Vol. 44文章信息
- 陈子川, 潘国营, 陈灿, 徐云鹏, 林晗, 陈煜, 谢安强, 范海兰
- CHEN Zichuan, PAN Guoying, CHEN Can, XU Yunpeng, LIN Han, CHEN Yu, XIE Anqiang, FAN Hailan
- 光强对木麻黄幼苗根系形态、解剖结构及其碳氮含量的影响
- Effects of light intensity on root morphology, anatomical structure and carbon and nitrogen content of Casuarina equisetifolia seedlings
- 生态学报. 2024, 44(10): 4377-4387
- Acta Ecologica Sinica. 2024, 44(10): 4377-4387
- http://dx.doi.org/10.20103/j.stxb.202307231573
-
文章历史
- 收稿日期: 2023-07-23
- 网络出版日期: 2024-02-28






2. 森林生态系统过程与经营福建省高校重点实验室, 福州 350002;
3. 晋江坫头国有防护林场, 晋江 362200;
4. 福建农林大学生命科学学院, 福州 350002
2. Key Lab for Forest Ecosystem Processes and Management in Fujian Province, Fuzhou 350002, China;
3. Jinjiang Goblet Head State-owned Protective Forestry Farm, Jinjiang 362200, China;
4. College of Life Sciences, Fujian Agriculture and Forestry University, Fuzhou 350002, China
光照是植物生命活动中关键且必要环境因子, 是植物光合作用、合成有机质和获取能量的根本来源[1]。森林中光照具有较高异质性, 对林下植被可塑性、自然生长和更新产生深远的影响[2]。自然光通过植物光感受器传递信号分子改变植物生理条件和形态, 体现为植物的地上和地下部分均有较高的可塑性[3—4]。其中, 根系外部形态结构是研究植物根系重要的表型指标, 也是生长发育水平和对外界环境适应的直接体现, 其差异将直接影响植物对土壤水分和养分吸收能力[5]。研究表明光通过影响植物的根系的生长, 导致根、茎间通信及信号分子分布调整, 引起侧根系外部形态改变, 降低有效养分吸收, 并对植物应对胁迫刺激的适应能力造成负面影响[4, 6—7], 但与茎、枝和叶的研究间形成鲜明对比的是根分布在地下, 错综复杂, 研究难度大, 导致植物地上和地下研究不平衡。这种知识差距阻碍陆地生物圈模型中根系过程的表达, 并限制未来预测生态系统对变化环境反应的能力[8]。
为解决上述研究困难, 乔木幼苗是较为理想研究材料, 细根是根系研究重要内容和切入点。乔木幼苗与成熟个体相比便于挖掘且保留完整形态结构;细根是植物光合产物和资源的重要投资。每年全球陆地生态系统中22%以上的净初级生产力通过细根系周转转移到地下并在水、营养物质和碳的循环中起着重要作用[9]。细根研究通常采用单一阈值法[10]和根序法[11]。前者通常将直径≤2 mm的根系划为细根, 而直径>2 mm划为粗根, 但有学者认为单一直径阈值的方法忽略了个体根在根系分支上位置的不同, 并影响反映生理生态规律的准确性[10];后者基于根序的划分能更充分判定细根系统内结构和功能的异质性, 表现出不同的生理和形态特征。此外, 根系的解剖学特性与外部形态和生理生态功能密切相关, 例如皮层与中柱的结构反映植株吸收和运输的两种功能[9]。当前基于不同根序解剖结构异质性受到广泛关注和重视, 对特殊生境或用途的乔木根系的研究将有助于森林资源更好的开发和经营管理。
木麻黄(Casuarina equisetifolia)阳性树种, 具喜光、耐旱、固风沙等特性, 广泛种植于世界热带和亚热带地区[12—13], 也是我国东南沿海漫长海岸线防护林主要树种, 近年二代更新困难等问题日益突出。防护林密植有利于防风效果, 但过密的林分结构不仅会影响林下幼苗的光照并间接改变温度、水分等微环境, 还导致林下幼苗生长受限[14—15]。环境条件塑造植物的整个结构水平和单个器官水平, 但现今木麻黄对光照响应研究着重于育种[16—17]、共生菌[18]、光合特性[19—20]等方面影响, 而根系结构的响应变化亦是其获取生存资源保证存活和正常生长的重要原因[21—22]。目前, 关于木麻黄根系研究主要涉及到根系微生物, 包括内生真菌、细菌等[23]对根系生理活性[24]、根系形态特征[25]等方面的影响, 而乔木类根系内部解剖结构对光照的响应机制尚未见报道。自然条件下, 光强变化在多大程度上影响乔木幼苗根系内外结构、组织化学含量以及生长策略?为验证和量化此问题本研究以1年生木麻黄苗木为试供材料, 模拟林分内外光环境设置4种遮光处理, 结合根径和根序分级两种方法分析光限制下根系外部形态结构、内部解剖结构及养分变化等指标变化特征, 以期为沿海防护林的可持续经营提供参考。
1 材料与方法 1.1 实验地点幼苗试验地位于北纬26°05′, 东经119°13′的福建省福州市福建农林大学田间科技园内, 当地属于亚热带季风气候, 地势平坦, 无霜期达326 d, 年平均日照数为1700—1980 h, 年平均降水量为900—2100 mm, 年相对湿度约77%。
1.2 实验材料及设计选择长势良好且一致的1年生木麻黄幼苗(选自福建省惠安赤湖国有防护林场)种植到花盆中, 1盆1株, 综合前期野外调查结果, 每盆装土(纯土∶粗沙=1∶3)10 kg, 日常管理缓苗一月后测定植株基本数据, 平均株高36.6 cm, 平均基径4.23 mm。实验用土pH为6.5, 含水量14.62%, 电导率29.89 s/m, 速效氮4.59 mg/kg, 速效磷1.64 mg/kg, 速效钾1.13 mg/kg。搭建遮阳棚使用不同针数的黑色尼龙网模拟森林幼苗生长的林外(CK)、林缘(L1)、林窗(L2)和林下(L3)光环境设置3个遮荫光照强度, 其值分别约为自然全光照的55%、30%和15%(表 1)[26], 于晴天采用TES1339光照度计测量光照强度, 分别为2700 Lux、1500 Lux、810 Lux和405 Lux, 以透光率=遮荫下的数值/无遮荫下的数值换算透光率[27], 每个处理10个重复。
| 处理 Treatment |
光照强度/% Light intensity |
株高增量/cm Relative growth of plant height |
地径增量/mm Relative growth of ground diameter |
叶干重/g Leaf biomass |
茎干重/g Stem biomass |
根干重/g Root biomass |
地上部分干重/g Aboveground biomass |
根冠比 Root-shoot ratio |
| CK | 100 | 8.98±0.92a | 0.77±0.13a | 1.38±0.16a | 1.00 ±0.08a | 1.08±0.15a | 2.38±0.24a | 0.47±0.09a |
| L1 | 55 | 10.18±1.01a | 0.12±0.03b | 1.0±0.15ab | 1.05±0.25a | 1.30±0.33a | 2.15±0.40ab | 0.60±0.11a |
| L2 | 30 | 4.80±1.94b | 0.11±0.0.3 b | 0.72±0.20b | 0.69±0.15a | 0.77±0.23a | 1.41±0.32ab | 0.53±0.08a |
| L3 | 15 | 1.12±0.45b | 0.09±0.05b | 0.59±0.04b | 0.70±0.05a | 0.66±0.17a | 1.29±0.09b | 0.50±0.11a |
| 表中数值为平均值±标准误差, 同一列不同的小写字母表示不同处理之间的差异(P < 0.05);CK:100%自然全光照100% natural full light;L1:55%自然全光照55% natural full light;L2:30%自然全光照30% natural full light;L3:15%自然全光照15% natural full light | ||||||||
经4个月生长期后每个处理选取6棵, 测定株高、地径, 并将根茎叶称取鲜重, 105℃杀青后80℃恒温烘干后测定干重。根系采用根系扫描仪(Expression 11000XL)描后用WinRHIZO(Pro 2009c)根系图像分析仪得到根系形态指标参数:根长(Len)、表面积(RA)、体积(Vol)、根尖数(T)、根平均直径(Avg D)以及4个不同根系径级Ⅰ级(d≤0.5 mm)、Ⅱ级(0.5<d≤2 mm)、Ⅲ级(2 <d≤ 5 mm)和Ⅳ级(d>5 mm)内的根长、根表面积、根体积和根尖数[28—29], 并计算比根长等指标[30], 公式如下:
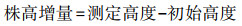
|
(1) |
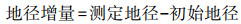
|
(2) |
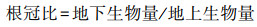
|
(3) |

|
(4) |
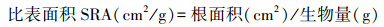
|
(5) |
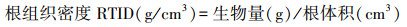
|
(6) |
各处理选取3盆植株, 采用根序法对根取样, 按照最末段的为1级根, 1级根的母根为2级根, 依次为3级根[11]。用刀片切取各分级根3—5 cm的根段后立即放入到FAA固定液, 每株取5段, 冰箱进行保存24 h后, 使用微波快速石蜡切片法[31]对根段进行脱水、透明、浸蜡、包埋和切片, 后对切片进行脱蜡、复水等[32]。使用全电动荧光立体显微镜(HT1600CN)观察木麻黄根系结构, 每个切片观察10个视野;应用Axio-Vision分析软件测量根直径、根横截面积、表皮厚度、皮层细胞面积、维管柱面积、导管数量和导管直径等解剖结构指标, 并根据上述数据计算中柱占比。
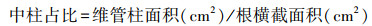
|
(7) |
将幼苗前三级根序细根依次混合后粉碎过筛[33], 用碳氮元素分析仪(VARIO MAX, Elementar, Germany)灼烧测得样品碳、氮含量(TC、TN)[34], 每个处理重复3次。
1.4 数据分析采用单因素方差分析(One-way ANOVA)进行显著性检验(P < 0.05), 使用Origin 2022软件作图。
2 结果分析 2.1 光强对木麻黄生长及生物量累积及分配的影响光限制下木麻黄生物量的累积及分配发生改变。从表 1看出, CK下地径增量、叶干重和地上部分生物量均为最大。伴随光强减弱, L1下地径增量、叶干重和地上部分生物量较CK下降, 但仅地径增量显著下降(P<0.05), 较CK下降84%, 株高、茎干重、根干重及根冠比则较CK有所上升, 但均不显著(P>0.05), L2下各指标除根冠比外较CK均有所下降, 其中株高增量、地径增量及叶干重达到显著水平(P<0.05), L3下株高、地径增量、叶干重、根干重和地上部分生物量则降至最低, 分别较CK降低87.5%、88.3%、57.2%、38.8%、45.8%。光限制下, 茎干重、根干重及根冠比呈现相同变化, 但处理间并不存在显著差异(P>0.05)。
2.2 光强对根系外部形态特征的影响如图 1所示, 木麻黄幼苗根系外部形态生长表现出明显规律性, 细根(0<d≤0.5 mm)生长受限明显, 除Ⅰ级根尖数外, Ⅰ级根长、根表面积、根体积逐渐下降并在L3降至最低;Ⅱ级根表面积、根体积及根尖数虽然在L1时较CK有所上升, 但均不显著, 整体上Ⅱ级根长、根表面积、根体积及根尖数与Ⅰ级指标变化一致并在L3达到最小值。适当的遮光促进粗根(d>2 mm)的生长, 除Ⅲ级根尖数外, Ⅲ、Ⅳ级各指标均表现为L1>L2>CK>L3。除Ⅳ级各指标及Ⅱ级根尖数外, Ⅰ、Ⅱ、Ⅲ指标间差异均达到显著水平(P < 0.05)。

|
| 图 1 光强对不同径级木麻黄幼苗根系外部形态构建的影响 Fig. 1 Effect of light intensity on the external morphological construction of the root system of different diameter classes of C.equisetifolia seedings CK:100%自然全光照;L1:55%自然全光照;L2:30%自然全光照;L3:15%自然全光照;d:根系径级 |
从根系形态指标总量来看, 木麻黄幼苗根系各指标随光照强度的降低呈现“收缩加粗”趋势(图 2)。其中木麻黄总根长逐渐下降, L1、L2、L3下根长较CK降低15.4%、22.7%、30.3%;根平均直径和根体积则表现为L2>L3>L1>CK, 处理间存在显著差异(P<0.05)。此外, 根生物量、比根长、比表面积、根组织密度、根尖数随光强下降呈现波动变化, 但均不存在显著差异(P>0.05)。

|
| 图 2 光强对木麻黄幼苗根系外部形态构建的影响 Fig. 2 Effect of light intensity on the external morphological construction of the root system of C.equisetifolia seedings |
在不同光强下不同径级解剖结构的响应不同(图 3)。随光限制加重, 1级细根横截面积、根半径、表皮厚度、维管柱面积、中柱占比、导管平均直径等指标逐渐下降, 并均在L3达到最小值(1579.25 μm2、22.03 μm、0.73 μm、115.39 μm2、0.07、0.66 μm), 皮层细胞面积逐渐上升在L3达到最大值1463.85 μm2, 导管数量表现为CK>L3>L2>L1, 各指标间存在显著差异(P<0.05)。

|
| 图 3 光强对木麻黄各级根系解剖结构的影响 Fig. 3 Effect of light intensity on the anatomical structure of the root system at all levels of C. equisetifolia seedings |
CK下2级细根横截面积、根半径、皮层细胞面积、皮层厚度、导管平均直径均在最高水平, 但随光强下降, 其余处理横截面积、根半径、皮层细胞面积、表皮厚度较CK显著降低, 而CK下维管柱面积和导管平均直径较L1无显著差异, 但L2、L3较CK则明显下降;
L1下中柱占比、导管数量及L2导管数量则较CK显著上升, 其中L1下中柱占比、导管数量达到最大值(0.19、14.09), 且显著高于其他组(P < 0.05)。就L1、L2、L3而言, 横截面积、皮层细胞面积表现为L3>L2>L1, 维管柱面积、中柱占比、导管数量则相反, 具体表现为L1>L2>L3。此外, L1、L2及L3中根半径有所波动变化但均无显著差异(P>0.05), 表皮厚度则逐渐下降并在L3时达到最小值(0.46 μm)。
随光照强度下降, 3级细根的横截面积、根半径、皮层细胞面积表现为L1>L2>L3>CK, 表皮厚度仅L2较CK明显下降, 其余处理较CK无显著差异(P>0.05)。维管柱面积和中柱占比均在L1达到最小值(457.614 μm2、0.0675), 较CK明显下降, 但随光强减弱程度加深, L2、L3维管柱面积和中柱占比较CK呈显著上升。导管数量则仅L3较CK明显下降(P < 0.05)。CK下导管平均直径与各处理间存在显著差异(P>0.05), 并在L2时达到最小值0.79 μm。
2.4 光强对细根C、N含量的影响由图 4看出, 木麻黄幼苗C、N含量及CN比在不同光照处理下存在着明显的差异。随光强下降, CK至L2时TC含量逐渐降低, 而L3下TC含量骤升并达到最高(363.99 mg/g);L1的TN含量较CK有所下降但无显著差异(P>0.05), 但随光照减弱程度加深, TN含量在L2、L3出现明显上升, 并在L3达到最大值(13.93 mg/g)。C﹕N在CK时最大(27.79), 各处理较CK均显著下降(P<0.05), L2时达到最小值(23.84), 较CK下降14.21%。

|
| 图 4 光强对木麻黄根系碳氮含量的影响 Fig. 4 Effect of light intensity on the carbon and nitrogen content of C. equisetifolia seedings |
光照不足通过限制光合作用来胁迫植物, 导致碳积累减少和生长速率减缓, 甚至引起植株的死亡, 但植物也会通过表型可塑性、生物量变化来适应环境, 增加生存率[2, 4]。适度遮荫下, 木麻黄幼苗以株高生长来增加获得更多光能的接触面积;但光限制过强, 幼苗生长及形态调节能力减弱, 捕获的光能量减少导致CO2固定和碳水化合物积累下降造成生长、发育受限[35—36]。光照降低后植物生长能力减弱与有机物质减少是一个相互影响的过程, 叶片的发育不良将进一步导致光合能力的下降[2]。
植物生物量取决于提供给植物的光总量, 而生物量分配的变化反映其生长或存活的策略[1]。光资源减少后, 木麻黄生物量下降显著, 但茎、根生物量下降幅度远小于叶。平衡生长假说认为植物会将生物量优先分配给当前处于生长资源限制的植物器官[37], 而木麻黄幼苗对不同器官的碳分配策略似乎与此并不完全相同。研究认为荫蔽下幼苗以纵向生长优先, 并会牺牲叶的扩展将更多的生物量分配给茎的生长[2]。光限制下根冠比呈波动变化但并不显著, 即不能改变木麻黄幼苗生物量的分配的总格局, 这可能与木麻黄一直以来的生活策略有关。沿海土壤养分贫瘠, 木麻黄必须不断更新细根, 以利于吸收水分和养分, 研究表明木麻黄群落生物量研究中其茎、枝、根的生物量分配占比远大于叶[38—40]。在过去的研究中也发现其它植物具有类似木麻黄幼苗生物量分配方式即将更多的光合产物用于茎、根系储备而非生长[41], 这对于幼苗存活到成年成为优势个体具有重要意义。
3.2 光强对木麻黄根系外部形态的影响光限制抑制木麻黄幼苗根系纵向伸长但促进横向生长。根长、根表面积、根体积、根直径和根尖数等作为衡量植物生长状态的重要指标, 反映根系的延伸范围以及根系的发育状况等, 并直接影响到地上部分的生长和林木的抗逆性[42]。木麻黄幼苗总根长随光强降幅增大下降趋势愈加明显, 原因是光照强度的改变会直接影响光合产物向根系的运转, 改变根系形态结构的建成[43];总根平均直径随光强的降低而增大, 表明弱光胁迫时, 植物根系通过加粗根直径, 提升根体积以增大吸水面积和内部通道, 加大对土壤水分和养分的汲取与运输, 适应地上部分分配物质的减少以应对逆境条件[44]。
光强变化引起根系不同径级的异质生长。适度的遮荫, 粗根生长有所促进, 粗根长度、表面积、体积和根尖数的上升有助于养分及水分的摄取运输;但细根表现相反。一方面, 根系被认为是光合产物的“汇”[45], 根系呼吸是森林土壤中CO2排出的主要手段, 根系中细根的呼吸作用远高于粗根, 细根生长速度快, 在根系动态变化过程中较为活跃, 周转要消耗大量的碳, 为适应光限制下碳源的减少, 幼苗减缓细根生长发育避免过多的呼吸作用消耗更多的能量[46]。另一方面, 光照胁迫下木麻黄在根系构建上选择粗根而非细根, 因为粗根较细根有更长的寿命[47], 动态变化较小。
此外, 实验结果显示各处理间的总根尖数、比根长、比表面积及组织密度间差异未达显著水平可能是前者主要受树种遗传物质控制, 而后三者主要与根系干物质积累以及根长、根表面积及根体积变化相关, 除自身特性外, 还与环境变化直接影响有较大关系[4]。
3.3 光强对木麻黄根系解剖形态的影响不同根序根系结构对光照强度的适应程度不同, 应对策略不同(图 3、图 5)。随着光限制的提升1级细根功能改变, 幼苗1级细根维管柱面积减少而皮层细胞面积的增加, 幼苗1级细根横向运输能力增强而纵向运输能力减弱, 下降的横截面积及表皮厚度使得细根更易进入小体积的土壤中探索并减少水分及养分进入木质结构的障碍, 中柱和导管结构上的改变意味1级细根由运输转向吸收和存储养分转变[48—49]。

|
| 图 5 光强对木麻黄1—3级根序解剖结构的影响 Fig. 5 Pictures of the anatomical structure of the first to third order roots of C.equisetifolia seedings under different light intensities 1级和2级根系均于20×0.4倍物镜下拍摄, 3级根于10×0.25倍物镜下拍摄 |
2级细根对不同程度光照响应不同, 轻度光限制下2级细根在牺牲保护功能和存储功能的前提下保证养分、水分运输效率, 提高在土壤养分中汲取能力和向上运输能力, 减小由光照降低导致的植物体内光合产物减少的伤害, 弥补植物各部分可用性物质分配比例不足;重度光限制变化与1级细根相似, 以维管柱面积和导管直径下降及皮层面积上升有助于为后期生存和长期生长储存植物体内有机物质[41, 50]。
3级细根在光限制下生长得到促进, 光限制下3级细根半径增加, 意味着3级细根获得更多资源, 获得更长的寿命[51]。轻度的光限制下3级细根增强皮层吸收功能但不改变维管柱结构的运输能力, 仅导管直径下降, 与之相反则是导管数量有所提升, 这种变化常见于植物应对逆境胁迫[52]。而整体变化说明伴随光限制的加重幼苗依然稳定3级细根的发育, 提升地下部分养分吸收能力的同时也加强运输能力以适应地面光合产物减少的环境, 这可能与3级细根为过渡根系在功能上具有多样性有关[49]。目前关于根系对光照环境变化的响应规律及策略尚明确, 木麻黄各级细根结构差异可能原因是光照强度调节植物体内生长素生成等间接影响到根系结构变化[53]。Tomáš W[54]等研究表明光和细胞分裂素共同调节植物地上、地下器官发育的许多重要方面, 而在郁闭度由弱到强的光合适应梯度中, 阴影叶输入的根源性细胞分裂素减少, 致使其反应基因表达水平降低, 影响根顶端分生组织活性[55], 抑制根系发育。
此外, 各分级根系表皮、皮层、维管柱及导管组织发育程度除受到植株养分、水分的有效性利用影响外, 还与物种间的差异相关。Hong[21]等人对91种植物细根进行解剖, 其中草本植物随根序增加皮层厚度上升, 而白桦(Betula platyphylla)、水曲柳(Fraxinus mandschurica)、黄檗(Phellodendron amurense)等9种乔木树种细根皮层结构在前4级无显著差异, 这意味着前4级根均为吸收根, 一般认为随着根序增加, 木本植物细根皮层细胞逐渐减少(或消失), 中柱逐渐增加[21, 56], 而在Valenzuela Estrada等人[57]对高丛蓝莓(Vaccinium corymbosum)研究发现, 前3级根均未出现木质结构, 因此不同物种间吸收和运输能力的根系所包括的根序范围可能是不同的[49]。从解剖结果来看, 伴随根序增加木麻黄幼苗直径增加、表皮变薄、皮层组织逐渐完善、维管束退化、导管由宽导管变为窄导管、木质化程度减弱、代谢慢、养分吸收速率加快, 功能由运输向存储及吸收养分转变, 这可能是木麻黄细根在光限制下变化的关键。
3.4 光强对细根C、N浓度的影响研究发现随光照强度下降, 细根逐渐衰老且存在顺序性(低级根序碳水化合物浓度低而随根序增加碳水化合物浓度增加), 而在养分上细根的N含量明显提高[58—59]。光限制下细根TC含量与根系形态变化相关, 当胁迫加重时, 碳源减少让幼苗在构建形态选择粗根, 而较粗的根木质化程度较高, 根形态的构建需要更多的C(图 4)[60]。TN则是在L2显著上升并于L3达到最大(图 4), 充足的N素保证皮层组织构建, 皮层作为初生组织, 内部含有大量以蛋白质形式存在的N, 在胁迫下增强细根的吸收及周转功能[9, 61]。植物形态变化决定其功能作用, 而功能作用归根到底是其生理生化的外在表现[33], C、N含量的变化与木麻黄根系幼苗存活具有更重要的意义相符, 这表明即使在CO2固定减少和碳水化合物稀缺的情况下, 幼苗仍将大量C、N分配给细根以完成自身生长发育和植物养分供应。以往研究显示C∶N比与细根寿命呈正相关[62], 而在碳氮比下降的情况下意味着光合产物与呼吸作用间的不平衡, 这将加快细根的衰老和死亡, 表明光限制下细根受损和生长受限是木麻黄幼苗生长缓慢甚至死亡的主要因素, 并由此影响林下木麻黄自我更新。
4 结论木麻黄是强阳性树种, 轻度遮荫有利幼苗生长, 但弱光环境显著影响到正常生长。轻度光限制促进株高, 但低于适宜光照时, 植物生长势逐渐下降, 株高增长率降低, 不利于植株幼苗的生长和物质的积累。根系形态特征表现为轻度光限制下粗根生长有所提升, 但伴随光限制加重, 粗根、细根生长均受到抑制, 幼苗通过增加根系直径、向周围环境延伸, 提高吸收能力。光强减弱, 1、2级细根由运输功能由吸收为主转为存储为主, 根系皮层细胞面积和减小维管柱面积增强;3级细根各项功能增强以提高植株储存有机质能力以维持正常生长。整体上, 木麻黄幼苗根系会采取“弱化吸收, 强化储存”收缩型生长策略, 通过增强根系直径、增大皮层细胞或减小维管柱面积等增强储存能力的方式来应对恶劣环境。光限制条件也会造成细根C﹕N的不平衡导致细根受损和养分投入的浪费并最终影响林下幼苗成活。因此, 今后的经营管理策略上可以考虑空间上交替进行适当的间择伐, 维持林下适当的光照强度以利于林下苗木的天然自我更新, 维持种群和群落的稳定性。
| [1] |
Bochenek G M, Fällström I. The effect of diurnal light intensity distribution on plant productivity in a controlled environment. Acta Horticulturae, 2016(1134): 155-162. |
| [2] |
Shafiq I, Hussain S, Ali RAZA M, Iqbal N, Asghar M A, Raza A, Fan Y F, Mumtaz M, Shoaib M, Ansar M, Manaf A, Yang W Y, Yang F. Crop photosynthetic response to light quality and light intensity. Journal of Integrative Agriculture, 2021, 20(1): 4-23. DOI:10.1016/S2095-3119(20)63227-0 |
| [3] |
Mishio M, Kawakubo N. Variations in leaf morpho-anatomy and photosynthetic traits between Sun and shade populations of Eurya japonica (Pentaphylacaceae) whose seeds are dispersed by birds across habitats. Plant Species Biology, 2015, 30(2): 147-158. DOI:10.1111/1442-1984.12045 |
| [4] |
Yokawa K, Fasano R, Kagenishi T, Baluška F. Light as stress factor to plant roots-case of root halotropism. Frontiers in Plant Science, 2014, 5: 718. |
| [5] |
潘晓迪, 张颖, 邵萌, 马黎明, 郭新宇. 作物根系结构对干旱胁迫的适应性研究进展. 中国农业科技导报, 2017, 19(2): 51-58. |
| [6] |
Dyachok J, Zhu L, Liao F Q, He J, Huq E, Blancaflor E B. SCAR mediates light-induced root elongation in Arabidopsis through photoreceptors and proteasomes. The Plant Cell, 2011, 23(10): 3610-3626. DOI:10.1105/tpc.111.088823 |
| [7] |
Silva-Navas J, Moreno-Risueno M A, Manzano C, Pallero-Baena M, Navarro-Neila S, Téllez-Robledo B, Garcia-Mina J M, Baigorri R, Gallego F J, del Pozo J C. D-Root: a system for cultivating plants with the roots in darkness or under different light conditions. The Plant Journal: for Cell and Molecular Biology, 2015, 84(1): 244-255. DOI:10.1111/tpj.12998 |
| [8] |
Warren J M, Hanson P J, Iversen C M, Kumar J, Walker A P, Wullschleger S D. Root structural and functional dynamics in terrestrial biosphere models—evaluation and recommendations. The New Phytologist, 2015, 205(1): 59-78. DOI:10.1111/nph.13034 |
| [9] |
McCormack M L, Dickie I A, Eissenstat D M, Fahey T J, Fernandez C W, Guo D L, Helmisaari H S, Hobbie E A, Iversen C M, Jackson R B, Leppälammi-Kujansuu J, Norby R J, Phillips R P, Pregitzer K S, Pritchard S G, Rewald B, Zadworny M. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. The New Phytologist, 2015, 207(3): 505-518. DOI:10.1111/nph.13363 |
| [10] |
Vogt K A, Grier C C, Gower S T, Sprugel D G, Vogt D J. Overestimation of net root production: a real or imaginary problem?. Ecology, 1986, 67(2): 577-579. DOI:10.2307/1938601 |
| [11] |
Pregitzer K S, DeForest J L, Burton A J, Allen M F, Ruess R W, Hendrick R L. Fine root architecture of nine North American trees. Ecological Monographs, 2002, 72(2): 293-309. DOI:10.1890/0012-9615(2002)072[0293:FRAONN]2.0.CO;2 |
| [12] |
De Zoysa M. Casuarina coastal forest shelterbelts in hambantota city, Sri Lanka: assessment of impacts. Small-scale Forestry, 2008, 7(1): 17-27. DOI:10.1007/s11842-008-9038-2 |
| [13] |
Hardman C J, Williams S, Manco B N, Hamilton M A. Predicting the potential threat of Casuarina equisetifolia to three endemic plant species on the Turks and Caicos Islands. Oryx, 2012, 46(2): 204-212. DOI:10.1017/S0030605311000251 |
| [14] |
杨青青, 杨众养, 余雪标, 薛杨, 王小燕, 陈小花, 崔喜博, 李然, 高刘. 不同林龄木麻黄沿海防护林林下植被多样性研究. 热带作物学报, 2016, 37(2): 359-364. |
| [15] |
杨彬, 王玉, 郝清玉. 海南岛木麻黄林林下植物天然更新影响因素的研究. 广西植物, 2020, 40(3): 422-432. |
| [16] |
管康林. 木麻黄种子的光萌发生理研究. 浙江林学院学报, 1984, 1(0): 89-95. |
| [17] |
王玉, 杨彬, 郝清玉. 木麻黄种子萌发的限制生态因子. 广西植物, 2020, 40(3): 403-411. |
| [18] |
张勇, 仲崇禄, 陈羽, 陈珍. 木麻黄共生菌的研究进展. 广东林业科技, 2006, 22(1): 70-75. |
| [19] |
黄云鹏, 聂森, 黄雍容, 叶功富, 廖蒙蒙. 不同林龄木麻黄光合生理日变化及影响因子. 福建林业科技, 2022, 49(4): 1-7. |
| [20] |
王明怀, 许秀玉, 魏龙, 陈建新, 温小莹, 胡新颖, 张方友. 木麻黄无性系光响应特性研究. 华南农业大学学报, 2011, 32(2): 76-79. |
| [21] |
Wang H F, Wang Z Q, Dong X Y. Anatomical structures of fine roots of 91 vascular plant species from four groups in a temperate forest in Northeast China. PLoS One, 2019, 14(5): e021512. |
| [22] |
Weigelt A, Mommer L, Andraczek K, Iversen C M, Bergmann J, Bruelheide H, Fan Y, Freschet G T, Guerrero-Ramírez N R, Kattge J, Kuyper T W, Laughlin D C, Meier I C, van der Plas F, Poorter H, Roumet C, van Ruijven J, Sabatini F M, Semchenko M, Sweeney C J, Valverde-Barrantes O J, York L M, McCormack M L. An integrated framework of plant form and function: the belowground perspective. The New Phytologist, 2021, 232(1): 42-59. DOI:10.1111/nph.17590 |
| [23] |
孙战, 王圣洁, 王旭, 邹文涛, 马海宾. 木麻黄与根系微生物关系研究进展. 世界林业研究, 2020, 33(4): 25-30. |
| [24] |
梁安洁, 龙凤, 洪滔, 林勇明, 陈灿, 谢安强, 吴承祯, 洪伟, 李键. 槲皮黄素-3-α-阿拉伯糖苷胁迫下内生真菌对木麻黄幼苗根系形态和生理特性的影响. 应用与环境生物学报, 2018, 24(4): 797-804. |
| [25] |
林微微. 两种化感物质胁迫下内生真菌对短枝木麻黄幼苗根系形态和生理特性的影响研究[D]. 福州: 福建农林大学, 2020.
|
| [26] |
徐飞, 郭卫华, 徐伟红, 王仁卿. 不同光环境对麻栎和刺槐幼苗生长和光合特征的影响. 生态学报, 2010, 30(12): 3098-3107. |
| [27] |
程晶, 刘济明, 王灯, 王姝, 李丽霞, 陈敬忠. 喀斯特特有植物罗甸小米核桃幼苗对光照强度的可塑性响应. 应用与环境生物学报, 2021, 27(1): 23-30. |
| [28] |
吴敏, 张文辉, 周建云, 马闯, 韩文娟. 干旱胁迫对栓皮栎幼苗细根的生长与生理生化指标的影响. 生态学报, 2014, 34(15): 4223-4233. DOI:10.5846/stxb201212101774 |
| [29] |
王力朋, 晏紫伊, 李吉跃, 王军辉, 何茜, 苏艳, 陈博, 马建伟, 董菊兰. 指数施肥对楸树无性系生物量分配和根系形态的影响. 生态学报, 2012, 32(23): 7452-7462. DOI:10.5846/stxb201203040288 |
| [30] |
王贞升, 李彦雪, 于成龙, 狄小琳, 陈鹏, 田静瑶, 王竞红. 不同模拟降水量下草地早熟禾根系形态与解剖结构的动态变化. 草业学报, 2020, 29(10): 70-80. |
| [31] |
魏欣悦, 苏源, 杨静, 刘林, 李成云. 微波快速石蜡切片法观察水稻叶片组织. 云南农业大学学报: 自然科学版, 2011, 26(4): 454-457, 478. |
| [32] |
杨虎彪, 李晓霞, 罗丽娟. 植物石蜡制片中透明和脱蜡技术的改良. 植物学报, 2009, 44(2): 230-235. |
| [33] |
洪梓明, 邢亚娟, 闫国永, 张军辉, 王庆贵. 长白山白桦山杨次生林细根形态特征和解剖结构对氮沉降的响应. 生态学报, 2020, 40(2): 608-620. |
| [34] |
舒李祥, 席国荣, 李月娥, 王晓雪, 马闯, 王博琳. 栓皮栎根系解剖结构、水力特性及碳、氮含量研究. 广西植物, 2020, 40(8): 1203-1210. |
| [35] |
刘泽彬, 程瑞梅, 肖文发, 郭泉水, 王娜. 遮荫对中华蚊母树苗期生长及光合特性的影响. 林业科学, 2015, 51(2): 129-136. |
| [36] |
王振兴, 朱锦懋, 王健, 汪滢, 卢钰茜, 郑群瑞. 闽楠幼树光合特性及生物量分配对光环境的响应. 生态学报, 2012, 32(12): 3841-3848. DOI:10.5846/stxb201112021842 |
| [37] |
Müller I, Schmid B, Weiner J. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology, Evolution and Systematics, 2000, 3(2): 115-127. DOI:10.1078/1433-8319-00007 |
| [38] |
Xue Y, Yang Z Y, Wang X Y, Lin Z P, Li D X, Su S F. Tree biomass allocation and its model additivity for Casuarina equisetifolia in a tropical forest of Hainan Island, China. PLoS One, 2016, 11(3): e0151858. DOI:10.1371/journal.pone.0151858 |
| [39] |
叶功富, 张清海, 林益明, 吴锡麟. 沿海防护林生态系统不同群落生物量和能量的研究. 林业科学, 2003, 39(S1): 8-14. |
| [40] |
叶功富, 张立华, 侯杰, 卢昌义, 吴柳清, 李秀明. 滨海沙地木麻黄人工林细根生物量及其动态研究. 应用与环境生物学报, 2007, 13(4): 481-485. |
| [41] |
Canham C D, Kobe R K, Latty E F, Chazdon R L. Interspecific and intraspecific variation in tree seedling survival: effects of allocation to roots versus carbohydrate reserves. Oecologia, 1999, 121(1): 1-11. DOI:10.1007/s004420050900 |
| [42] |
嵇晓雷, 杨平. 关于植物根系形态分布研究进展与新方法探讨. 森林工程, 2011, 27(4): 54-57. |
| [43] |
吕军, 王伯伦, 孟维韧, 赵凤艳. 不同穗型粳稻的光合作用与物质生产特性. 中国农业科学, 2007, 40(5): 902-908. |
| [44] |
Zadworny M, McCormack M L, Mucha J, Reich P B, Oleksyn J. Scots pine fine roots adjust along a 2000-km latitudinal climatic gradient. The New Phytologist, 2016, 212(2): 389-399. DOI:10.1111/nph.14048 |
| [45] |
杨振亚, 周本智, 陈庆标, 葛晓改, 王小明, 曹永慧, 童冉, 石洋. 干旱对杉木幼苗根系构型及非结构性碳水化合物的影响. 生态学报, 2018, 38(18): 6729-6740. DOI:10.5846/stxb201803260604 |
| [46] |
Bartsch N. Responses of root systems of young Pinussylvestris and Piceaabies plants to water deficits and soil acidity. Canadian Journal of Forest Research, 1987, 17(8): 805-812. DOI:10.1139/x87-128 |
| [47] |
于水强, 王静波, 郝倩葳, 王维枫, 王琪, 詹龙飞. 四种不同生活型树种细根寿命及影响因素. 生态学报, 2020, 40(9): 3040-3047. DOI:10.5846/stxb201904020641 |
| [48] |
刘洪凯, 陈旭, 张明忠, 王强, 王延平. 鲁中丘陵山地干旱生境上11个树种的细根解剖特征与耐旱策略. 林业科学, 2020, 56(7): 185-193. |
| [49] |
谷加存, 王东男, 夏秀雪, 王韶仲. 功能划分方法在树木细根生物量研究中的应用: 进展与评述. 植物生态学报, 2016, 40(12): 1344-1351. |
| [50] |
汪攀, 陈奶莲, 邹显花, 马祥庆, 吴鹏飞. 植物根系解剖结构对逆境胁迫响应的研究进展. 生态学杂志, 2015, 34(2): 550-556. |
| [51] |
Ma Z Q, Guo D L, Xu X L, Lu M Z, Bardgett R D, Eissenstat D M, McCormack M L, Hedin L O. Evolutionary history resolves global organization of root functional traits. Nature, 2018, 555(7694): 94-97. DOI:10.1038/nature25783 |
| [52] |
陈旭, 陈冬倩, 刘洪凯, 赵春周, 赵文太, 董智, 张永涛, 王延平. 黄河三角洲滨海盐碱地11个造林树种细根解剖性状对土壤条件的响应. 生态学报, 2021, 41(10): 4150-4159. DOI:10.5846/stxb201911122384 |
| [53] |
Yun F H, Liu H W, Deng Y Z, Hou X M, Liao W B. The role of light-regulated auxin signaling in root development. International Journal of Molecular Sciences, 2023, 24(6): 5253. DOI:10.3390/ijms24065253 |
| [54] |
Werner T, Schmülling T. Cytokinin action in plant development. Current Opinion in Plant Biology, 2009, 12(5): 527-538. DOI:10.1016/j.pbi.2009.07.002 |
| [55] |
Boonman A, Prinsen E, Gilmer F, Schurr U, Peeters A J M, Voesenek L A C J, Pons T L. Cytokinin import rate as a signal for photosynthetic acclimation to canopy light gradients. Plant Physiology, 2007, 143(4): 1841-1852. DOI:10.1104/pp.106.094631 |
| [56] |
Guo D L, Xia M X, Wei X, Chang W J, Liu Y, Wang Z Q. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. The New Phytologist, 2008, 180(3): 673-683. DOI:10.1111/j.1469-8137.2008.02573.x |
| [57] |
Valenzuela-Estrada L R, Vera-Caraballo V, Ruth L E, Eissenstat D M. Root anatomy, morphology, and longevity among root orders in Vaccinium corymbosum (Ericaceae). American Journal of Botany, 2008, 95(12): 1506-1514. DOI:10.3732/ajb.0800092 |
| [58] |
徐文静, 王政权, 范志强, 孙海龙, 贾淑霞, 吴楚. 遮荫对水曲柳幼苗细根衰老的影响. 植物生态学报, 2006, 30(1): 104-111. |
| [59] |
霍常富, 王政权, 孙海龙, 范志强, 赵晓敏. 光照和氮交互作用对水曲柳幼苗生长、生物量和氮分配的影响. 应用生态学报, 2008, 19(8): 1658-1664. |
| [60] |
常文静, 郭大立. 中国温带、亚热带和热带森林45个常见树种细根直径变异. 植物生态学报, 2008, 32(6): 1248-1257. |
| [61] |
闫国永, 王晓春, 邢亚娟, 韩士杰, 王庆贵. 兴安落叶松林细根解剖结构和化学组分对N沉降的响应. 北京林业大学学报, 2016, 38(4): 36-43. |
| [62] |
许旸, 谷加存, 董雪云, 刘颖, 王政权. 海南岛4个热带阔叶树种前5级细根的形态、解剖结构和组织碳氮含量. 植物生态学报, 2011, 35(9): 955-964. |