 2024, Vol. 44
2024, Vol. 44文章信息
- 王春丽, 陈峰, 蒋日进, 张洪亮, 朱文斌, 朱凯
- WANG Chunli, CHEN Feng, JIANG Rijin, ZHANG Hongliang, ZHU Wenbin, ZHU Kai
- 典型气候事件对浙江近海底层鱼类时空分布的影响
- Effects of typical climate events on spatio-temporal distribution of bottom fish in the coastal waters of Zhejiang
- 生态学报. 2024, 44(10): 4231-4243
- Acta Ecologica Sinica. 2024, 44(10): 4231-4243
- http://dx.doi.org/10.20103/j.stxb.202305171043
-
文章历史
- 收稿日期: 2023-05-17
- 网络出版日期: 2024-02-28






2. 浙江省海洋水产研究所, 农业农村部重点渔场渔业资源科学观测实验站, 浙江省海洋渔业资源可持续利用技术研究重点实验室, 舟山 316021
2. Key Laboratory of Sustainable Utilization of Technology Research for Fishery Resources of Zhejiang Province, Scientific Observing and Experimental Station of Fishery Resources for Key Fishing Grounds, Ministry of Agriculture and Rural Affairs, Zhejiang Marine Fisheries Research Institute, Zhoushan 316021, China
气候变化对海洋生态系统的影响越来越受到国内外学者的重视, 其不仅引起大范围的海洋物理和化学性质的改变, 也对物种的变迁和更替产生显著的影响[1—2]。作为全球气候变化的典型, 厄尔尼诺及拉尼娜事件也对我国浙江近海鱼类生物量及其时空分布产生了一定的影响。厄尔尼诺现象又称厄尔尼诺海流, 是太平洋赤道带大范围内海洋和大气相互作用后失去平衡而产生的一种气候现象, 会导致赤道太平洋东部和中部海面温度持续异常偏暖, 拉尼娜恰好与之相反[3]。虽然厄尔尼诺拉尼娜是太平洋赤道带海域发生的周期性海面温度变化, 但也会通过大气遥相应对我国近海海洋环境产生一定的影响:厄尔尼诺发生当年中国近海及邻近海域海温偏低, 次年海温偏高, 为暖年[4—6]。厄尔尼诺发生当年海表温度距平值处于较低水平, 发生次年海表温度距平明显升高, 结束后一年又有所下降;拉尼娜变动规律与之相反[7];厄尔尼诺发生时, 闽浙沿岸表层盐度较低[8]。这些变化会引起鱼类生物量和栖息地的改变, 间接影响着渔业生产。
浙江沿岸海域岛屿众多, 长期受长江冲淡水、台湾暖流、浙江沿岸流等海流和水系的综合影响, 该海域营养盐类和饵料生物丰富[9], 是各种海洋生物繁殖、索饵、育肥等天然栖息场所, 生物多样性较高[10], 为我国重要的渔场之一[11]。该海域的水温、盐度等参数变化较大, 表层平均水温以4月最低、12月次之, 9月最高、6月次高, 平均水温在19.9℃左右, 最大时空温差17.56℃[10]。近海是众多经济种类的主要产卵场和索饵场[12], 外海则是主要的越冬场[13]。关于浙江近海海域的渔获种类组成、区系分布、与环境因子间的关系等方面已有众多相关研究[14—16], 但国内关于气候变化对鱼类资源量变动、栖息地变动的影响多为单一鱼种, 鲜少有关于气候变化对鱼类群落变动的研究:厄尔尼诺的发生有利于带鱼渔获量的增加[8], 但会对鲐鲹鱼渔获量产生不利影响[17];拉尼娜事件不利于鲐鱼资源[18]。当ENSO(El Niño-Southern Oscillation)处于发展阶段时, 其指数与本年度的中型秋刀鱼和鲐鱼单位捕捞努力量渔获量(Catch Per Unit of Effort, CPUE)呈负相关, 与次年大型秋刀鱼和鳀鱼的丰度指数呈正相关[19];太平洋年代际振荡(Pacific Decadal Oscillation, PDO)暖期内的秋刀鱼CPUE一般高于PDO冷期[20]。除了会影响渔获量, 厄尔尼诺拉尼娜对鱼类时空分布也会产生一定影响:在大部分厄尔尼诺年, 黄鳍金枪鱼围网作业重心会向东、向北移;而拉尼娜年则向西移[21], 厄尔尼诺强度与鲐鱼渔场适宜栖息地面积呈负相关[22], 且在外界温度增加的影响下, 鲐鱼的栖息地由北纬25°N转移到北纬38°N, 有向北迁徙的趋势[23]。有关近年来研究大部分基于局部海域或为单一鱼种, 本文基于近10年浙江近海底拖网调查资料, 对不同气候事件下整个浙江近海鱼类生物量及其时空分布变化进行研究, 探讨厄尔尼诺、拉尼娜对资源变动的影响, 以期在气候变化的大背景下为东海鱼类资源的科学管理及可持续利用提供理论依据。
1 材料与方法 1.1 数据来源 1.1.1 渔业数据本研究渔业资源调查数据来自浙江省海洋水产研究所2013—2022年浙江近海底拖网调查项目, 调查海域为27°—31°N, 120.5°—124°E, 调查站点以15'×15'进行布设(图 1)。调查时间为每年的春季(4月)和秋季(11月), 部分年份其他季节也有进行调查。调查网具网口拉紧周长为25 m, 囊网网目尺寸为2.0cm, 浙北、浙南站位调查网口宽9.9m, 浙中站位调查网口宽6.6m;记录内容包括拖网站位、起放网时间和位置、拖速和拖网时间等, 拖网结束后, 将渔获样品带回实验室进行种类鉴定、生物学测定。生物学测定包括渔获名称、尾数、重量等。调查期间共鉴定出鱼类226种, 主要为底层及近底层鱼类, 占90.7%;冷水性鱼类5种, 热带鱼类1种, 其他均为暖水性或暖温性鱼类。春季优势种为龙头鱼、小黄鱼、矛尾虾虎鱼、细条天竺鲷;秋季优势种为龙头鱼、细条天竺鲷、七星底灯鱼、棘头梅童鱼、鮸鱼、矛尾虾虎鱼;其中龙头鱼为调查海域调查期间的绝对优势种。

|
| 图 1 浙江近海渔业资源调查站位图 Fig. 1 Survey site map of Zhejiang offshore fishery resources |
本研究选取海表温度(SST)、海表温度距平(SSTA)、叶绿素a浓度(Chl-a)作为环境变量的指标, SST、SSTA、Chl-a遥感环境资料来源于美国国家海洋和大气管理局(NOAA)数据库(https://oceanwatch.pifsc.noaa.gov/erddap/griddap/index.html)。
厄尔尼诺/拉尼娜事件采用Nino3.4区尼诺指数(Ocean Nino Index, ONI)来表征, 时间分辨率为月, 数据来源于NOAA气候预报中心(https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.phpg1)。当Nino3.4区指数3个月滑动平均≥0.5℃, 且持续至少五个月, 则判定为一次厄尔尼诺事件;当Nino3.4区指数3个月滑动平均≤-0.5℃, 且持续至少五个月, 则判定为一次拉尼娜事件。
1.2 研究方法统计分析近10年气候事件发生频次, 选取生物量作为浙江近海鱼类资源丰度指标, 使用SPSS软件利用双独立样本T检验分析不同气候事件下春、秋两季浙江近海鱼类生物量差异, 分析ONI、海洋环境因子(SST、SSTA、Chl-a)与鱼类生物量相关性, P<0.05判定为具有显著差异性或相关性。探究不同海洋环境因子对浙江近海鱼类生物量的影响, 及对气候事件响应的滞后性。
鱼类的生物量估算采用扫海面积法[11], 计算公式如下:
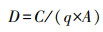
|
(1) |
式中D为生物量(kg/km2);C为每小时取样面积内的渔获量(kg);q为网具渔获率, 本文中q取0.5;A为网具每小时扫海面积(km2)。
交叉小波可以直观地对气象和水文要素之间的相关性进行分析[24], 使用Matlab软件采用交叉小波分析探究ONI对海洋环境因子SST、SSTA、Chl-a及生物量变动的影响, 计算两者的交叉小波谱及频域变化周期, 进而分析两个时间序列在不同时域内的滞后性特征及相关性。文中分析所使用的交叉变换工具包括小波功率谱(Cross Wavelet, XWT) 和小波凝聚谱(Wavelet Transform Coherence, WTC)。假设两时间序列分别为X={xi|i=1, 2, …, n}与Y={yi|i=1, 2, …, n}, 连续小波变换分别为WnX(s)和WnY(s), 则交叉小波谱定义[25—27]为:
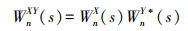
|
(2) |
式中, WnY*(s)为WnX(s)的复共轭, 交叉小波功率谱为WnXY(s)的绝对值。
WTC定义为[28]:
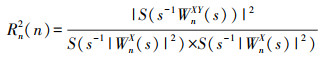
|
(3) |
式中, s为尺度, S为平滑器。
利用ArcGIS空间分析模块, 绘制浙江近海鱼类生物量在不同气候事件下SST、SSTA、Chl-a分布图, 分析厄尔尼诺、拉尼娜事件下海洋环境因子与鱼类生物量的时空分布差异。
2 结果 2.1 气候事件及海洋环境变化2013—2022年近10年间, 全球发生过2次厄尔尼诺事件(2014年10月—2016年4月、2018年9月—2019年6月), 4次拉尼娜事件(2016年8月—2016年12月、2017年10月—2018年4月、2020年8月—2021年5月、2021年8月—2022年6月)(表 1)。其中2014年开始的厄尔尼诺事件持续19个月之久。
| 年份 Year |
月份Month | |||||||||||
| 1月 Jan. |
2月 Feb. |
3月 Mar. |
4月 Apr. |
5月 May |
6月 Jun. |
7月 Jul. |
8月 Aug. |
9月 Sep. |
10月 Oct. |
11月 Nov. |
12月 Dec. |
|
| 2013 | N | N | N | N | N | N | N | N | N | N | N | N |
| 2014 | N | N | N | N | N | N | N | N | N | E | E | E |
| 2015 | E | E | E | E | E | E | E | E | E | E | E | E |
| 2016 | E | E | E | E | N | N | N | L | L | L | L | L |
| 2017 | N | N | N | N | N | N | N | N | N | L | L | L |
| 2018 | L | L | L | L | N | N | N | N | E | E | E | E |
| 2019 | E | E | E | E | E | E | N | N | N | N | N | N |
| 2020 | N | N | N | N | N | N | N | L | L | L | L | L |
| 2021 | L | L | L | L | L | N | N | L | L | L | L | L |
| 2022 | L | L | L | L | L | L | ||||||
| E、L和N分别代表发生厄尔尼诺、拉尼娜事件以及正常气候 | ||||||||||||
浙江近海近10年间SST、Chl-a、SSTA波动明显(图 2—图 4), 2013年11月至2015年11月, ONI呈现逐步上升的趋势, 气候从正常年份向厄尔尼诺转变;2016—2020年, 气候处于厄尔尼诺和正常年份的波动状态, 2020年8月之后, 气候基本处于拉尼娜发生期。浙江近海ONI与SST呈近乎正相波动, 2014年3月为10年间的最低值, 2020年8月为10年间的最高值;SSTA在2016年4月之前, 与ONI呈正相变化, 而在2016年之后, 与ONI呈近乎负相关变动, 2019年11月之前, SSTA呈阶梯上升趋势;之后SSTA月际间跨度较大, 相较于之前变动较为异常;Chl-a在2014年3月—2016年5月之间呈负相关变动, 2015年5月—2015年8月ONI上升到峰值, 而此时Chl-a处于较低水平, 2017年11月—2019年11月也处于近10年间的较低水平, 2020年4月份之后, Chl-a同ONI呈正相关变动, 4月份远高于11月份。

|
| 图 2 浙江近海2013—2022年ONI与SST Fig. 2 Changes in ONI and SST in Zhejiang coastal waters from 2013 to 2022 |

|
| 图 3 浙江近海2013—2022年ONI与SSTA Fig. 3 Changes in ONI and SST in Zhejiang coastal waters from 2013 to 2022 |

|
| 图 4 浙江近海2013—2022年ONI与Chl-a Fig. 4 Changes in ONI and SST in Zhejiang coastal waters from 2013 to 2022 |
对比分析发现, SST、Chl-a、SSTA均会有一段时期在厄尔尼诺发生后次年同月份升高, 拉尼娜发生后次年同月份降低, 其中SST在整个研究期间呈此规律, 受其他因素的影响, 个别月份温度变化不明显;Chl-a变动在2017年之后更明显。
2.2 生物量变动及相关性近10年来, 浙江近海春、秋季生物量在不同事件发生期间差异明显(图 5), 春季近10年平均生物量为567.81kg/km2, 其中正常年份平均生物量为768.72kg/km2, 厄尔尼诺事件发生期间平均生物量为486.76kg/km2, 拉尼娜事件发生期间平均生物量为447.97kg/km2;秋季近10年平均生物量为754.46kg/km2, 其中正常年份平均生物量为849.46kg/km2, 厄尔尼诺事件发生期间平均生物量为661.31kg/km2, 拉尼娜事件发生期间平均生物量为776.83kg/km2。从不同事件发生期间生物量来看, 厄尔尼诺与拉尼娜发生时期春、秋两季生物量的平均值、中位数均小于正常年份期间(P<0.05), 其中拉尼娜事件期间春季平均生物量显著低于正常年份(P<0.05), 厄尔尼诺期间秋季生物量显著低于正常年份(P<0.01), 表明厄尔尼诺及拉尼娜事件均会对浙江近海生物量产生不利影响, 且对不同季节的影响程度不同。

|
| 图 5 2013—2022年不同气候事件期间春秋季生物量 Fig. 5 Biomass during different climatic events in spring and autumn from 2013 to 2022 |
对生物量与月平均SST、Chl-a、SSTA进行相关性检验, 结果表明生物量与月平均SST及Chl-a呈极显著正相关(P<0.01)(图 6—图 7), 与SSTA无显著性;相关性分析结果表明SST及Chl-a是影响浙江近海鱼类生物量的主要因素。

|
| 图 6 生物量与海表温度变化图 Fig. 6 Changes of biomass and SST |

|
| 图 7 生物量与叶绿素a变化图 Fig. 7 Changes of biomass and Chl-a |
利用交叉小波对ONI与SST、Chl-a及生物量进行分析, 计算两者的交叉小波谱。ONI与SST在时间局部的响应关系如图 8所示:在2013年11月—2022年4月存在一个明显的2—3.9个月频域同步, SST滞后ONI3个月;ONI与Chl-a在时间局部的响应关系为图 9:在2013年11月—2022年4月存在一个明显的1.7—3个月频域同步, Chl-a与ONI呈正相关;ONI与生物量在时间局部的响应关系为图 10:在2013年11月—2022年4月存在一个明显的3—3.6个月频域同步, 生物量滞后于ONI3个月;ONI与SSTA在研究期间不存在明显的频域同步。

|
| 图 8 SST同ONI的交叉小波功率谱 Fig. 8 Cross wavelet power spectrum of SST with ONI |

|
| 图 9 Chl-a同ONI的交叉小波功率谱 Fig. 9 Cross wavelet power spectrum of Chl-a with ONI |

|
| 图 10 生物量同ONI的交叉小波功率谱 Fig. 10 Cross wavelet power spectrum of biomass with ONI ↗、↙表示生物量/海表温度/叶绿素滞后厄尔尼诺拉尼娜事件且分别为正相关关系、负相关关系, ↘、↖表示生物量/海洋表面温度/叶绿素超前厄尔尼诺拉尼娜事件且分别为正相关关系、负相关关系 |
不同气候事件对不同季节海洋环境的影响不同:春季厄尔尼诺发生期间浙江近海SST整体较正常年份及拉尼娜期间为高, 相较于正常年份, 拉尼娜发生期暖水范围缩小, 厄尔尼诺期间冷水面积大幅缩小(图 11);Chl-a浓度在拉尼娜及厄尔尼诺期间均高于正常年份, 且近岸浓度远高于外海(图 12)。秋季SST以厄尔尼诺期间最高, 拉尼娜期间最低(图 13);近岸Chl-a浓度以厄尔尼诺期间最高, 且远高于拉尼娜和正常年份(图 14)。

|
| 图 11 不同气候事件生物量与海表温度分布图 Fig. 11 Distribution map of biomass and SST in different climate events |

|
| 图 12 不同气候事件生物量与叶绿素a分布图 Fig. 12 Distribution map of biomass and Chl-a in different climate events |

|
| 图 13 不同气候事件生物量与海表温度分布图 Fig. 13 Distribution map of biomass and SST in different climate events |

|
| 图 14 不同气候事件生物量与叶绿素a分布图 Fig. 14 Distribution map of biomass and Chl-a in different climate events |
对比不同气候事件发生期, 发现鱼类生物量、SST和Chl-a分布呈明显差异:正常年份春季鱼类分布相对均匀, 生物量高值分布区的SST和Chl-a浓度分别为14—16℃和5—9mg/m3;拉尼娜期间鱼类主要分布在30°N以南, 生物量高值分布区的SST和Chl-a浓度分别为15—17℃和2—4mg/m3;厄尔尼诺期间鱼类分布整体偏北;生物量高值分布区的SST和Chl-a浓度分别为14—16℃和3—6mg/m3。相对于正常年份, 厄尔尼诺及拉尼娜期春季鱼类整体分布远离近岸。
不同气候事件发生期秋季鱼类分布情况与春季不同, 秋季生物量高值分布区的SST为21—24℃;厄尔尼诺期鱼类向南洄游, 生物量高值分布区的Chl-a浓度为0—4mg/m3;拉尼娜期则向北洄游, 生物量高值分布区的Chl-a浓度为2—6mg/m3;正常年份生物量高值分布区的Chl-a浓度为0—4mg/m3;在适宜的SST及Chl-a浓度范围下, 鱼类群落向着更高的SST及Chl-a浓度的范围洄游聚集。相较于拉尼娜期, 厄尔尼诺发生期鱼类群落向近岸洄游。表明厄尔尼诺及拉尼娜事件不仅对鱼类群落在纬度上的分布产生影响, 还会对鱼类向近岸洄游的快慢产生影响。
3 讨论 3.1 典型气候事件对浙江近海海洋环境的影响海洋是气候系统储存能量的主要载体[29], 气候变化影响海洋环境物理特性(海表温度、海洋环流、海平面高度、盐度、垂直混合度和海冰厚度等海洋环境因素)[30], 这些变化是对全球气候变化的一种响应。近60年的历史数据显示, 厄尔尼诺指数和长江径流量存在一定的联系, 当厄尔尼诺指数达到峰值时, 同年或者稍微滞后长江径流量峰值就会出现, 长江径流量的谷值则容易出现在厄尔尼诺指数处于正负值之间转换时; 陆地径流的变化会改变陆源营养盐的输入进而改变甲藻孢囊丰度[31]。浙江近海海洋环境在气候事件发生期间波动明显, 其中SST在厄尔尼诺事件发生后次年升高, 拉尼娜与之相反, 与曹丛华[4]、张守文[5]、陈美榕等[6]的研究结果一致。浙江近海Chl-a呈近岸浓度远高于外海分布, 与历史研究结果一致[32]。Chl-a是海洋浮游植物进行光合作用的主要色素[33], 因此浮游植物的数量可以决定Chl-a的含量, ENSO事件发生期间浮游生物群落在分布上会发生改变[34], 进而改变Chl-a的空间分布, 西北太平洋副热带环流区的Chl-a浓度在厄尔尼诺年偏高, 在拉尼娜年偏低[35]。本文研究结果显示不同气候事件会对浙江近海Chl-a的分布产生影响, 春季厄尔尼诺及拉尼娜发生期, 海域整体Chl-a浓度略高于正常年份;秋季厄尔尼诺发生期近岸Chl-a浓度明显高于正常年份及拉尼娜发生期, 拉尼娜发生期近海海域Chl-a浓度略高于正常年份。表明厄尔尼诺及拉尼娜事件均会导致浙江近海Chl-a浓度升高, 其中又以厄尔尼诺事件的影响更大, 与气候变化背景下东海Chl-a浓度上升的结论一致[36]。
本文结果表明, 不同气候事件的发生对海洋环境SST、Chl-a的分布影响较大, 相同气候事件对不同季节的海洋环境产生的影响也不尽相同, 进而导致鱼类生物量及时空分布都有所不同。
3.2 典型气候事件对浙江近海鱼类生物量的影响气候变化通过对海洋物理环境的影响来作用于其中的各种生物导致海洋物种对正在发生的气候变化作出了一系列的响应, 这些改变范围包括但不限于其分布范围、季节性活动、迁徙模式、丰度以及物种间的相互作用[1]。对鱼类的影响主要是由鱼类的产卵和索饵行为引起的, 而温度是影响鱼类行为最基本的因素之一[37], 盐度也是影响鱼类群落的重要环境因素, 不仅影响鱼类的生长、发育和繁殖, 而且影响种类和数量的时空分布[38]。浙江近海鱼类以暖水种为主、暖温种次之, SST是影响浙江近海鱼类生物量及时空变化的主要影响因子[6]。ENSO通过改变海表温度、盐度、叶绿素等海洋环境影响鱼类的生长、死亡率、繁殖力、产卵分布和移动, 对鱼类群落结构产生影响, 历史研究表明水温与渔获量之间存在显著的正相关[39—42], 而气候变化直接驱动SST的变化, 不同事件及相同事件不同强度对不同生物产生的影响不同, 已有研究表明中太平洋厄尔尼诺和东太平洋厄尔尼诺均会对鲣鱼资源丰度产生负面影响, 但前者的负面作用更加明显[43];相对于中强度厄尔尼诺事件(或中强度拉尼娜事件), 超强厄尔尼诺事件(或强拉尼娜事件)导致鲐鱼单位捕捞努力量渔获量骤减(或显著增加)[22];3—4月尼诺指数与次年东海南部渔场CPUE呈显著正相关[44];李菲萍等通过分析广西北部湾捕捞产量、厄尔尼诺的强度及持续时间等数据, 发现捕捞产量的增长率与厄尔尼诺强度、持续时间呈正相关关系[45], 管卫兵等对长江口日本鳗鲡苗的历史数据进行分析发现, 当以强厄尔尼诺事件为主时, SSTA越大, 鳗鲡捕捞量越低[46]。SST的变化会进一步影响Chl-a浓度的分布[47], 历史研究表明磷虾分布与Chl-a浓度呈正相关关系[48—49], 中心渔场的形成、变动与海域中的Chl-a浓度等关系紧密[50], 作为海洋中的初级生产力, Chl-a的改变会对浮游生物产生直接影响, ENSO期间浮游生物群落在分布上发生改变, 导致以浮游生物为饵料的鱼类发生相应的变化[51]。关于鱼类生物量变动与Chl-a浓度的关系历史研究较少。本文研究结果表明浙江近海生物量与SST及Chl-a均呈极显著正相关(P<0.01)。
拖网捕鱼对中低营养级的影响可能导致级联通过食物网向上或向下的作用, 进而对鱼类群落结构产生影响, 浙江省近海捕捞产量波动变化可分为三个阶段:第一阶段是2004—2008年, 捕捞产量波动下降阶段。第二个阶段是2008—2016年, 捕捞产量上升并稳定在较高水平上, 2016年之后, 捕捞产量大幅波动下降, 其中, 2018年最低[52]。而调查年间底层鱼类生物量在2013—2016年春秋季均呈现先降低后上升的趋势, 2016—2018年春季底层生物量呈先上升后下降趋势, 秋季则是呈现先下降后上升趋势, 与年际间的捕捞强度变化趋势并非一致, 可见捕捞强度的差异虽然会影响渔获量进而通过食物网对底层鱼类生物量产生一定影响, 但却不是影响底层鱼类生物量的决定性因素, 底层鱼类生物量还会受到气候变化的影响。对比不同气候事件期间春秋季生物量变化特征时发现:厄尔尼诺发生期间春秋季生物量均低于正常年份期间, 且以秋季最低;拉尼娜事件期间春秋季生物量均低于正常年份期间, 且以春季最低。T检验结果显示春季生物量在正常年份及拉尼娜事件发生期间具有显著性差异(P<0.05);秋季生物量在正常年份及厄尔尼诺事件发生期间具有显著性差异(P<0.01), 表明厄尔尼诺拉尼娜事件均对浙江近海鱼类生物量造成一定的影响。与陆鑫一等[53]拉尼娜年份(2007年)东海日本鲭的产量、CPUE均高于厄尔尼诺年份(2015年)的研究结果一致。
3.3 典型气候事件对浙江近海鱼类时空分布的影响气候变化会影响SST、Chl-a等海洋环境因子的分布, 当海洋环境因子变化超过鱼类耐受范围上限时, 鱼类活动水平和栖息地的分布就会发生改变, 进而影响鱼类生物量及在空间上的移动[54]。例如中国近海鲐鱼栖息地适宜性与厄尔尼诺和拉尼娜事件显著相关, 且随着异常气候事件强度的不同而发生变化:相对于中强度厄尔尼诺事件(或中强度拉尼娜事件), 超强厄尔尼诺事件(或强拉尼娜事件), 鲐鱼渔场适宜栖息地面积显著减小(或增大)[22];中太平洋厄尔尼诺和东太平洋厄尔尼诺对鲣鱼栖息地质量均产生负面影响[43]。随着北海变暖, 暖水性鱼类从东英吉利海峡向北迁移至北海, 而冷水性鱼类则是逐渐迁移至更深、更冷的地区[55]。在拉尼娜事件发生时, 东海鲐鱼渔场重心逐渐北移;而厄尔尼诺事件发生时, 渔场重心主要分布在南部海域[56]。陆鑫一等[53]发现拉尼娜年份日本鲭渔场纬度重心相对2015年(厄尔尼诺年份)偏北。厄尔尼诺期间, 印度洋鲣鱼围网作业渔场重心会向东、向北移动, 拉尼娜期间则相反[57—58]。除鱼类外, 厄尔尼诺和拉尼娜还会对柔鱼的产卵场产生影响:拉尼娜事件将使柔鱼产卵场的补充量减少, 且其渔场向北移动;而厄尔尼诺事件使产卵场的海洋环境更适于柔鱼生存。超强厄尔尼诺事件发生时, 柔鱼渔场环境不适于柔鱼生长, 适宜栖息地面积减少且向南移动, 导致该年份柔鱼资源丰度骤减, 渔场向南偏移[59]。
本文研究结果表明, 春季厄尔尼诺发生期鱼类重心偏北, 相较于正常年份期间, 厄尔尼诺及拉尼娜发生期间鱼类聚集性更强;秋季厄尔尼诺发生期鱼类重心向南移动, 拉尼娜期间则向北移动, 相较于正常年份及拉尼娜期间, 厄尔尼诺期间鱼类群落聚集性更强。不同典型气候事件在不同月份对海洋环境发生的影响也不尽相同。
鱼类的生物量及时空分布除受气候及环境影响外, 也受到其他外部因素影响。本文对浙江近海鱼类生物量及时空分布对典型气候事件的响应进行了初步研究, 后续将从更多方面切入, 例如加入不同气候事件的强度或多气候事件叠加, 更加全面系统地分析气候变化对浙江近海鱼类群落的影响。
| [1] |
Houghton J. Climate Change 2013-The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2014.
|
| [2] |
Hutchins B. Biodiversity of shallow reef fish assemblages in Western Australia using a rapid censusing technique. Records of the Western Australian Museum, 2001, 20: 247-270. |
| [3] |
Bjerknes J. Atmospheric teleconnections from the equatorial pacific1. Monthly Weather Review, 1969, 97(3): 163-172. DOI:10.1175/1520-0493(1969)097<0163:ATFTEP>2.3.CO;2 |
| [4] |
曹丛华, 曲如美, 张惠滋. 东中国海海温场对厄尔尼诺与反厄尔尼诺事件的响应. 海洋预报, 1999, 16(4): 39-45. |
| [5] |
张守文, 王辉, 姜华, 宋春阳, 杜凌. 厄尔尼诺和台风共同影响下的7月份黄、东海海温变化. 海洋学报, 2017, 39(12): 32-41. DOI:10.3969/j.issn.0253-4193.2017.12.004 |
| [6] |
陈美榕, 石少华, 沈红梅. 长江口海域海水表层温度与厄尔尼诺事件的关系. 海洋预报, 2005, 22(1): 80-85. DOI:10.3969/j.issn.1003-0239.2005.01.012 |
| [7] |
王平, 毛克彪, 孟飞, 袁紫晋. 中国东海海表温度时空演化分析. 国土资源遥感, 2020, 32(4): 227-235. |
| [8] |
何发祥, 陈清花, 郑爱榕. 厄尔尼诺与浙江近海冬汛带鱼渔获量关系. 海洋湖沼通报, 1995(3): 17-23. |
| [9] |
陈亚瞿, 朱启琴. 东海带鱼摄食习性、饵料基础及与渔场的关系. 水产学报, 1984, 8(2): 135-145. |
| [10] |
张秋华. 东海区渔业资源及其可持续利用. 上海: 复旦大学出版社, 2007.
|
| [11] |
唐嘉威, 于南京, 郑基, 许永久, 俞存根. 舟山渔场大小鱼山附近海域鱼类群落结构及生物多样性. 浙江海洋大学学报: 自然科学版, 2021, 40(3): 209-217, 233. DOI:10.3969/j.issn.1008-830X.2021.03.003 |
| [12] |
胡翠林, 张亚洲, 李德伟, 朱文斌, 蒋日进, 李鹏飞, 王洋, 周永东, 张洪亮. 浙江沿岸产卵场春、夏季鱼类资源密度和群落多样性研究. 水生生物学报, 2018, 42(5): 984-995. |
| [13] |
刘尊雷, 杨林林, 袁兴伟, 金艳, 严利平, 程家骅. 基于集成模型的小黄鱼越冬群体适宜生境及其环境影响因素. 应用生态学报, 2020, 31(6): 2076-2086. |
| [14] |
于南京, 俞存根, 许永久, 郑基, 刘坤, 张佩怡. 舟山群岛外海域春秋季鱼类群落结构及生物多样性. 水产学报, 2021, 45(8): 1374-1383. |
| [15] |
王迎宾, 郑基, 郑献之, 覃涛. 舟山渔场禁渔线以外海域单拖网鱼类群落结构变动分析. 南方水产科学, 2012, 8(1): 8-15. DOI:10.3969/j.issn.2095-0780.2012.01.002 |
| [16] |
张洪亮, 宋之琦, 潘国良, 陈峰, 周永东. 浙江南部近海春季鱼类多样性分析. 海洋与湖沼, 2013, 44(1): 126-134. |
| [17] |
洪华生, 何发祥, 杨圣云. 厄尔尼诺现象和浙江近海鲐鲹鱼渔获量变化关系──长江口ENSO渔场学问题之二. 海洋湖沼通报, 1997(4): 8-16. |
| [18] |
官文江, 陈新军, 李纲. 海表水温和拉尼娜事件对东海鲐鱼资源时空变动的影响. 上海海洋大学学报, 2011, 20(1): 102-107. |
| [19] |
李非, 陈新军, 朱清澄, 花传祥. 不同气候模态下西北太平洋秋刀鱼海况特征分析. 南方水产科学, 2018, 14(3): 20-28. DOI:10.3969/j.issn.2095-0780.2018.03.003 |
| [20] |
刘祝楠, 陈新军. 不同气候模态下西北太平洋秋刀鱼资源丰度预测模型建立. 海洋学报, 2018, 40(6): 74-82. |
| [21] |
代丹娜, 刘洪生, 戴小杰, 田思泉. ENSO现象与东太平洋黄鳍金枪鱼围网CPUE时空分布的关系. 上海海洋大学学报, 2011, 20(4): 571-578. |
| [22] |
郭爱, 张扬, 余为, 陈新军, 钱卫国, 李曰嵩. 两类强度厄尔尼诺和拉尼娜事件对中国近海鲐鱼栖息地的影响. 海洋学报, 2018, 40(12): 58-67. |
| [23] |
气候变化对鲐鱼栖息地的迁徙影响. 2014年中国水产学会学术年会论文摘要集. 长沙, 2014: 382.
|
| [24] |
Su L, Miao C Y, Duan Q Y, Lei X H, Li H. Multiple-wavelet coherence of world's large rivers with meteorological factors and ocean signals. Journal of Geophysical Research: Atmospheres, 2019, 124(9): 4932-4954. DOI:10.1029/2018JD029842 |
| [25] |
张强, 孙鹏, 王野乔. 鄱阳湖流域气候变化及水文响应研究. 北京: 中国水利水电出版社, 2015.
|
| [26] |
杨若文, 曹杰, 黄玮, 年艾冰. 太阳常数与太阳黑子数关系的交叉小波分析. 科学通报, 2009, 54(7): 871-875. |
| [27] |
余丹丹, 张韧, 洪梅, 刘科峰, 王辉赞, 桂祁军. 赤道中太平洋对流活动与西太平洋副高西伸的时延相关分析. 海洋科学进展, 2008, 26(3): 292-304. |
| [28] |
莫崇勋, 蓝华鲲, 杨莹, 阮俞理, 孙桂凯, 黄亚. 广西典型岩溶区流域降雨侵蚀力变化特征及影响因素研究. 广西大学学报: 自然科学版, 2021, 46(5): 1204-1213. |
| [29] |
肖启华, 黄硕琳. 气候变化对海洋渔业资源的影响. 水产学报, 2016, 40(7): 1089-1098. |
| [30] |
D'Antoni E. Climate change implications for fisheries and aquaculture Overview of current scientific knowledge, 2009.
|
| [31] |
戴鑫烽, 曾宇兰, 李冬融, 王鹏斌, 郭若玉, 王瑞芳, 陆斗定. 长江口甲藻孢囊与水体富营养化和厄尔尼诺指数关系研究. 海洋学报, 2021, 43(2): 78-87. |
| [32] |
胡明娜, 赵朝方. 浙江近海夏季上升流的遥感观测与分析. 遥感学报, 2008, 12(2): 297-304. |
| [33] |
林丽贞, 陈纪新, 刘媛, 曹振锐, 黄邦钦. 东、黄海典型海区分粒级浮游植物叶绿素a的周日波动及影响因子. 台湾海峡, 2007, 26(3): 342-350. |
| [34] |
李淼, 许友伟, 孙铭帅, 张俊, 李佳俊, 陈作志, 张魁. 气候变化对海洋鱼类群落结构的影响研究进展. 海洋科学, 2022, 46(7): 120-129. |
| [35] |
陈小燕. 基于遥感的长时间序列浮游植物的多尺度变化研究[D]. 杭州: 浙江大学, 2013.
|
| [36] |
张俊鹏, 蔡榕硕. 东海冷涡对东亚季风年代际变化响应的数值试验. 海洋与湖沼, 2013, 44(6): 1427-1435. |
| [37] |
García Molinos J, Halpern B S, Schoeman D S, Brown C J, Kiessling W, Moore P J, Pandolfi J M, Poloczanska E S, Richardson A J, Burrows M T. Climate velocity and the future global redistribution of marine biodiversity. Nature Climate Change, 2016, 6(1): 83-88. |
| [38] |
陈伟峰, 彭欣, 汪振华, 谢起浪, 陈少波, 叶深, 陈骁, 艾为明. 浙南近海秋冬季鱼类群落结构研究. 海洋开发与管理, 2017, 34(11): 111-119. |
| [39] |
Zhang C I, Lee J B, Kim S, Oh J H. Climatic regime shifts and their impacts on marine ecosystem and fisheries resources in Korean waters. Progress in Oceanography, 2000, 47(2/3/4): 171-190. |
| [40] |
何发祥, 洪华生, 陈刚. ENSO现象与台湾海峡西部海区中下层鱼类渔获量关系. 海洋湖沼通报, 2003(1): 27-34. |
| [41] |
王继隆, 李继龙, 杨文波, 王国伟. 主要气侯因子对东中国海主要经济鱼种生物量的影响研究. 湖南农业科学, 2010(9): 142-147. |
| [42] |
Tian Y J, Kidokoro H, Watanabe T, Iguchi N. The late 1980s regime shift in the ecosystem of Tsushima warm current in the Japan/East Sea: evidence from historical data and possible mechanisms. Progress in Oceanography, 2008, 77(2/3): 127-145. |
| [43] |
Yen K W, Wang G H, Lu H J. Evaluating habitat suitability and relative abundance of skipjack (Katsuwonus pelamis) in the Western and Central Pacific during various El Niño events. Ocean & Coastal Management, 2017, 139: 153-160. |
| [44] |
崔科, 陈新军. 东黄海鲐鱼资源丰度与表温关系. 南方水产, 2007, 3(4): 20-25. |
| [45] |
李菲萍, 吴志强, 钟志坚, 黄德练, 姜恒. 厄尔尼诺现象对广西海洋捕捞产量的影响. 海洋湖沼通报, 2011(3): 62-68. |
| [46] |
管卫兵, 丁华腾, 戴小杰, 陈新军, 宣富君. ENSO对长江口日本鳗鲡苗捕捞量的影响. 台湾海峡, 2009, 28(1): 45-51. |
| [47] |
江杉, 王益澄, 马仁锋. 中国东海叶绿素浓度变化分析及其海水温度响应. 测绘通报, 2020(6): 39-44. |
| [48] |
Marrari M, Hu C M, Daly K. Validation of SeaWiFS chlorophyll a concentrations in the Southern Ocean: a revisit. Remote Sensing of Environment, 2006, 105(4): 367-375. |
| [49] |
Meguro H, Toba Y, Murakami H, Kimura N. Simultaneous remote sensing of chlorophyll, sea ice and sea surface temperature in the Antarctic waters with special reference to the primary production from ice algae. Advances in Space Research, 2004, 33(7): 1168-1172. |
| [50] |
沈新强, 王云龙, 袁骐, 黄洪亮, 周爱忠. 北太平洋鱿鱼渔场叶绿素a分布特点及其与渔场的关系. 海洋学报, 2004, 26(6): 118-123. |
| [51] |
Hwang K, Jung S. Decadal changes in fish assemblages in waters near the Ieodo Ocean research station (East China Sea) in relation to climate change from 1984 to 2010. Ocean Science Journal, 2012, 47(2): 83-94. |
| [52] |
辛艺. 浙江省近海捕捞作业结构现状及调整策略研究[D]. 舟山: 浙江海洋大学, 2020.
|
| [53] |
陆鑫一, 刘子历, 余为, 陈新军, 李纲. 拉尼娜(2007年)和厄尔尼诺(2015年)事件对东海日本鲭栖息地时空分布的影响. 海洋渔业, 2021, 43(2): 137-148. |
| [54] |
Free C M, Thorson J, Pinsky M, Oken K L, Wiedenmann J, Jensen O. Impacts of historical warming on marine fisheries production. Science, 2019, 363: 979-983. |
| [55] |
McLean M, Mouillot D, Lindegren M, Engelhard G, Villéger S, Marchal P, Brind'Amour A, Auber A. A climate-driven functional inversion of connected marine ecosystems. Current Biology, 2018, 28(22): 3654-3660. |
| [56] |
李宜锴, 方星楠, 余为, 李曰嵩. 2005—2016年中国东海鲐鱼渔场的时空分布及与海表面温度的关联. 上海海洋大学学报, 2022, 31(3): 710-720. |
| [57] |
李亚楠, 陈新军. 印度洋鲣鱼围网资源渔场时空变化及其与ENSO的关系. 海洋学报, 2017, 39(4): 72-78. |
| [58] |
周甦芳. 厄尔尼诺-南方涛动现象对中西太平洋鲣鱼围网渔场的影响. 中国水产科学, 2005, 12(6): 739-744. |
| [59] |
Chen X J, Zhao X H, Chen Y. Influence of El Niño/La Niña on the western winter-spring cohort of neon flying squid (Ommastrephes bartramii) in the northwestern Pacific Ocean. ICES Journal of Marine Science, 2007, 64(6): 1152-1160. |