 2024, Vol. 44
2024, Vol. 44文章信息
- 叶曦, 方笛熙, 张锋
- YE Xi, FANG Dixi, ZHANG Feng
- 高阶作用对植物-传粉者-植食动物集合群落续存的影响
- Effects of Higher-Order Interactions on the persistence of plant-pollinator-herbivore metacommunities
- 生态学报. 2024, 44(1): 246-255
- Acta Ecologica Sinica. 2024, 44(1): 246-255
- http://dx.doi.org/10.20103/j.stxb.202211173321
-
文章历史
- 收稿日期: 2022-11-17
- 网络出版日期: 2023-09-28

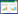




2. 云南财经大学统计与数学学院, 昆明 650221
2. Statistics and Mathematics School, Yunnan University of Finance and Economics, Kunming 650221, China
种间关系是物种共存和生物多样性维持的重要驱动力[1]。传统的种间关系研究聚焦成对物种间的直接作用[2], 然而越来越多的实验和理论研究开始关注更为复杂的间接作用[2—3]。其中, 高阶作用(Higher-Order Interactions, HOIs)在调节多物种系统动态方面的作用尤其引人关注[4—5]。高阶作用的定义和相关术语的使用目前还没有一个统一的标准[2]。最初, 高阶作用和间接作用的定义较为混淆, 都被广义地定义为一个物种对另外两个物种之间的相互作用所施加的影响[6—7]。而后Wootton[8]强调间接作用需要有中间物种的存在, 将间接作用进一步细分为两类:一类为“相互作用链(interaction chains)”, 即一个物种对中间物种密度的改变最终影响到了第三个目标物种; 另一类为“相互作用的调节(interaction modification)”[9], 即一个物种的存在对另外两个物种之间的相互作用强度产生的影响, 具体来说是一个物种对中间物种可塑性性状的改变产生影响, 最终影响到了第三个目标物种。这是传统意义上高阶作用的定义, 也被称为性状介导的间接作用和狭义高阶作用(Hard-HOIs)[2, 10—11], 这一定义描述的是发生高阶作用的具体生态过程, 将高阶作用适用范围划分为涉及至少三个物种的相互作用[9, 12]。近年来生态学家将高阶作用的定义扩展为“种群密度对每员增长率的非加性效应(non-additive effects)”[13—15], 该定义被称为广义高阶作用(Soft-HOIs)[2, 11]。这里的加性效应指的是一个物种受到一个或几个物种的影响可以表示为这些物种影响的累加总和[16], 反之则是非加性效应。由于相互作用的调节的模型一定是非加性的[4], 但非加性并不意味着只有相互作用的调节这一情况[16], 因此定义“非加性效应”除了包括定义“相互作用的调节”以外, 还包括其他的非加性情况, 如两个物种间存在的高阶作用[5]。Abrams[11]根据对中间物种的调节方式将间接作用分为密度调节(相互作用链)和性状调节(高阶作用)两类。然而, 也有生态学家认为高阶作用和间接作用属于两种不同的互作类型。Billick & Case[4]提出成对关系可以预测间接作用但不可以预测高阶作用、两者在模型中表示方法不同和所发生的时间尺度不同等理由, 以此说明高阶作用不属于间接作用。
高阶作用在生态系统中的作用往往被低估和轻视[17], 但理论研究和实验研究都有证据表明它的作用是显著的[18—19]。高阶作用能够促进竞争物种的长期稳定共存[20], 在维持生态系统多样性、群落结构和调节生态系统功能方面有重要影响[3]。植物、传粉者和植食动物三者之间存在高阶作用[21—23], 植食动物影响植物和传粉者之间的互惠关系, 既包括对传粉者对植物传粉效果的影响, 也包括植物对传粉者传粉回报的影响。已有研究表明植食动物会削弱传粉者对植物的传粉作用, 如植食动物通过损害或移除花朵(减少了花朵的数量、形态结构、花粉和花蜜产量等), 减少植物对传粉者的有利资源, 降低传粉者对植物的传粉频率[24—26]; 但也有研究表明植食动物可诱导植物产生次生代谢物作为传粉者的抗生素来增强传粉作用, 如植食动物对芥菜Sinapis arvensis根部的食草性增加了传粉者对芥菜花的访问[27]。然而, 过往研究很少将植物、传粉者和植食动物三者间的复杂相互作用同时考虑在一起[28—29], 特别是这三者间存在的高阶作用不可忽视。
集合种群是指在一定地域内, 由若干相邻的、半隔离的, 通过个体迁移而联系起来的局域种群组成的集合体, 局域种群随时会发生随机性灭绝, 但集合种群是较为稳定而持久的存在[30]。集合群落的概念是由集合种群的概念延伸而来, 它指定了一组空间分布不同但通过扩散连接而成的群落[31]。集合种群的研究对象大致可分为两类, 一类是物种生境本身即为斑块化的情况, 其具有较明显的离散化特性; 另一类是因生境丧失和生境破碎化导致该物种的生境由连续变为斑块镶嵌结构的情况[31—32]。传粉者和植食动物的生境即为分布在不同斑块上的植物, 三者的局域种群也面临着随机灭绝的风险, 因此集合种群尺度适用于研究这三者之间的高阶作用。
在已有研究的基础上[33], 建立了植物-传粉者-植食动物集合群落模型, 考虑了由植食动物改变植物和传粉者之间互惠关系而形成的“相互作用的调节”(高阶作用), 同时也考虑了植食动物引起的植物灭绝而导致传粉者灭绝而形成的“相互作用链”(间接作用), 着重研究高阶作用对植物-传粉者-植食动物集合群落平衡态和续存的影响。
1 模型与方法考虑由植物、传粉者和植食动物构成的集合群落, 每个斑块处于五种可能状态中一种, 或只有植物, 或植物和传粉者共同占有, 或植物和植食动物共同占有, 或植物、传粉者和植食动物共同占有, 亦或是空斑块。传粉动物和植食动物完全依赖于植物提供的资源生存和繁殖, 所以在无植物的斑块不能生存。植物不完全依赖该传粉动物生存, 可以通过其它方式输出繁殖体(比如无性繁殖)。传粉动物的出现可以通过给植物传粉增强植物的繁殖输出。植食动物的出现由于植食作用对植物造成负面影响, 使植物灭绝率增加。假设在三者都有的斑块上会产生高阶作用, 即植食动物影响植物和传粉者的互惠关系, 既包括对传粉效果的影响, 也包括对传粉者回报的影响。同时空斑块可能被植物的繁殖体重新侵占而变成只被植物占有的斑块, 只有植物的斑块由于传粉者或植食动物的迁入而变成动植物共同占有的斑块, 传粉者或植食动物与植物共同占有的斑块可能被另一种动物迁入而变成三物种共同占有的斑块。斑块上的动植物局域种群都存在灭绝风险, 特别需要注意的是, 由于动物对植物的依赖性, 斑块中植物的灭绝使伴随其上的动物也灭绝。
如果用p1, p12, p13, p123, p0分别表示只有植物占有的斑块, 有植物和传粉者共同占有的斑块, 有植物和植食动物共同占有的斑块, 有植物、传粉者和植食动物三者共同占有的斑块, 空斑块这五种斑块状态所占比例, 显然有p1+p12+p13+p123+p0=1, 可见这五个变量中只有四个独立, 这里我们选取变量p1, p12, p13, p123描述它们随时间的动态变化:
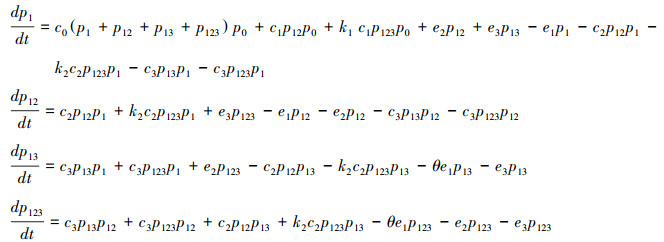
|
(1) |
式中, c0是植物独立于传粉者的固有侵占率, c1是由传粉造成的侵占率, c2和c3分别是传粉者和植食动物的侵占率, e1、e2和e3分别是植物、传粉者和植食动物局域种群的灭绝率, 这些参数都大于0。另外, 参数θ(>)表示了植食动物对植物局域种群灭绝率的影响; 参数k1(>0)表示植食动物对传粉效果的影响, k2(>0)表示植食动物对传粉者回报的影响, 大于1表示正高阶作用(positive HOIs), 小于1表示负高阶作用(negative HOIs), 它们都是植食动物对传粉关系的影响, 即高阶作用。
P1=p1+p12+p13+p123表示植物占有的斑块比例, P2=p12+p123表示传粉者占有的斑块比例, P3=p13+p123表示植食动物占有的斑块比例。分别求它们关于时间的导数, 可以将方程(1)等价地写成关于变量P1、P2和P3的微分方程:
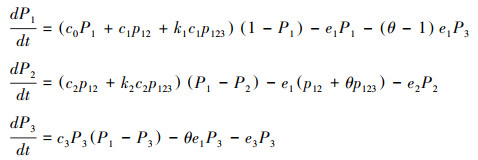
|
(2) |
方程(2)由于包含了额外变量p12和p123, 并不是封闭系统。根据概率原理, p123=p23q1|23, 其中p23表示斑块中同时有传粉者和植食动物的概率, q1|23表示同时有传粉者和植食动物的斑块中也有植物的条件概率。根据模型假设, 传粉者和植食动物依赖于植物, 所以有传粉者和植食动物的斑块中必然有植物, 即q1|23=1。如果进一步假设传粉者和植食动物对斑块的占有相互独立, 即p23=P2P3, 那么p123=P2P3。另外知p12=P2-p123, 由此方程(2)可以变成如下的封闭系统:
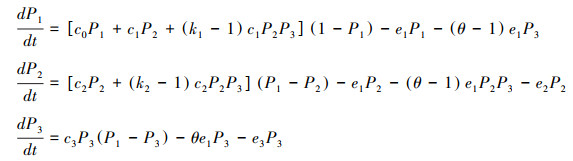
|
(3) |
方程(3)中出现的三个高次项, 其中(k1-1)c1P2P3描述了植食动物影响传粉效果的高阶作用, (k2-1)c2P2P3描述了植食动物影响传粉回报的高阶作用, (θ-1)e1P2P3表示了由于植食动物引起的植物灭绝而导致传粉者灭绝的间接作用。此外, (θ-1)e1P3表示植食动物引起植物的灭绝, θe1P3表示植食动物引起植物的灭绝而导致自身灭绝的反噬作用。特别当k1、k2或θ等于1时, 表示这些高阶作用或间接作用消失。
模型抓住植食动物在局域尺度上影响动植物传粉关系而形成的高阶作用, 以及它们对植物的伤害而造成的对传粉者的间接作用, 描述这三个物种在集合群落水平上的种群动态过程。所以, 可用于研究这些高阶作用和间接作用对集合群落平衡态和续存的影响。首先, 分析这三个物种集合群落在无高阶作用和间接作用时(k1=k2=θ=1)的动态行为, 再探索高阶作用和间接作用如何影响群落的平衡态和续存。
2 分析和结果 2.1 无高阶作用和间接作用的情况在无高阶作用和间接作用时(k1=k2=θ=1), 给出该系统所有非平凡平衡点的平衡条件, 并且借助软件Matlab R2016a, 基于方程(3)使用牛顿迭代法对其进行数值模拟, 展示这些平衡点所属的四种集合群落的续存情况:
(a) 当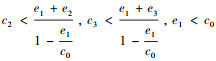

|
| 图 1 无高阶作用和间接作用时集合种群相空间图 Fig. 1 Phase space diagram of the pooled population in the absence of Higher-Order Interactions and indirect interactions |
(1) 实心圆点表示稳定的平衡点, 空心圆点表示不稳定的平衡点; (2)A为情况(a), 参数设置:c0=2, c1=c2=c3=1, e1=1, e2=0.4, e3=0.1;B为情况(b), 参数设置:c0=2, c1=1, c2=2, c3=8, e1=1, e2=0.4, e3=0.1; C和D为情况(c), 参数设置:c0=2, c1=8, c2=8, c3=1, e1=1.2, e2=0.4, e3=0.1和c0=2, c1=8, c2=8, c3=1, e1=1.8, e2=0.4, e3=0.1;E和F为情况(d), 参数设置:c0=2, c1=8, c2=8, c3=20, e1=1.2, e2=0.4, e3=0.1和c0=2, c1=8, c2=8, c3=20, e1=1.7, e2=0.4, e3=0.1
(b) 当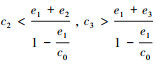
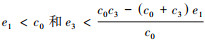
(c) 当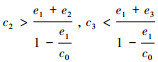
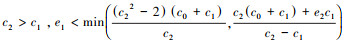
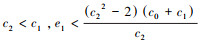
(d) 当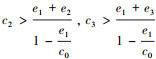
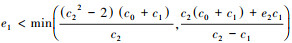
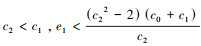

|
| 图 2 无高阶作用和间接作用时集合种群占有率变化情况 Fig. 2 Dynamics of metapopulation occupancy in the absence of Higher-Order Interactions and indirect interactions (1)实线表示达到稳定时集合种群占有率变化曲线, 虚线表示不稳定时集合种群占有率变化曲线; (2)参数设置:c0=2, c1=8, c2=8, c3=20, e2=0.4, e3=0.1 |
分别调查影响传粉效果的高阶作用项k1(图 3A、B和C)、影响传粉回报的高阶作用项k2(图 3 D、E和F)和间接作用参数θ(图 3 G、H和I)对集合群落动态的影响。基于系统(3)的分岔图表明, 高阶作用和间接作用都会显著影响互惠关系(正反馈)引起的双稳定性。与无高阶作用的情况(图 3中绿线)相比, 两种正高阶作用都扩大集合种群出现双稳态现象的参数范围(植物灭绝率的取值范围; 图 3A-F中红线); 负高阶作用减小集合种群出现双稳态现象的参数范围(图 3A-F中蓝线), 但未使得双稳态现象消失。高阶作用也显著地影响集合群落的续存, 正高阶作用降低集合群落的灭绝阈值, 有利于集合群落续存, 并且使得集合群落达到动态平衡时占有率增加(图 3A—F中红线); 负高阶作用抬高该灭绝阈值, 不利于集合群落续存, 集合群落达到动态平衡时占有率也降低(图 3A—F中蓝线)。间接作用的增强会缓解双稳态现象, 不仅灭绝阈值抬高, 而且双稳态范围也会缩小(图 3G—I)。特别地, 随着间接作用的增强, 集合种群出现第一种双稳态现象(图 2蓝色区域)的植物灭绝率更低(图 3G—I), 这是间接作用和高阶作用的不同之处, 但间接作用也并未使得双稳态现象消失。

|
| 图 3 高阶作用k1、k2和间接作用θ分别对集合种群占有率变化情况 Fig. 3 Higher-Order Interactions k1, k2 and indirect interactions θ on metapopulation occupancy changes respectively (1)实线表示集合群落达到动态平衡时占有率变化曲线, 虚线表示集合群落未达到动态平衡时占有率变化曲线; (2)参数设置:图A、B和C中k1变化, k2=θ=1;图D、E和F中k2变化, k1=θ=1;图G、H和I中θ变化, k1=k2=1。 |
在简化系统(1)为系统(3)的过程中, 假设传粉者和植食动物对斑块的占有相互独立(p123=P2P3), 为了探究这种独立性假设所造成的影响, 我们对系统(1)和系统(3)进行对比(图 4)。相较于系统(3), 未假设独立性时正高阶作用(负高阶作用)能够扩大(减小)集合种群出现双稳态现象的参数范围, 抬高(降低)集合种群的灭绝阈值(图 4D—F)。此外, 进一步对比了系统(1)中的p123和系统(3)中的P2P3在各高阶作用项和间接作用项下的差异性(图 5)。所有的模拟结果均显示p123>P2P3, 这表明假设(p123=P2P3)低估斑块中同时存在传粉者和植食动物的比率, 说明系统(1)中物种不是随机分布的, 传粉者和植食动物更倾向于占有同一斑块。但由数值解的结果可知, 尽管这种独立性假设在量上改变集合群落占有率, 但并非质的影响(集合群落中双稳态现象并没有消失), 因此这种假设合理, 可以达到简化模型的目的。

|
| 图 4 高阶作用下集合种群占有率变化情况 Fig. 4 Changes in metapopulation occupancy by Higher-Order Interactions (1)实线表示集合群落达到动态平衡时占有率变化曲线, 虚线表示集合群落达到动态平衡时占有率变化曲线; (2)图A、B和C为假设独立性的情况, 图D、E和F为未假设独立性的情况。所有图中θ=1, 在没有高阶作用和间接作用时(图中绿线)集合种群占有率变化曲线重合 |

|
| 图 5 独立性假设对集合种群占有率的影响 Fig. 5 Effect of hypothesized independence on metapopulation occupancy (1)蓝线表示未假设独立性的情况, 绿线表示假设独立性的情况; (2)参数设置:图A中k1变化, k2=θ=1;图B中k2变化, k1=θ=1;图C中θ变化, k1=k2=1 |
通过建立植物-传粉者-植食动物集合群落模型, 研究高阶作用对该集合群落续存的影响。经过模拟, 随着植物灭绝率增加, 集合群落的稳定状态从单稳态转变为双稳态。这种双稳态现象在有互惠关系(正反馈)的群落中常有出现。正反馈可能会引发自然生态系统在不同稳定状态之间交替, 如水生植物生长衰退导致浅水湖由清澈态转变为浑浊态, 生长旺盛则导致浅水湖由浑浊态转变为清澈态[34—36]。研究结果表明, 正高阶作用(负高阶作用)扩大(缩小)集合群落出现双稳态现象的参数范围, 本质上是扩大(缩小)物种共存范围, 但高阶作用未使得双稳态现象消失和物种灭绝, 这说明高阶作用会改变物种间相互作用强度, 但不会打破多物种原本的共存状态[33]; 正高阶作用降低集合群落的灭绝阈值, 增加集合群落稳定时的占有率, 对生物多样性的维持起促进作用, 有利于集合群落的续存, 负高阶作用则相反。野外研究表明, 分布于北美洲的红凯木(Alnus rubra)在被植食昆虫采食后会产生化学防御反应, 通过降低叶片氮含量来提高叶片碳氮比(植食昆虫和水生分解者均偏好低碳氮比的叶片), 在维持植食昆虫较低丰度的同时并未使得水生分解者灭绝[2, 37]; 加拿大蓟(Cirsium arve) 释放花香会同时引来传粉者与植食动物, 植食动物通过破坏加拿大蓟的花朵来影响传粉者的适应性及丰度[22]。这些结果与本文研究结果一致, 说明高阶作用对调节多物种系统动态具有重要作用。
研究结果表明间接作用会出现高次项, 系统(3)中(k1-1)c1P2P3, (k2-1)c2P2P3和(θ-1)e1P2P3均有高次项, 其中(k1-1)c1P2P3和(k2-1)c2P2P3描述由植食动物影响传粉效果和回报的高阶作用, (θ-1)e1P2P3描述由于植食动物引起的植物灭绝而导致传粉者灭绝的间接作用。出现这种由间接作用产生的高次项, 可能是因为中两种相互关系的生态过程实际上发生在斑块内部, 而对斑块占有率所进行的建模研究建立在集合种群尺度上, 对斑块之间的关系进行模型描述, 其中变量并未表示斑块内部的属性。不同生态尺度上间接作用的表现形式可能并不相同, 在种群尺度上定义的间接作用是改变物种密度来传递的效应, 系统不会出现高次项; 但在集合种群尺度上对斑块的占有率进行建模, 系统可能会出现高次项。
基于集合种群理论, 从斑块类型相互转化入手推导出模型, 直观地展示模型中高次项的形成机制, 相较于直接给出高次项的做法更具说服力。假设传粉者和植食动物侵占斑块的独立性, 模拟结果显示这种假设只会影响集合种群占有率的大小, 不会产生本质影响, 因此这种独立性假设合理, 模型得到有效简化。此外, 在植物和传粉者是互惠关系的前提下进行研究, 但自然界同样存在植物和传粉者之间非互惠关系或两种关系都存在的情况, 如榕树-榕小蜂系统中的榕小蜂分为传粉榕小蜂和非传粉榕小蜂(欺骗者)[38], 本研究的模型依然适用于此种情况, 仅需将数值模拟时将传粉者帮助增强植物繁殖输出的参数c1的值取0, 但此类情况在自然界中是否还存在高阶作用, 有待进一步研究。
| [1] |
Levine J M, Bascompte J, Adler P B, Allesina S. Beyond pairwise mechanisms of species coexistence in complex communities. Nature, 2017, 546(7656): 56-64. DOI:10.1038/nature22898 |
| [2] |
李远智, 肖俊丽, 刘翰伦, 王酉石, 储诚进. 生物间高阶相互作用研究进展. 生物多样性, 2020, 28(11): 1333-1344. DOI:10.17520/biods.2020217 |
| [3] |
朱玉, 王德利, 钟志伟. 生态系统基于性状调节的物种间接作用: 特征、成因及后果. 生态学报, 2017, 37(23): 7781-7790. DOI:10.5846/stxb201610011986 |
| [4] |
Billick I, Case T J. Higher order interactions in ecological communities: what are they and how can they be detected?. Ecology, 1994, 75(6): 1529-1543. DOI:10.2307/1939614 |
| [5] |
Case T J, Bender E A. Testing for higher order interactions. The American Naturalist, 1981, 118(6): 920-929. DOI:10.1086/283885 |
| [6] |
Strauss S Y. Indirect effects in community ecology: their definition, study and importance. Trends in Ecology & Evolution, 1991, 6(7): 206-210. |
| [7] |
Worthen W B, Moore J L. Higher-order interactions and indirect effects: a resolution using laboratory Drosophila communities. The American Naturalist, 1991, 138(5): 1092-1104. DOI:10.1086/285271 |
| [8] |
Wootton J T. The nature and consequences of indirect effects in ecological communities. Annual Review of Ecology and Systematics, 1994, 25: 443-466. DOI:10.1146/annurev.es.25.110194.002303 |
| [9] |
Wootton J T. Indirect effects and habitat use in an intertidal community: interaction chains and interaction modifications. The American Naturalist, 1993, 141(1): 71-89. DOI:10.1086/285461 |
| [10] |
Kleinhesselink A, Kraft N J B, Levine J. Mechanisms underlying higher order interactions: from quantitative definitions to ecological processes. BioRxiv, 2019. |
| [11] |
Abrams P A. Implications of dynamically variable traits for identifying, classifying, and measuring direct and indirect effects in ecological communities. The American Naturalist, 1995, 146(1): 112-134. DOI:10.1086/285789 |
| [12] |
Werner E E. Individual behavior and higher-order species interactions. The American Naturalist, 1992, 140: S5-S32. DOI:10.1086/285395 |
| [13] |
Mayfield M M, Stouffer D B. Higher-order interactions capture unexplained complexity in diverse communities. Nature Ecology & Evolution, 2017, 1(3): 1-7. |
| [14] |
Mickalide H, Kuehn S. Higher-order interaction between species inhibits bacterial invasion of a phototroph-predator microbial community. Cell Systems, 2019, 9(6): 521-533. DOI:10.1016/j.cels.2019.11.004 |
| [15] |
Vandermeer J H. The competitive structure of communities: an experimental approach with Protozoa. Ecology, 1969, 50(3): 362-371. DOI:10.2307/1933884 |
| [16] |
Adler F R, Morris W F. A general test for interaction modification. Ecology, 1994, 75(6): 1552-1559. DOI:10.2307/1939616 |
| [17] |
Chesson P. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 2000, 31: 343-366. DOI:10.1146/annurev.ecolsys.31.1.343 |
| [18] |
Abrams P A. Arguments in favor of higher order interactions. The American Naturalist, 1983, 121(6): 887-891. DOI:10.1086/284111 |
| [19] |
Barabás G, Michalska-Smith M J, Allesina S. The effect of intra- and interspecific competition on coexistence in multispecies communities. The American Naturalist, 2016, 188(1): E1-E12. DOI:10.1086/686901 |
| [20] |
Grilli J, Barabás G, Michalska-Smith M J, Allesina S. Higher-order interactions stabilize dynamics in competitive network models. Nature, 2017, 548(7666): 210-213. DOI:10.1038/nature23273 |
| [21] |
Krupnick G A, Weis A E, Campbell D R. The consequences of floral herbivory for pollinator service to Isomeris arborea. Ecology, 1999, 80(1): 125. DOI:10.1890/0012-9658(1999)080[0125:TCOFHF]2.0.CO;2 |
| [22] |
Theis N. Fragrance of Canada thistle (Cirsium arvense) attracts both floral herbivores and pollinators. Journal of Chemical Ecology, 2006, 32(5): 917-927. DOI:10.1007/s10886-006-9051-x |
| [23] |
Danderson C A, Molano-Flores B. Effects of herbivory and inflorescence size on insect visitation to Eryngium yuccifolium (Apiaceae) a prairie plant. The American Midland Naturalist, 2010, 163(1): 234-246. DOI:10.1674/0003-0031-163.1.234 |
| [24] |
Strauss S Y. Floral characters link herbivores, pollinators, and plant fitness. Ecology, 1997, 78(6): 1640-1645. DOI:10.1890/0012-9658(1997)078[1640:FCLHPA]2.0.CO;2 |
| [25] |
Adler L S, Wink M, Distl M, Lentz A J. Leaf herbivory and nutrients increase nectar alkaloids. Ecology Letters, 2006, 9(8): 960-967. DOI:10.1111/j.1461-0248.2006.00944.x |
| [26] |
Kessler A, Halitschke R. Testing the potential for conflicting selection on floral chemical traits by pollinators and herbivores: predictions and case study. Functional Ecology, 2009, 901-912. |
| [27] |
Richardson D M, Bond W J. Determinants of plant distribution: evidence from pine invasions. The American Naturalist, 1991, 137(5): 639-668. DOI:10.1086/285186 |
| [28] |
Sauve A M C, Fontaine C, Thébault E. Structure-stability relationships in networks combining mutualistic and antagonistic interactions. Oikos, 2014, 123(3): 378-384. DOI:10.1111/j.1600-0706.2013.00743.x |
| [29] |
Fontaine C, Guimarães P R Jr, Kéfi S, Loeuille N, Memmott J, van der Putten W H, van Veen F J F, Thébault E. The ecological and evolutionary implications of merging different types of networks. Ecology Letters, 2011, 14(11): 1170-1181. DOI:10.1111/j.1461-0248.2011.01688.x |
| [30] |
张大勇, 雷光春, ILKKA HANKI. 集合种群动态: 理论与应用. 生物多样性, 1999, 7(2): 81-90. |
| [31] |
张彦宇. 集合种群时空动态对生态位构建的响应研究[D]. 兰州: 兰州大学, 2008.
|
| [32] |
储诚进, 王酉石, 刘宇, 蒋林, 何芳良. 物种共存理论研究进展. 生物多样性, 2017, 25(4): 345-354. |
| [33] |
Zhang H L, Chen D D, Ying Z X, Zhang F, Liao J B. Robustness of the pollination-herbivory system with high-order interactions to habitat loss. Ecological Modelling, 2019, 414: 108826. DOI:10.1016/j.ecolmodel.2019.108826 |
| [34] |
Zhang F, Tao Y, Hui C. Organism-induced habitat restoration leads to bi-stability in metapopulations. Mathematical Biosciences, 2012, 240(2): 260-266. DOI:10.1016/j.mbs.2012.08.006 |
| [35] |
Zhang D, Song W X, Chen N L, Zhang F. The role of spatial scale in organism-environment positive feedback. Nonlinear Dynamics, 2019, 95(3): 2019-2029. DOI:10.1007/s11071-018-4674-3 |
| [36] |
李玉照, 刘永, 赵磊, 邹锐, 王翠榆, 郭怀成. 浅水湖泊生态系统稳态转换的阈值判定方法. 生态学报, 2013, 33(11): 3280-3290. DOI:10.5846/stxb201203140340 |
| [37] |
Jackrel S L, Wootton J T. Cascading effects of induced terrestrial plant defences on aquatic and terrestrial ecosystem function. Proceedings Biological Sciences, 2015, 282(1805): 20142522. |
| [38] |
徐睿, 张媛, 彭艳琼, 杨大荣. 高榕雌花期传粉榕小蜂和欺骗性小蜂的繁殖特点. 生态学报, 2016, 36(04): 1134-1140. DOI:10.5846/stxb201406251310 |