 2024, Vol. 44
2024, Vol. 44文章信息
- 雷德芳, 王志杰
- LEI Defang, WANG Zhijie
- 黔中喀斯特地区马尾松群落主要物种种间联结、生态位及群落稳定性特征
- Interspecific associations, niches and community stability of main species in Pinus massoniana community in karst mountainous cities, the central Guizhou Province
- 生态学报. 2024, 44(10): 4353-4367
- Acta Ecologica Sinica. 2024, 44(10): 4353-4367
- http://dx.doi.org/10.20103/j.stxb.202305261119
-
文章历史
- 收稿日期: 2023-06-26
- 网络出版日期: 2024-02-28






群落是具有特定功能的生物集合体, 群落内的物种通过相互作用形成复杂的稳定共存关系[1], 这种共存关系常常由物种的种间联结及生态位特征反映。种间联结指群落内不同物种在空间上的相互关联性[2], 是植物群落数量和结构的一个重要特征, 也是揭示种群竞争状况及群落演替的有效方法[3]。生态位是指群落中各种群所占据的时空位置及其与其他相关种群间的功能关系, 对预测群落演替和理解物种共存机制有重要作用[4]。生态位测度包括生态位宽度和生态位重叠, 生态位宽度不仅可用于衡量物种在特定栖息地综合利用环境的能力, 还代表它们在生态系统中的地位和分布[5—6]。群落稳定性是森林生态系统持续运作的基础, 是植物群落结构与功能的综合特征[7]。一般而言, 群落内正联结种对越多, 生态位越宽, 功能多样性越高, 群落越稳定;反之, 群落内负关联种对越多, 生态位越窄, 功能多样性越低, 群落越不稳定[8—9]。近年来, 国内外学者针对不同群落类型或不同生活型植物的物种生态位和种间联结的关系, 以及群落内种间联结性与群落稳定性的关系等方面开展了大量的研究[10—14], 揭示了群落植物种间联结与生态位重叠关系密切、与群落稳定性具有同步性等特征[15—16], 但关于群落内植物种间联结、生态位宽度与重叠以及群落稳定性的综合研究较为薄弱, 而三者的综合研究可以全面有效的反映群落的基本结构、功能以及演替规律, 为森林管理和自然植被恢复提供理论依据[10]。
马尾松(Pinus massoniana)作为中国南方主要造林用材树种, 具有适应能力强、生长速度快、耐干旱瘠薄的特点, 是重要的荒山造林和生态建设树种之一, 有较高的生态、经济和社会价值[17—18]。马尾松森林生态系统在生物多样性保护、生态系统功能维持以及人类福祉方面发挥着关键作用[19]。以贵州为中心的我国西南地区是全球面积最大、最集中连片的喀斯特区域, 其独特的地质背景和不合理的土地利用, 导致了土壤退化、生态环境恶化等严重的生态环境问题, 严重制约了当地经济和社会的可持续发展[20—21]。马尾松作为先锋树种是喀斯特地区植被恢复和生态建设的关键前提[22]。因此, 针对喀斯特地区典型的马尾松群落的种内/种间关系、生态位和群落稳定性进行研究, 对于科学开展森林经营管理, 提高森林质量, 挖掘绿水青山的生态价值, 推进森林可持续发展具有重要意义。
近年来, 针对马尾松森林生态系统的研究已取得丰硕成果[23—26], 主要集中在群落结构[27—28]、不同生长阶段的生物量与碳储量[29]以及种间联结性[30—32]等方面。然而, 马尾松群落不同层次(乔木层、灌木层、草本层)的种间联结性、生态位特征和稳定性的综合研究鲜有报道。鉴于此, 本研究以黔中喀斯特地区典型的马尾松群落为研究对象, 在植物群落学调查的基础上, 采用方差比率法(VR)、χ2检验、Spearman秩相关检验、生态位测定和M.Godron稳定性等方法, 对马尾松群落主要物种的种间关联、生态位特征及群落稳定性进行定量分析, 以期揭示黔中喀斯特地区马尾松群落不同垂直层次主要物种的种间关系、群落组成的结构特点以及群落的演替趋势, 为喀斯特地区植被恢复与重建、生物多样性保护、森林可持续经营提供理论指导和决策依据。
1 材料与方法 1.1 研究区概况贵阳市乌当区(106°30′—107°03′ E、26°55′—26°33′ N), 位于贵阳市东北部, 地处黔中腹地, 属云贵高原苗岭山脉中段, 贵州高原的第二台阶上, 长江流域和珠江流域的分水岭带, 地势北高南低, 平均海拔1242 m[33]。全区地貌以山地、丘陵、坝地为主, 气候类型属于亚热带季风湿润气候, 冬无严寒, 夏无酷暑, 年平均降水量1179.8—1271 mm, 年平均气温13.5℃[34]。全区基岩主要为石灰岩, 土壤类型以黄棕壤、石灰土为主, 厚度分布不均匀[35]。研究区的马尾松主要为次生林, 分布在海拔较低的坡地和峡谷, 高度4—27 m[36]。群落层次结构相对简单, 成层现象明显, 乔木层以马尾松为建群种, 伴生种主要有栗(Castanea mollissima)、亮叶桦(Betula luminifera)、槲栎(Quercus aliena);灌木层物种组成丰富, 主要物种铁仔(Myrsine africana)、烟管荚蒾(Viburnum utile)、山莓(Rubus corchorifolius)以及槲栎和栗等乔木幼树;草本层物种种类较丰富, 盖度较低, 主要物种芒萁(Dicranopteris pedata)、贵州鳞毛蕨(Dryopteris wallichiana var. kweichowicola)、芒(Miscanthus sinensis)、狗脊(Woodwardia japonica)等。
1.2 群落学调查在实地踏查的基础上, 于2021年7月, 在贵阳市乌当区选取受人为干扰较轻且具有代表性的马尾松成熟林, 采用样带—样地调查法, 沿水平方向和垂直方向分别设置调查样带, 根据山体地形确定样地位置, 样地间隔50—100 m, 共设置了9个投影面积为400 m2的样地, 每个样地设置3个调查样方, 共27个调查样方, 其中:乔木样方10 m×10 m, 灌木样方5 m×5 m, 草本样方2 m×2 m。
对乔木层调查样方中所有胸径≥5 cm的木本植株计入乔木层, 灌木层调查对象包括灌木物种以及未满足乔木层测量标准的更新幼树(胸径<5 cm), 记录物种名称、胸径、丰富度、基径和冠幅等指标, 同时记录每个样方的经纬度、坡向、坡位、海拔、坡度、岩石裸露率和干扰程度等基本信息。
1.3 研究方法 1.3.1 物种重要值27个样方共记录植物112种, 其中, 乔木层植物19种, 灌木层植物52种, 草本层植物41种。优势物种对群落结构和群落环境的形成具有很大影响[37]。因此, 选用重要值代表各物种的优势度, 分乔木层、灌木层和草本层计算重要值, 计算公式如下[38]:

|
本研究分层选取乔木层、灌木层和草本层中重要值较高的主要植物进行种间关联分析。
1.3.2 总体关联性分析采用Schluter[39]提出的基于物种存在或者不存在的方差比率法(VR)分析该群落各层次主要物种之间的总体关联性。其计算公式如下:

|
式中, S为物种总数;N为样方总数;Tj为出现在样方j中物种数;ni为含有物种i的样方数;t为所有样方中物种的平均数。VR为各层次主要物种间的总体联结指数, 若VR>1, 种间总体呈正关联;反之, 呈负关联。利用统计量W检验VR值是否显著偏离1, 若W>x0.05(N)2或W<x0.95(N)2, 则表示物种间关联显著;若x0.95(N)2<W<x0.052(N), 则说明物种间不显著关联。
1.3.3 种间关联性分析采用基于2×2列联表的χ2统计量测定种间关联性, 以确定实测值与期望值之间偏差的显著程度。由于取样方法为非连续性的, 因此计算采用Yates的连续校正公式[40], 其计算公式如下:

|
式中, N为样方总数;a为同时存在物种A和B的样方数;b仅存在物种A的样方数;c为仅存在物种B的样方数;d为物种A和B均未出现的样方数。当ad>bc, 种间关联性为正关联;当ad<bc, 种间关联性为负关联。若χ2<3.841, 种对间独立分布, 种间关联不显著(P>0.05);若3.841≤χ2≤6.635, 种间关联显著(0.01≤P≤0.05);若χ2>6.635, 种间关联极显著(P<0.01)。
1.3.4 种间相关性测定采用各层次主要物种的重要值作为数量指标, 对种对间的线性关系进行Spearman秩相关检验[13]。计算公式如下:

|
式中, xik和xjk分别是物种i和物种j在样方k中的秩, rs(i, j)值域为[-1, 1], 大于零, 表示正相关, 小于零, 表示负相关[4]。
1.3.5 生态位宽度及重叠度分析生态位宽度反映物种对资源开发利用的程度, 数值越大, 代表对环境的适应能力越强[2]。采用Levins[41]提出经修正的公式计算生态位宽度BL:
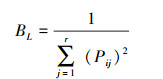
|
式中, Pij为物种i在资源位j上重要值与该物种在所有资源位上重要值总和之比, r为样方总数。
生态位重叠反映了物种之间利用资源或对环境的适应能力的相似程度, 重叠值越大, 物种间的竞争越大[40]。采用Pianka1973[42]提出的公式计算生态位重叠Oij:

|
式中, Oij表示物种i和j的生态位重叠指数, 范围为[-1, 1];r为样方数量, Pik和Pjk分别为物种i和j在第k个资源水平下的重要值占该物种在所有资源水平上重要值总和的比例。
1.3.6 群落稳定性分析采用郑元润[43]改进后的M.Godron稳定性测定法, 将群落中的植物按照相对频度从高到低排列, 计算物种累积倒数百分比作为自变量, 计算累积相对频度作为因变量。然后, 利用散点平滑曲线对二项式方程进行拟合, 使其与直线y=100-x相交, 交点坐标即为所求的群落稳定性比值。根据M.Godron稳定性判断方法, 距离值越小, 群落稳定性越高。
基于R 4.0.4软件程序包spaa中函数sp.pair、sp.assoc、niche.width、niche.overlap, 分别计算种间联结和生态位。
2 结果与分析 2.1 群落总体关联性马尾松群落乔木层、灌木层和草本层的主要物种间的总体关联性基本一致(表 1), 总体关联性的方差比率VR均大于1, 且检验统计量W均落入x0.95(N)2<W<x0.05(N)2中, 表明该群落乔木层、灌木层和草本层的主要物种间的总体关联性均呈不显著正关联。
| 层次 Levels |
方差比率VR Variance ratio |
检验统计量W Test statistic |
x2临界值 x2threshold |
检验结果 Results |
| 乔木层Tree layer | 1.27 | 34.38 | (16.51, 40.11) | 不显著正关联 |
| 灌木层Shrub layer | 1.04 | 28.13 | (16.51, 40.11) | 不显著正关联 |
| 草本层Herb layer | 1.35 | 36.58 | (16.51, 40.11) | 不显著正关联 |
马尾松群落不同层次主要物种的种间关联性性χ2检验结果表明(图 1、2、3):马尾松群落乔木层、灌木层和草本层的主要物种的种间关联主要以不显著或无关联为主, 各层次物种种间关联较松散, 物种间总体呈独立分布格局。具体而言, 乔木层13个主要物种的78个种对中, 66.67%的种对无显著关联, 其中极显著(P<0.01)或显著(P<0.05)正关联和负关联的种对分别为15对和11对(图 1), 如构与朴树、南烛与杉木呈极显著正联结, 朴树与槲栎、槲栎与柃木呈极显著负联结。灌木层15个主要物种的105个种对中, 80.95%的种对无显著关联, 其中极显著(P<0.01)或显著(P<0.05)正关联和负关联的种对分别为5对和15对(图 2), 如山莓和樟呈极显著正联结, 铁仔和樟、山莓和油茶呈极显著负联结。草本层15个主要物种的105个种对中, 67.62%的种对无显著关联, 其中极显著(P<0.01)或显著(P<0.05)正关联和负关联的种对分别为23对和11对(图 3), 如臭鸡屎藤和三叶地锦、毛蕨和三脉紫菀呈极显著正联结, 芒和三叶地锦、淡竹叶和狗脊呈极显著负联结。

|
| 图 1 马尾松群落乔木层主要物种的χ2检验半矩阵图 Fig. 1 Semi-matrix diagram of interspecific association χ2 test among main species in tree layer of Pinus massoniana community Ae:楤木Aralia elata;Bp:构Broussonetia papyrifera;Qa:槲栎Quercus aliena;Cm:栗Castanea mollissima;Bl:亮叶桦Betula luminifera;Ej:柃木Eurya japonica;Pm:马尾松Pinus massoniana;Vb:南烛Vaccinium bracteatum;Cs:朴树Celtis sinensis;Cl:杉木Cunninghamia lanceolata;Gp:算盘子Glochidion puberum;Mr:杨梅Myrica rubra;Cao:油茶Camellia oleifera |

|
| 图 2 马尾松群落灌木层主要物种的χ2检验半矩阵图 Fig. 2 Semi-matrix diagram of interspecific association χ2 test among main species in shrub layer of Pinus massoniana community Ae:楤木Aralia elata;Bp:构Broussonetia papyrifera;Qa:槲栎Quercus aliena;Cm:栗Castanea mollissima;Ej:柃木Eurya japonica;Co:南蛇藤Celastrus orbiculatus;Ll:女贞Ligustrum lucidum;Cs:朴树Celtis sinensis;Rc:山莓Rubus corchorifolius;Ag:蛇葡萄Ampelopsis glandulosa;Ma:铁仔Myrsine africana;Vu:烟管荚蒾Viburnum utile;Dk:野柿Diospyros kaki;Cao:油茶Camellia oleifera;Cc:樟Camphora officinarum |

|
| 图 3 马尾松群落草本层主要物种的χ2检验半矩阵图 Fig. 3 Semi-matrix diagram of interspecific association χ2 test among main species in herb layer of Pinus massoniana community Pc:臭鸡矢藤Paederia cruddasiana;Lg:淡竹叶Lophatherum gracile;Ft:地果Ficus tikoua;Pt:地锦Parthenocissus tricuspidata;Wj:狗脊Woodwardia japonica;Dw:贵州鳞毛蕨Dryopteris wallichiana var kweichowicola;Rb:寒莓Rubus buergeri;Ms:芒Miscanthus sinensis;Dp:芒萁Dicranopteris peddata;Ci:毛蕨Cyclosorus interruptus;Ab:牛膝Achyranthes bidentata;Pq:五叶地锦Parthenocissus quinquefolia;Aa:三脉紫菀Aster ageratoides;Ps:三叶地锦Parthenocissus semicordata;Ca:天名精Carpesium abrotanoides |
马尾松群落不同层次主要物种的Spearman秩相关检验结果表明(图 4、5、6):马尾松群落乔木层、灌木层和草本层的主要物种的种间相关性均呈不显著相关, 各层次物种种间相关性较弱, 绝大部分种对相互独立, 这同χ2检验的结果基本一致。具体而言, 乔木层13个主要物种的78个种对中, 62.83%的种对呈不显著相关, 其中极显著(P<0.001)或显著(P<0.05)正相关和负相关的种对分别为21对和88对(图 4), 如构和朴树, 栗和油茶呈显著正相关(P<0.05)。灌木层15个主要物种的105个种对中, 61.90%的种对呈不显著相关, 其中极显著(P<0.001)或显著(P<0.05)正相关和负相关的种对分别为17对和23对(图 5), 如山莓和构呈显著正相关(P<0.05)。草本层15个主要物种的105个种对中, 63.81%的种对呈不显著相关, 其中极显著(P<0.001)或显著(P<0.05)正相关和负相关的种对分别为15对和23对(图 6), 如狗脊和三脉紫菀、三脉紫菀和牛膝呈显著正相关(P<0.05)。

|
| 图 4 马尾松群落乔木层主要物种的Spearman秩相关检验半矩阵图 Fig. 4 Semi-matrix diagram of Spearman′s rank correlation test among main species in tree layer of Pinus massoniana community * P<0.05;** P<0.01;*** P<0.001 |

|
| 图 5 马尾松群落灌木层主要物种的Spearman秩相关检验半矩阵图 Fig. 5 Semi-matrix diagram of Spearman′s rank correlation test among main species in shrub layer of Pinus massoniana community * P<0.05;** P<0.01;*** P<0.001 |

|
| 图 6 马尾松群落草本层主要物种的Spearman秩相关检验半矩阵图 Fig. 6 Semi-matrix diagram of Spearman′s rank correlation test among main species in herb layer of Pinus massoniana community * P<0.05;** P<0.01;*** P<0.001 |
从马尾松群落各层次主要物种的生态位宽度可以看出(表 2):乔木层主要物种的生态位宽度差异较大, 其中马尾松的生态位宽度最大, 其次是油茶、栗、槲栎、亮叶桦, 均大于4.94;灌木层主要物种生态位宽度较大的物种为栗、槲栎、楤木、铁仔、油茶, 均大于10.61;草本层主要物种生态位宽度较大的物种包括芒萁、贵州鳞毛蕨、臭鸡矢藤、芒、狗脊等, 均大于8.56。
| 生长型 Growth form |
序号 No. |
物种 Species |
相对多度 Relative abundance |
重要值 Importance value |
生态位宽度 Niche width |
| 乔木层 Tree layer |
1 | 马尾松Pinus massoniana | 0.55 | 1.79 | 20.78 |
| 2 | 栗Castanea mollissima | 0.09 | 0.24 | 8.65 | |
| 3 | 亮叶桦Betula luminifera | 0.10 | 0.22 | 4.94 | |
| 4 | 槲栎Quercus aliena | 0.05 | 0.17 | 8.05 | |
| 5 | 杉木Cunninghamia lanceolata | 0.08 | 0.12 | 2.16 | |
| 6 | 南烛Vaccinium bracteatum | 0.02 | 0.06 | 4.00 | |
| 7 | 楤木Aralia elata | 0.02 | 0.06 | 2.78 | |
| 8 | 构Broussonetia papyrifera | 0.02 | 0.05 | 2.67 | |
| 9 | 朴树Celtis sinensis | 0.01 | 0.05 | 3.00 | |
| 10 | 油茶Camellia oleifera | 0.02 | 0.04 | 11.60 | |
| 11 | 柃木Eurya japonica | 0.01 | 0.04 | 1.85 | |
| 12 | 算盘子Glochidion puberum | 0.01 | 0.03 | 2.00 | |
| 13 | 杨梅Myrica rubra | 0.01 | 0.03 | 2.00 | |
| 灌木层 Shrub layer |
1 | 槲栎Quercus aliena | 0.10 | 0.19 | 16.63 |
| 2 | 蛇葡萄Ampelopsis glandulosa | 0.02 | 0.04 | 6.40 | |
| 3 | 楤木Aralia elata | 0.08 | 0.15 | 13.41 | |
| 4 | 构Broussonetia papyrifera | 0.04 | 0.07 | 5.88 | |
| 5 | 樟Camphora officinarum | 0.03 | 0.06 | 4.90 | |
| 6 | 栗Castanea mollissima | 0.09 | 0.19 | 18.47 | |
| 7 | 油茶Camellia oleifera | 0.06 | 0.13 | 10.61 | |
| 8 | 朴树Celtis sinensis | 0.03 | 0.07 | 6.72 | |
| 9 | 南蛇藤Celastrus orbiculatus | 0.03 | 0.05 | 3.70 | |
| 10 | 女贞Ligustrum lucidum | 0.02 | 0.04 | 3.60 | |
| 11 | 野柿Diospyros kaki | 0.02 | 0.04 | 5.40 | |
| 12 | 柃木Eurya japonica | 0.03 | 0.07 | 5.57 | |
| 13 | 铁仔Myrsine africana | 0.07 | 0.13 | 11.95 | |
| 14 | 山莓Rubus corchorifolius | 0.06 | 0.10 | 9.44 | |
| 15 | 烟管荚蒾Viburnum utile | 0.07 | 0.12 | 9.13 | |
| 草本层 Herb layer |
1 | 牛膝Achyranthes bidentata | 0.06 | 0.12 | 5.71 |
| 2 | 三脉紫菀Aster ageratoides | 0.03 | 0.05 | 4.26 | |
| 3 | 天名精Carpesium abrotanoides | 0.02 | 0.04 | 3.27 | |
| 4 | 毛蕨Cyclosorus interruptus | 0.06 | 0.12 | 8.07 | |
| 5 | 芒萁Dicranopteris pedata | 0.18 | 0.29 | 15.17 | |
| 6 | 贵州鳞毛蕨 Dryopteris wallichiana var. kweichowicola |
0.11 | 0.20 | 15.09 | |
| 7 | 地果Ficus tikoua | 0.03 | 0.05 | 3.57 | |
| 8 | 淡竹叶Lophatherum gracile | 0.05 | 0.09 | 8.00 | |
| 9 | 芒Miscanthus sinensis | 0.07 | 0.15 | 10.90 | |
| 10 | 臭鸡矢藤Paederia cruddasiana | 0.05 | 0.12 | 12.80 | |
| 11 | 三叶地锦Parthenocissus semicordata | 0.02 | 0.05 | 5.44 | |
| 12 | 地锦Parthenocissus tricuspidata | 0.03 | 0.07 | 7.14 | |
| 13 | 寒莓Rubus buergeri | 0.04 | 0.07 | 5.44 | |
| 14 | 狗脊Woodwardia japonica | 0.07 | 0.12 | 8.56 | |
| 15 | 五叶地锦Parthenocissus quinquefolia | 0.02 | 0.05 | 1.00 |
进一步分析群落各层次物种间Pianka生态位重叠指数, 可以发现各物种间普遍存在生态位重叠, 生态位宽度越大的物种, 与其他物种产生生态位重叠的概率越大。具体而言, 乔木层中, 62.83%的种对(49对)的Pianka生态位重叠指数大于0, 37.18%的种对(29对)未产生生态位重叠, 其中, 0.5>Oij>0的种对有46对, 占总物种对数的58.97%, Oij>0.5的种对数仅有3对, 占总对数的3.85%。乔木层生态位重叠度指数较大的物种为油茶和栗(0.71)、构和朴树(0.71)、杨梅和油茶(0.67)、杨梅和栗(0.59)(表 3), 表明这些物种对资源利用的相似性很高, 可能存在激烈的竞争。灌木层中, 94.29%的种对(99对)的Pianka生态位重叠指数大于0, 5.71%的种对(6对)未产生生态位重叠, 其中, 0.5>Oij>0的种对有84对, 占总物种对数的80.00%, Oij>0.5的种对数有15对, 占总对数的14.29%。灌木层中生态位重叠度指数较大的有栗和铁仔(0.77)、构和山莓(0.77)、栗和槲栎(0.74)、楤木和山莓(0.69)(表 4)。草本层中, 87.62%的种对(92对)的Pianka生态位重叠指数大于0, 12.38%的种对(13对)未产生生态位重叠, 其中, 0.5>Oij>0的种对有79对, 占总物种对数的75.24%, Oij>0.5的种对数有13对, 占总对数的12.38%。草本层生态位重叠度较大的有牛膝和三脉紫菀(0.71)、狗脊和三脉紫菀(0.70)、狗脊和牛膝(0.69)、地锦和狗脊(0.66)(表 5)。
| 物种 Species |
楤木 Aralia elata |
构 Broussonetia papyrifera |
槲栎 Quercus aliena |
栗 Castanea mollissima |
亮叶桦 Betula luminifera |
柃木 Eurya japonica |
马尾松 Pinus massoniana |
南烛 Vaccinium bracteatum |
朴树 Celtis sinensis |
杉木 Cunninghamia lanceolata |
算盘子 Glochidion puberum |
杨梅 Myrica rubra |
| 构Broussonetia papyrifera | 0.14 | |||||||||||
| 槲栎Quercus aliena | 0.29 | 0.00 | ||||||||||
| 栗Castanea mollissima | 0.00 | 0.06 | 0.40 | |||||||||
| 亮叶桦Betula luminifera | 0.13 | 0.16 | 0.11 | 0.07 | ||||||||
| 柃木Eurya japonica | 0.09 | 0.11 | 0.00 | 0.02 | 0.00 | |||||||
| 马尾松Pinus massoniana | 0.21 | 0.23 | 0.45 | 0.52 | 0.36 | 0.09 | ||||||
| 南烛Vaccinium bracteatum | 0.00 | 0.00 | 0.22 | 0.21 | 0.19 | 0.07 | 0.28 | |||||
| 朴树Celtis sinensis | 0.19 | 0.71 | 0.00 | 0.00 | 0.22 | 0.16 | 0.25 | 0.00 | ||||
| 杉木Cunninghamia lanceolata | 0.00 | 0.00 | 0.20 | 0.09 | 0.02 | 0.00 | 0.18 | 0.45 | 0.00 | |||
| 算盘子Glochidion puberum | 0.24 | 0.29 | 0.00 | 0.10 | 0.14 | 0.19 | 0.34 | 0.35 | 0.41 | 0.00 | ||
| 杨梅Myrica rubra | 0.00 | 0.29 | 0.15 | 0.59 | 0.00 | 0.00 | 0.39 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 油茶Camellia oleifera | 0.00 | 0.00 | 0.21 | 0.71 | 0.00 | 0.00 | 0.36 | 0.00 | 0.00 | 0.18 | 0.00 | 0.67 |
| 物种 Species |
楤木 Aralia elata |
构 Broussonetia papyrifera |
槲栎 Quercus aliena |
栗 Castanea mollissima |
柃木 Eurya japonica |
南蛇藤 Celastrus orbiculatus |
女贞 Ligustrum lucidum |
朴树 Celtis sinensis |
山莓 Rubus corchorifolius |
蛇葡萄 Ampelopsis glandulosa |
铁仔 Myrsine africana |
烟管荚蒾 Viburnum utile |
野柿 Diospyros kaki |
油茶 Camellia oleifera |
| 构Broussonetia papyrifera | 0.62 | |||||||||||||
| 槲栎Quercus aliena | 0.35 | 0.17 | ||||||||||||
| 栗Castanea mollissima | 0.60 | 0.46 | 0.74 | |||||||||||
| 柃木Eurya japonica | 0.16 | 0.11 | 0.44 | 0.27 | ||||||||||
| 南蛇藤Celastrus orbiculatus | 0.41 | 0.01 | 0.36 | 0.21 | 0.09 | |||||||||
| 女贞Ligustrum lucidum | 0.24 | 0.00 | 0.24 | 0.20 | 0.00 | 0.39 | ||||||||
| 朴树Celtis sinensis | 0.55 | 0.35 | 0.39 | 0.48 | 0.14 | 0.09 | 0.27 | |||||||
| 山莓Rubus corchorifolius | 0.69 | 0.77 | 0.39 | 0.61 | 0.17 | 0.23 | 0.21 | 0.40 | ||||||
| 蛇葡萄Ampelopsis glandulosa | 0.21 | 0.00 | 0.53 | 0.56 | 0.16 | 0.08 | 0.46 | 0.29 | 0.23 | |||||
| 铁仔Myrsine africana | 0.35 | 0.30 | 0.61 | 0.77 | 0.22 | 0.05 | 0.00 | 0.25 | 0.39 | 0.33 | ||||
| 烟管荚蒾Viburnum utile | 0.59 | 0.21 | 0.46 | 0.49 | 0.17 | 0.29 | 0.49 | 0.45 | 0.30 | 0.50 | 0.27 | |||
| 野柿Diospyros kaki | 0.19 | 0.19 | 0.56 | 0.46 | 0.05 | 0.09 | 0.00 | 0.16 | 0.22 | 0.49 | 0.57 | 0.11 | ||
| 油茶Camellia oleifera | 0.26 | 0.08 | 0.49 | 0.44 | 0.31 | 0.06 | 0.02 | 0.11 | 0.12 | 0.19 | 0.40 | 0.21 | 0.21 | |
| 樟Camphora officinarum | 0.52 | 0.38 | 0.32 | 0.26 | 0.00 | 0.41 | 0.51 | 0.36 | 0.49 | 0.20 | 0.06 | 0.34 | 0.12 | 0.08 |
| 物种 Species |
臭鸡矢藤 Paederia cruddasiana |
淡竹叶 Lophatherum gracile |
地果 Ficus tikoua |
地锦 Parthenocissus tricuspidata |
狗脊 Woodwardia japonica |
贵州鳞毛蕨 Dryopteris wallichiana var kweichowicola |
寒莓 Rubus buergeri |
芒 Miscanthus sinensis |
芒萁 Dicranopteris pedata |
毛蕨 Cyclosorus interruptus |
牛膝 Achyranthes bidentata |
五叶地锦 Parthenocissus quinquefolia |
三脉紫菀 Aster ageratoides |
三叶地锦 Parthenocissus semicordata |
| 淡竹叶Lophatherum gracile | 0.47 | |||||||||||||
| 地果Ficus tikoua | 0.00 | 0.20 | ||||||||||||
| 地锦Parthenocissus tricuspidata | 0.48 | 0.05 | 0.30 | |||||||||||
| 狗脊Woodwardia japonica | 0.13 | 0.04 | 0.46 | 0.66 | ||||||||||
| 贵州鳞毛蕨 Dryopteris wallichiana var kweichowicola |
0.54 | 0.48 | 0.31 | 0.44 | 0.47 | |||||||||
| 寒莓Rubus buergeri | 0.26 | 0.56 | 0.19 | 0.13 | 0.12 | 0.45 | ||||||||
| 芒Miscanthus sinensis | 0.43 | 0.31 | 0.22 | 0.44 | 0.42 | 0.58 | 0.64 | |||||||
| 芒萁Dicranopteris pedata | 0.39 | 0.47 | 0.28 | 0.37 | 0.38 | 0.43 | 0.27 | 0.33 | ||||||
| 毛蕨Cyclosorus interruptus | 0.38 | 0.18 | 0.29 | 0.52 | 0.70 | 0.54 | 0.06 | 0.49 | 0.20 | |||||
| 牛膝Achyranthes bidentata | 0.32 | 0.04 | 0.23 | 0.45 | 0.69 | 0.33 | 0.04 | 0.29 | 0.25 | 0.59 | ||||
| 五叶地锦 Parthenocissus quinquefolia |
0.22 | 0.35 | 0.00 | 0.00 | 0.00 | 0.29 | 0.00 | 0.00 | 0.30 | 0.00 | 0.00 | |||
| 三脉紫菀Aster ageratoides | 0.21 | 0.08 | 0.26 | 0.55 | 0.70 | 0.42 | 0.19 | 0.26 | 0.15 | 0.56 | 0.71 | 0.00 | ||
| 三叶地锦 Parthenocissus semicordata |
0.52 | 0.41 | 0.00 | 0.09 | 0.12 | 0.36 | 0.06 | 0.04 | 0.37 | 0.39 | 0.28 | 0.33 | 0.23 | |
| 天名精Carpesium abrotanoides | 0.06 | 0.00 | 0.05 | 0.21 | 0.36 | 0.45 | 0.17 | 0.16 | 0.03 | 0.27 | 0.12 | 0.00 | 0.47 | 0.00 |
乔木层、灌木层、草本层和群落总体的模拟曲线的R2值分别为0.97、0.99、0.99、0.99, 回归方程极显著(P<0.01), 物种累积倒数百分比与累积相对频度比值分别为30.16/69.84、46.84/53.16、45.27/54.73、46.91/53.09, 均远离20/80, 表明马尾松群落无论是群落整体还是群落各垂直层次均处于不稳定状态, 进一步验证了种间联结性分析结果(图 7)。

|
| 图 7 马尾松群落各层次群落稳定性 Fig. 7 Community stability at all levels of the Pinus massoniana community |
种间联结作为衡量种间关系和群落结构稳定性的指标, 在植物群落分类和演替预测方面具有重要的作用[44]。一般而言, 在植物群落达到演替顶级时, 群落中的物种能最大化地利用资源, 生态位分化显著, 群落结构趋于完整和平衡, 总体物种呈正关联[45]。本研究对黔中喀斯特地区马尾松群落种间总体关联性分析发现, 该群落乔木层、灌木层和草本层主要物种间的总体关联性均表现为不显著正关联(P>0.05), 表明该群落中的物种之间存在一定程度的正相关关系, 但关联性较弱, 这与江焕等[46]、简小枚等[47]和杨青青等[48]的研究发现基本一致。
χ2检验能够较客观、准确地评估种对之间连接的显著性, 但不可避免地会损失一些植物多度等信息[40], 而Spearman秩相关检验则基于数量数据, 能够有效地补充和完善χ2检验的结果。本研究通过这两种方法分析马尾松群落主要物种之间的种间关联性和种间相关性, 结果在总体上一致, 但也存在差异。马尾松群落不同层次(乔木层、灌木层、草本层)主要物种的种间关联性整体表现为种对间不显著关联或相关性不强, 不同层次的物种之间并未出现显著的竞争或者互惠关系, 这可能与物种的资源利用方式、生态位分化以及环境因素和干扰程度有关[49—50]。例如乔木层的栗和油茶、构和朴树、杨梅和油茶、马尾松和杨梅等, 灌木层的樟和山莓、山莓和构、野柿和铁仔、栗和铁仔以及草本层的臭鸡屎藤和三叶地锦、毛蕨和三脉紫菀、狗脊和三脉紫菀、三脉紫菀和牛膝等种对间呈极显著或显著正相关, 反映了这些物种对喀斯特地区环境的适应。相反, 有的种对的种间关系呈现为极显著或显著负相关, 如乔木层的栗和朴树、马尾松和柃木, 灌木层的楤木和槲栎、山莓和油茶以及草本层的芒和三叶地锦、淡竹叶和狗脊等, 这与物种的生物学特性和对特定生境的适应能力有关[51]。这些发现表明不同的物种在群落中具有不同的功能和生态位特征, 在群落中所扮演的角色也不尽相同。
3.2 马尾松群落主要物种生态位特征生态位宽度越大, 对资源的竞争力越大, 分布范围越广[50]。本研究中, 马尾松是研究区群落的绝对优势种, 占据最大的生态位宽度, 能够更充分地利用环境资源, 适应环境的能力更强。乔木层的油茶、栗、槲栎, 灌木层的栗、槲栎、楤木, 草本层的芒萁、贵州鳞毛蕨、臭鸡矢藤也占据了较大的生态位宽度, 在群落演替中也能获得较大的优势地位, 而相对较小的生态位宽度的物种适应了不同于其他优势物种的栖息地条件, 从而促进了物种共存[52]。
生态位重叠可以反映物种间的竞争或共存关系, 以及对环境变化的适应能力。一般认为, 如果重叠值大于0.5, 则表示物种之间存在明显的生态和资源利用相似性, 从而导致竞争加剧[7]。在本研究中, 马尾松群落主要优势物种的生态位重叠值大多低于0.5, 表明物种之间对环境资源需求的相似性较小, 种间竞争较弱, 种间关系较为稳定, 这与滇东菌子山喀斯特生境的研究结果相同[53], 但与滇中云南杨梅灌丛的研究结果相反[54]。油茶和栗(0.71)、构和朴树(0.71)、杨梅和油茶(0.67)以及杨梅和栗(0.59)的生态位重叠程度较大, 可能是因为这些物种生态位较宽以及重要值较大。此外, 一些物种的生态位重叠值为0, 例如, 乔木层的构和槲栎、栗和楤木、亮叶桦和柃木等, 灌木层的构和女贞、柃木和女贞、柃木和樟等, 草本层的五叶地锦和地锦、臭鸡矢藤和地果、三叶地锦和天名精等, 可能是由于分布不均匀或者物种稀疏, 在小的生境范围内没有重叠, 以不同的方式适应生境[55]。大多数研究表明, 生态位宽度较大的物种生态位重叠相对较高, 而生态位宽度较小的物种生态位重叠较小[56]。但本研究中生态位宽度大的物种, 不一定存在较高的生态位重叠, 例如乔木层的马尾松和油茶生态位宽度大, 但生态位重叠并不高, 这与胡正华等[57]、姚珊等[52]的研究结果一致, 说明生态位宽度与生态位重叠之间存在复杂的关系, 并不总是成正比关系, 种群内也存在互利性生态位重叠现象[58]。
3.3 马尾松群落稳定性种间联结性和群落稳定性之间存在紧密的联系[59]。为验证种间联结性对群落稳定性预测的准确性, 本研究利用M.Godron法对马尾松群落的乔木层、灌木层和草本层以及群落整体进行群落稳定性分析, 发现该群落不论是整体还是各垂直层次均处于不稳定状态, 表明其物种组成和数量结构在不断调节中。与张乐满等[13]和李锦婷等[60]在三峡水库消落带、贵州赤水的小黄花茶群落的相关研究结果基本一致。这可能与喀斯特地区独特的生境条件有关, 岩石裸露率大, 土层浅薄且分布不均, 生境异质性高[61];此外, 人类活动干扰、气候变化等外部因素也可能对马尾松群落的稳定性产生影响[62]。
总体而言, 通过对马尾松群落主要物种的种间联结、生态位宽度与重叠度及群落稳定性的综合研究, 能够深入了解黔中喀斯特山地典型马尾松群落的类型、结构以及生物多样性维持机制。此外, 对于喀斯特山地植被恢复与重建、森林经营与管理以及生物多样性保护具有重要的理论和实践意义。本研究结果表明研究区马尾松群落主要物种种间关联性弱, 群落稳定性不高。因此, 在喀斯特山地森林质量精准提升的过程中, 应着重选择生态位宽、生态位重叠小, 且能够与现有优势物种相互促进具有生态效益的物种, 增加林分的异质性, 将马尾松人工纯林逐步转变为多树种、异龄、复层的针阔混交林, 提高马尾松群落稳定性[63]。此外, 由于种间关联性只能揭示种间和种内竞争的结果或现状, 而未揭示其背后的生态学过程和机制[10]。因此, 为了全面了解群落物种共存机制以及生态影响因素, 今后亟需综合运用群落生态学、植物生理学、遗传生态学等学科的理论方法, 进一步深入研究不同生态因子对马尾松群落的种间关系和稳定性的影响, 科学揭示种间关联性的生态学机制, 以丰富和夯实喀斯特地区典型森林生态系统群落构建和物种共存机制的理论基础[10, 52]。
4 结论黔中喀斯特地区典型马尾松群落内各垂直层次(乔木层、灌木层、草本层)主要物种间的总体关联性均呈不显著正关联;61.90%—80.95%的种对呈不显著关联/不显著相关, 种间关联松散且相关性较弱, 各物种间呈相对独立分布格局;群落总体稳定性较弱(46.91/53.09)。在喀斯特地区植被恢复与重建过程中, 应尽可能选择生态习性和生物学特性相近、生态位重叠程度低、种间正关联较强的乔灌草物种。例如, 乔木层的栗和油茶、灌木层的樟和山莓、草本层的臭鸡屎藤和三叶地锦的搭配等。通过适当调节和优化各层次物种的组合, 有助于增强群落的垂直结构多样性和功能复杂性, 使植物充分利用资源, 提升群落内物种共存水平和群落稳定性。
| [1] |
刘翔宇, 何东, 田文斌, 宋彦君, 尹芳, 许洺山, 程浚洋, 阎恩荣. 浙江普陀山岛森林木本植物的种间关联格局. 植物生态学报, 2017, 41(12): 1219-1227. |
| [2] |
Gleason H A. The individualistic concept of the plant association. Bulletin of the Torrey Botanical Club, 1926, 53(1): 7. DOI:10.2307/2479933 |
| [3] |
徐满厚, 刘敏, 翟大彤, 刘彤. 植物种间联结研究内容与方法评述. 生态学报, 2016, 36(24): 8224-8233. |
| [4] |
Hutchinson G. Concluding remarks-cold spring harbor symposia on quantitative biology. Bulletin of Mathematical Biology, 1957, 53(1507): 193-213. |
| [5] |
王仁忠. 放牧影响下羊草草地主要植物种群生态位宽度与生态位重叠的研究. 植物生态学报, 1997, 21(4): 304-311. |
| [6] |
刘巍, 曹伟. 长白山云冷杉群落主要种群生态位特征. 生态学杂志, 2011, 30(8): 1766-1774. |
| [7] |
马姜明, 李昆. 森林生态系统稳定性研究的现状与趋势. 世界林业研究, 2004, 17(1): 15-19. |
| [8] |
李帅锋, 郎学东, 黄小波, 刘万德, 苏建荣, 徐崇华, 李智宏, 徐凡迪. 藤枣生境地木本植物种间关联性与群落稳定性. 生物多样性, 2020, 28(3): 350-357. |
| [9] |
张滋芳, 毕润成, 张钦弟, 许强. 珍稀濒危植物矮牡丹生存群落优势种种间联结性及群落稳定性. 应用与环境生物学报, 2019, 25(2): 291-299. |
| [10] |
涂洪润, 农娟丽, 朱军, 赵佳佳, 杨婉琳, 朱琪琪, 谢彦军, 刘润红. 桂林岩溶石山密花树群落主要物种的种间关联及群落稳定性. 生态学报, 2022, 42(9): 3688-3705. |
| [11] |
史浩伯, 陈亚宁, 李卫红, 孙桂丽. 塔里木河下游植被种间关系与稳定性分析. 干旱区研究, 2020, 37(1): 220-226. |
| [12] |
覃弦, 龙翠玲, 李娟. 茂兰喀斯特森林不同演替阶段优势乔木种群生态位研究. 西南大学学报: 自然科学版, 2017, 39(6): 42-49. |
| [13] |
张乐满, 兰波, 张东升, 刘英杰, 张丽红, 许文锋, 段晨辉, 敦静怡, 刘正学. 三峡水库涪陵-奉节段消落带优势草本植物生态位与种间联结性研究. 生态学报, 2022, 42(8): 3228-3240. |
| [14] |
薛卫星, 李春辉, 艾训儒, 姚兰, 郭秋菊, 朱江. 鹅掌楸天然林优势树种生态位与种间联结性. 森林与环境学报, 2023, 43(1): 26-34. |
| [15] |
丁茂, 汪宇坤, 何煜然, 徐晓阳, 周守标, 王智. 安徽鹞落坪国家级自然保护区落叶阔叶林树种多样性、种间联结及群落稳定性变化. 生态学报, 2023, 43(7): 2818-2830. |
| [16] |
Thevathasan N V, Gordon A M. Ecology of tree intercropping systems in the north temperate region: experiences from southern Ontario, Canada. Agroforestry Systems, 2004, 61(1): 257-268. |
| [17] |
宋永昌, 陈小勇. 中国东部常绿阔叶林生态系统退化机制与生态恢复. 北京: 科学出版社, 2007.
|
| [18] |
简尊吉, 倪妍妍, 徐瑾, 雷蕾, 曾立雄, 肖文发. 中国马尾松林土壤肥力特征. 生态学报, 2021, 41(13): 5279-5288. |
| [19] |
孟祥江, 何邦亮, 马正锐, 侯元兆, 李玉敏. 我国马尾松林经营现状及近自然育林探索. 世界林业研究, 2018, 31(3): 63-67. |
| [20] |
李瑞玲, 王世杰, 张殿发. 贵州喀斯特地区生态环境恶化的人为因素分析. 矿物岩石地球化学通报, 2002, 21(1): 43-47. |
| [21] |
王世杰. 喀斯特石漠化概念演绎及其科学内涵的探讨. 中国岩溶, 2002, 21(2): 101-105. |
| [22] |
何斌, 李青, 陈群利, 薛晓辉, 李望军. 贵州省西北部马尾松人工林种群数量特征与动态. 中南林业科技大学学报, 2020, 40(11): 129-137, 155. |
| [23] |
高祥, 丁贵杰, 翟帅帅, 陈模芳, 杜华东. 不同林分密度马尾松人工林根系生物量及空间分布研究. 中南林业科技大学学报, 2014, 34(6): 71-75. |
| [24] |
金鹏, 徐明, 文春玉, 聂坤, 杨雪, 唐雪娅, 魏珊, 张健. 黔中地区马尾松-甜槠针阔混交林群落结构特征与林木空间分布格局及其关联性. 西北林学院学报: 1-16. [2023-10-27]. http://kns.cnki.net/kcms/detail/61.1202.S.20230914.1638.002.html.
|
| [25] |
肖集泓, 赖敏英, 李媚, 王春晓, 钟址非, 温清燕, 钱志军, 罗真, 王迎雪, 邓清雅, 邓传远. 宁德三都澳海岛马尾松林群落特征及物种多样性. 生态学报, 2023(23): 1-17. [2023-10-27]. https://doi.org/10.20103/j.stxb.202211153300.
|
| [26] |
曾林辉, 周蕾, 寇亮, 迟永刚. 基于Dendrometer监测的亚热带地区马尾松多时间尺度径向变化及其与环境因子的关系. 生态学报, 2023(16): 1-12. [2023-10-27]. http://kns.cnki.net/kcms/detail/11.2031.Q.20230606.0923.001.html.
|
| [27] |
陈斯, 刘道聪, 林勇, 黄伟, 姚兰, 艾训儒. 不同海拔梯度马尾松天然群落结构特征研究. 湖北民族学院学报: 自然科学版, 2017, 35(3): 247-254. |
| [28] |
熊能, 金则新, 陈琢. 千岛湖姥山马尾松种群结构和分布格局研究. 植物研究, 2010, 30(5): 537-542. |
| [29] |
郭丽玲, 潘萍, 欧阳勋志, 宁金魁, 臧颢, 刘苑秋, 杨阳, 桂亚可. 赣南马尾松天然林不同生长阶段碳密度分布特征. 北京林业大学学报, 2018, 40(1): 37-45. |
| [30] |
王秀云, 朱汤军, 徐高福. 千岛湖马尾松群落种群生态位与种间联结. 浙江林业科技, 2015, 35(3): 36-42. |
| [31] |
刘鹏, 张合平. 马尾松群落的种间联结性研究. 湖南林业科技, 2017, 44(5): 22-28. |
| [32] |
黎露, 马姜明, 莫燕华, 王永琪, 黄柳欣, 陆绍浩. 广西不同林龄马尾松人工林林下木本植物种间联结动态. 广西科学, 2022, 29(6): 1160-1168. |
| [33] |
张育林, 程星, 王恒松, 张祥祥, 吴伟, 张辉华. 贵阳市乌当区北部地下水资源特征与开发利用研究. 贵州科学, 2018, 36(6): 74-80. |
| [34] |
黄世芹, 刘英春, 李宗参, 李杨. 2010-2019年贵阳市乌当区气候变化特征分析. 现代农业科技, 2020(9): 197-200, 204. |
| [35] |
李苇洁, 易朝阳, 陈志华, 段旭. 贵阳二环林带不同林地土壤养分状况研究. 贵州科学, 2007, 25(3): 28-32. |
| [36] |
范明阳. 黄山松林与马尾松林群落特征与植被分类初探[D]. 南京: 南京林业大学, 2023.
|
| [37] |
Baert J M, Janssen C R, Sabbe K, De Laender F. Per capita interactions and stress tolerance drive stress-induced changes in biodiversity effects on ecosystem functions. Nature Communications, 2016, 7: 12486. |
| [38] |
Gu L, Gong Z W, Li W Z. Niches and interspecific associations of dominant populations in three changed stages of natural secondary forests on Loess Plateau, P.R. China. Scientific Reports, 2017, 7: 6604. |
| [39] |
Schluter D. A variance test for detecting species associations, with some example applications. Ecology, 1984, 65(3): 998-1005. |
| [40] |
张金屯. 数量生态学 (3版). 北京: 科学出版社, 2018.
|
| [41] |
Levins R. Evolution in Changing Environments: Some Theoretical Explorations. Princeton: Princeton University Press, 1968.
|
| [42] |
Pianka E R. The structure of lizard communities. Annual Review of Ecology and Systematics, 1973, 4: 53-74. |
| [43] |
郑元润. 森林群落稳定性研究方法初探. 林业科学, 2000, 36(5): 28-32. |
| [44] |
Wu S N, Wen L, Dong S K, Gao X X, Xu Y D, Li S, Dong Q M, Wessell K. The plant interspecific association in the revegetated alpine grasslands determines the productivity stability of plant community across restoration time on Qinghai-Tibetan Plateau. Frontiers in Plant Science, 2022, 13: 850854. |
| [45] |
刘文婷, 张金峰, 刘琪璟. 长白山东坡白桦-长白落叶松林优势种群结构与动态特征. 生态学报, 2023, 43(18): 7462-7473. |
| [46] |
江焕, 张辉, 龙文兴, 方燕山, 符明期, 朱孔新. 金钟藤入侵群落的种间联结及生态位特征. 生物多样性, 2019, 27(4): 388-399. |
| [47] |
简小枚, 税伟, 王亚楠, 王前锋, 陈毅萍, 江聪, 项子源. 重度退化的喀斯特天坑草地物种多样性及群落稳定性——以云南沾益退化天坑为例. 生态学报, 2018, 38(13): 4704-4714. |
| [48] |
杨青青, 杨众养, 陈小花, 余雪标, 薛杨, 王小燕. 热带海岸香蒲桃天然次生林群落优势种群种间联结性. 林业科学, 2017, 53(9): 105-113. |
| [49] |
孙玉真, 王志泰, 包玉, 魏文飞, 杨兴艺. 城市遗存山体植被群落特征对不同人为干扰方式及强度的响应. 生态学报, 2023, 43(11): 4632-4650. |
| [50] |
卢炜丽, 张洪江, 陈奇伯, 吴霖, 栗宏林. 重庆四面山杉木林群落乔木层优势种群生态位特征研究. 西北林学院学报, 2016, 31(2): 60-65. |
| [51] |
姚珊. 贵州孟关林场马尾松人工林群落动态及灌草组成分析[D]. 贵阳: 贵州大学, 2022.
|
| [52] |
姚珊, 郭其强, 盘金文, 张亚琴, 罗丝琼. 马尾松人工林发育过程中林下灌草群落优势种生态位动态分析. 西北植物学报, 2021, 41(8): 1417-1424. |
| [53] |
刘雨婷, 侯满福, 贺露炎, 唐伟, 赵俊. 滇东菌子山喀斯特森林群落乔木优势树种生态位和种间联结. 应用生态学报, 2023, 34(7): 1771-1778. |
| [54] |
张零念, 朱贵青, 杨宽, 刘星月, 巩合德, 郑丽. 滇中云南杨梅灌丛主要木本植物生态位与种间联结. 植物生态学报, 2022, 46(11): 1400-1410. |
| [55] |
Coudun C, Gégout J C. Ecological behaviour of herbaceous forest species along a pH gradient: a comparison between oceanic and semicontinental regions in northern France. Global Ecology and Biogeography, 2005, 14(3): 263-270. |
| [56] |
刘彤, 李云灵, 周志强, 胡海清. 天然东北红豆杉(Taxus cuspidata)种内和种间竞争. 生态学报, 2007, 27(3): 924-929. |
| [57] |
胡正华, 吴芳芳, 刘巧辉, 陈书涛, 陈魁, 于明坚. 古田山国家自然保护区马尾松林主要种群生态位研究. 林业科学研究, 2009, 22(3): 330-334. |
| [58] |
俞昀, 白小军, 王志一. 大兴安岭次生林区不同龄级落叶松(Larix gmelinii)生态位特征和竞争关系. 生态学报, 2022, 42(12): 4912-4921. |
| [59] |
黄祥童, 王绍先, 黄炳军, 尹航, 崔凯峰, 赵伟, 范宇光, 顾德峰. 珍稀植物对开蕨与其伴生物种的联结性及群落稳定性. 生态学报, 2015, 35(1): 80-90. |
| [60] |
李锦婷, 穆君, 申开平, 郭云, 白小节, 臧丽鹏, 李青, 韩勖, 赵颖, 何跃军. 小黄花茶群落优势木本植物生态位与种间联结性. 生态学报: 1-12. [2023-10-27]. https://doi.org/10.20103/j.stxb.202212273676.
|
| [61] |
张忠华, 胡刚. 喀斯特山地青冈栎群落优势种的种间关系分析. 生态环境学报, 2011, 20(S2): 1209-1213. |
| [62] |
尹锴, 崔胜辉, 石龙宇, 吝涛, 郭青海, 吕静. 人为干扰对城市森林灌草层植物多样性的影响——以厦门市为例. 生态学报, 2009, 29(2): 563-572. |
| [63] |
吴瑶, 廖彩霞. 近自然林业的研究. 林业科技情报, 2007, 39(2): 1-2. |