 2024, Vol. 44
2024, Vol. 44文章信息
- 赵国清, 信忠保, 柳金昊, 刘亚玲, 黄艳章, 李宗善
- ZHAO Guoqing, XIN Zhongbao, LIU Jinhao, LIU Yaling, HUANG Yanzhang, LI Zongshan
- 澜沧江源区高山柏径向生长对气候变化的响应
- Response of radial growth of Sabina squamata to climatic change in the Source Region of Lancang River
- 生态学报. 2024, 44(1): 235-245
- Acta Ecologica Sinica. 2024, 44(1): 235-245
- http://dx.doi.org/10.20103/j.stxb.202210102868
-
文章历史
- 收稿日期: 2022-10-10
- 网络出版日期: 2023-09-28





2. 北京林业大学山西吉县森林生态系统国家野外科学观测研究站, 北京 100083;
3. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085
2. National Field Scientific Observation and Research Station of Forest Ecosystem in Jixian County, Shanxi Province, Beijing Forestry University, Beijing 100083, China;
3. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
IPCC第六次报告显示, 自1850—1900年以来, 全球地表的平均温度已上升约1℃, 并指出全球未来20年的平均气温预计升温或将超过1.5℃[1]。气候变暖会导致森林生态系统的结构和功能发生变化[2—3], 尤其是在中高纬度以及高海拔区域, 海拔通过影响水热分配, 进而影响树木的径向生长[4]。高原气候及其生态环境和地表特征对全球变化的响应敏感, 而且高原本身的变化反过来又作用于高原邻近地区, 甚至远离高原的地区。已有研究表明, 青藏高原是全球温度上升最快的区域之一, 该地区增温比全球变暖提前了大约30年[5—8]。澜沧江源区是青藏高原的一部分, 地域辽阔, 地形复杂, 是世界高海拔地区生物多样性最集中的自然保护区, 其独特的高寒生态系统非常脆弱, 是亚洲气候变化的“敏感区”和生态环境的“脆弱区”。因此对澜沧江源区近30年气候变化进行研究, 可为研究该区生态与人居环境对气候变化响应评估提供科学依据。
由于青藏高原气象站稀少、观测资料较短, 研究青藏高原气候高精细的时空变化明显地受器测气候资料缺乏的限制。而植被是连接大气圈、水圈、土壤圈和生物圈的重要纽带, 在陆地生态系统中表现活跃, 借助植被可以揭示树木径向生长对气候的敏感性, 恢复过去气候变化成为弥补器测气候资料不足的重要手段。树木年轮包含了树木历年的气候变化信息, 对气候变化信息具有很强的敏感性, 同时还具有定年准确、分辨率高、连续性强等优点[9], 已在我国的北方和西部被广泛应用于评估气候变化对树木径向生长影响的研究中[10—12]。有研究证明气候变化对不同区域不同树种的影响有很大差别, 例如刘晶晶等利用高原南部大果圆柏(Sabina tibetica)的树轮资料, 发现年总降水量是径向生长的主要限制因子[13], 而朱海峰等在青海省乌兰地区对祁连圆柏(Sabina przewalskii)的气候响应分析表明, 其径向生长主要由于温度变化受到限制[14]。王晓嘉等通过分析色季拉山区不同海拔方枝柏(Sabina saltuaria)侧枝的年轮宽度与气候的关系, 发现降水是年轮宽度的主要影响因子[15]。汪青春等通过建立青海高原不同区域的多个年表序列, 认为温度对径向生长的限制性最大, 尤其是最低气温[16]。田育红等在内蒙古大兴安岭对落叶松(Larix gmelinii)人工纯林的树木树轮指数(TRI)年表和胸高断面积增量(BAI)的分析表明, 森林在不同尺度上出现衰退现象[17]。潘磊通过分析气候对六盘山地区华北落叶松(Larix principis-rupprechtii)径向生长的影响, 发现当年降水量和年均温度分别达到820 mm和6.7℃时, 树木的BAI达到最大值, 二者继续增加则会抑制树木的径向生长[18]。以上研究的树种均为乔木, 近十年来, 越来越多的生态学者把灌木的生长和分布范围的变化作为对全球气候变化响应的敏感性生态指标[19]。相比于乔木, 分布于高寒区的灌木对高寒气候有着更好的适应性[20], 在大部分高海拔地区, 灌木种类较乔木更丰富, 并且灌木分布区域也是青藏高原人口分布集中区, 对于评估青藏高原人口主要分布区气候变化具有重要意义。本文通过研究高山柏径向生长过程及其与温度、降水等气候因素相关性, 可为气候暖湿化背景下, 开展青藏高原灌木生长对气候变化适宜性评估提供科技支撑。
高山柏(Sabina squamata)为深根性树种, 抗逆性强, 寿命长, 天然高山柏在澜沧江源区有广泛分布, 在水源涵养、水土保持、生物多样性保护和气候调控等方面发挥着重要作用。本文利用树木年轮气候学原理及方法, 以澜沧江源区的高山柏为研究对象, 建立高山柏标准年表、BAI年表, 揭示近30 a高山柏径向生长特征, 明确该区限制高山柏生长的主要气候因素, 研究可为开展气候变暖背景下澜沧江源区生态环境与人居气候环境评估提供支撑。
1 材料与方法 1.1 研究区概况研究在位于澜沧江源区的巴青县(94.28°E, 31.49°N, 海拔4121 m)、杂多县(95.21°E, 32.52°N, 海拔4080 m) 和囊谦县(96.25°E, 31.56°N, 海拔4309 m)开展高山柏的野外采样工作(图 1)。澜沧江源区是国际河流澜沧江(湄公河)的源头区, 河流自上而下发育由裸岩冰川、高寒草甸草原、灌木丛、大果圆柏林、湿地而组成的垂直植被地貌景观, 具有极为重要的水源涵养、生物多样性等生态服务功能[21]。澜沧江源区位于玉树藏族自治州杂多县, 该区域平均海拨4200 m以上, 高寒缺氧, 空气稀薄, 冬寒夏凉, 降水多集中在6—8月份(图 2), 年降水量约500—600 mm(图 3)。暖季只有6、7、8三个月, 而冷季却长达9个月(9—次年5月)[22]。年平均气温约为5.28℃(图 3), 澜沧江源区太阳辐射强, 日照时间长, 年平均日照时数为2310.3 h左右, 年平均蒸发量为1450.0 mm, 年平均风速为2.4 m/s, 最大风速大于40 m/s, 年大风日数达120 d, 无霜期短, 冬季多大风雪。研究区典型的高原气候特征使其成土过程缓慢, 土层薄且质地疏松, 保土能力弱, 土壤类型主要有高山寒漠土、高山草甸土、黑钙土、灰褐土等。澜沧江源区主要植被类型为高寒草甸类、高寒沼泽、高寒灌木草甸和高寒疏林, 常见灌木有高山柏、金露梅(Potentilla fruticosa)、银露梅(Potentilla glabra)等[23]。

|
| 图 1 澜沧江源区树木年轮采样点分布图 Fig. 1 Location map of tree-ring sample site in the Source Region of Lancang River |

|
| 图 2 1986—2019年巴青县、杂多县及囊谦县温度、降水季节分布 Fig. 2 Seasonal distribution of temperature and precipitation in Baqing County, Zaduo County and Nangqian County, China (1986—2019) |

|
| 图 3 1986—2019年澜沧江源区各气象要素的年际变化趋势 Fig. 3 Interannual trends of climatic factors in the Source Region of Lancang River from 1986 to 2019 |
在采样点选择长势良好、树龄较大、受病虫害干扰小的高山柏各取40个树盘, 将采集的样品根据国际树轮样品标准处理过程进行预处理。首先将采集的样品带回实验室晾干[24], 用不同粗细的干砂纸(400目、800目和1200目)依次打磨, 直到样品切面光滑明亮, 并且在电子显微镜下能看清高山柏的年轮。利用精度为0.001 mm的年轮分析仪Win DENDROTM(Regent instrument inc. Canada)测量高山柏树轮的原始宽度后, 用骨架图法交叉定年, 最后用COFECHA程序对交叉定年和测量结果进行校验, 剔除相关性差的样品。最后, 巴青、杂多、囊谦3个样地合格样品分别为34个、33个和40个, 以此完成三个样地的年表建立[25]。
由于样本树龄较短, 研究主要集中在树木生长的高频变化上, 故应去除树木随年龄增长的生长趋势和其他非一致性扰动导致的非气候信号, 而突出其中的气候信号[26]。利用ARSTAN程序建立年表, 去趋势方法为负指数函数, 该方法可减少树间竞争、地形、地貌等因素对年表指数拟合的影响, 最终选择去趋势后得到的标准年表(STD)进行数据分析。采用子样本信号强度SSS>0.85的样本量作为年表起始点, 高于样本量的年表序列具有可靠性[27], 巴青县、杂多县和囊谦县高山柏年表的可靠区间分别为1986—2019年、1979—2019年和1900—2019年, 取公共时段1986—2019年对高山柏径向生长趋势进行分析。
1.3 胸高断面积增量当树轮宽度相对稳定或变窄时, 树木生长量还会继续增大。为了保留气候条件对树木生长影响的信息, 本文使用BAI来表征研究区内高山柏的生长速率和生产力变化趋势。树木胸高断面积增量:
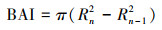
|
式中, Rn是n年的树轮半径(mm); Rn-1是n-1年的树轮半径(mm)。
1.4 气象数据及统计分析气象资料来源于中国气象数据网(http://data.cma.cn), 从靠近采样点的巴青站(94.03°E, 31.93°N, 海拔4174 m)、杂多站(95.17°E, 32.53°N, 海拔4066.4 m)和囊谦站(96.28°E, 32.12°N, 海拔3643.7 m)获取1986—2019年月降水量(PRE)、月平均气温(TMP)、月平均最低气温(TMN)、月平均最高气温(TMX)和月平均相对湿度(RH)气象数据。由于温度和降水等综合作用对土壤有效含水量的影响明显, 本研究选用帕默尔干旱指数(Palmer drought index, PDSI)反映高山柏生长与土壤水分的关系, 其数据来源于荷兰皇家气象局服务网(http://climexp.knmi.nl/start.cgi, 0.5°×0.5°)中的PDSI网格点数据集[28], 分别从距采样点最近的CUR网格点(94.00°—94.50°E, 31.00°—31.50°N; 95.00°—95.50°E, 32.50°—33.00°N; 96.00°—96.50°E, 31.50°—32.00°N)选取1986—2019年的PDSI格点数据。考虑到树木生长的连续性, 当年的树木生长还受到前一年气候的滞后影响, 在进行气候响应分析时选用上一年6月到当年10月共17个月的气象数据。本文利用Excel 2019软件分析三个样地高山柏的径向生长过程, 使用Dendroclim 2002国际树木年轮程序[29]分析年表与各气候要素间的相关性。
1.5 年表特征分析 1.5.1 序列间相关系数所有样本间的相关性, 又叫序列间相关系数。它是反映树木受气候变化而产生的生长量变化的同步性系数, 其值越大, 年表的质量越高, 年表中所含的气候信息越丰富[26]。
1.5.2 标准差树轮年表的标准差是反应年表中所含的气候信息含量的另外一个重要指标。标准差是随机抽取样本序列中的一定量。计算其标准误差所得到的数值。标准差反映年表的上下波动, 其值越大表明年表所包含的气候信息越多, 对气候越敏感[30]。一般来说, 优质的年表, 标准差均在0.2以上。
1.5.3 一阶自相关系数一阶自相关系数反映上一年气候状况对当年树轮宽度生长的影响强弱, 具有连续性和滞后性的特点。如果上一年的气候对树木生长的影响越大, 相应的一阶自相关系数就越高[31]。
1.5.4 平均敏感度平均敏感度是衡量树轮年表质量的重要指标之一, 其表达式为:

|
式中, xi表示第i年的树轮宽度值; xi+1是第i+1年的树轮宽度值; n为年表中的样表总数。平均敏感度是一个无量纲的指数, 主要反映的是该年表序列对气候信息的敏感程度。平均敏感度数值越大, 则说明该年表序列受气候因子的限制性作用越强, 与气候信息的相关性就越好。年表中所包含的气候信息就更加丰富。根据Ferguson的分类, 大于0.3的平均敏感度被认为是高的[32]。
1.5.5 信噪比信噪比反应的是在序列所包含的信息总量中, 有用信息和噪音音系的比例。通过方差分析可以得到总年表平均指数的方差贡献, 而剩余的即为噪音, 二者的比值被定义为“信噪比”。该指标主要代表气候的短期变化或高频变化。
1.5.6 样本总体代表性样本对总体的代表性是指所采集样本对整个区域的代表程度。样本对总体的代表性越高, 则说明样本对总体的代表性越好[33]。
2 结果 2.1 气候年际变化趋势自1986年以来, 澜沧江源区年平均气温以0.43℃/10a(P < 0.01)的速度上升(图 3), 年平均最低气温(0.57℃/10a, P < 0.01)的上升速率明显高于年平均最高气温(0.33℃/10a, P < 0.01);1986—2019年澜沧江源区年降水量上升速率为2.39 mm/a(P>0.10, 图 3), 呈波动特征, 这与许学莲等[34]在三江源地区对气温和降水变化的研究相符, 青藏高原不同地区海拔高度和下垫面的差异是导致澜沧江源区增温幅度较大的主要原因。研究区的相对湿度呈不显著的下降趋势(图 3), PDSI呈现略微上升趋势(0.27/10a, P < 0.10), 多年份的PDSI值大于-2, 说明该地区有效土壤水分较高, 并未出现明显的气候暖干化特征(图 3)。
2.2 树轮年表统计特征本研究建立了澜沧江源区3个样地的高山柏标准年表, 年表质量的高低决定着研究结果是否准确, 利用序列间相关系数、标准差、一阶自相关系数、平均敏感度、信噪比、样本总体代表性等来评价年表质量(表 1)。
| 类型 Type |
那曲巴青县 Naqu Baqing County |
玉树杂多县 Yushu Zaduo County |
玉树囊谦县 Yushu Nangqian County |
| 地点Location | 94.28°E, 31.49°N | 95.21°E, 32.52°N | 96.25°E, 31.56°N |
| 海拔Elevation/m | 4121 | 4080 | 4309 |
| 年平均温度Temperature/℃ | 7.03 | 3.89 | 4.91 |
| 年降水量Precipitation/mm | 687.93 | 539.78 | 579.77 |
| 年表总长度Total chronology length | 1981—2019 | 1952—2019 | 1880—2019 |
| 可靠区间Reliable range | 1986—2019 | 1979—2019 | 1900—2019 |
| 样本量Sample size/n | 34/40 | 33/40 | 40/40 |
| 标准差SD | 0.310 | 0.262 | 0.309 |
| 平均敏感度MS | 0.175 | 0.185 | 0.114 |
| 一阶自相关系数AR1 | 0.859 | 0.722 | 0.719 |
| 样本间相关系数Rbt | 0.395 | 0.252 | 0.241 |
| 样本总体代表性EPS | 0.925 | 0.814 | 0.888 |
| 信噪比SNR | 12.395 | 4.386 | 7.959 |
| MS:平均敏感度Mean sensitivity; SD:标准差Standard deviation; AR1:一阶自相关First-order autocorrelation; Rbt:样本间相关系数Mean inter-series correlation; SNR:信噪比Signal-to-noise ratio; EPS:样本总体代表性Express population siginal | |||
巴青县、杂多县和囊谦县高山柏标准年表的序列间相关系数分别为0.395、0.252、0.241, 表示不同树盘间的年轮宽度变化具有较好的一致性, 数据质量较好; 标准差分别为0.310、0.262、0.309, 均大于0.2, 说明该年表中所包含的气候信息较为丰富; 一阶自相关系数分别为0.859、0.722、0.719, 可见树龄越小的高山柏受气候影响越为强烈; 平均敏感度分别为0.175、0.185、0.114, 均小于0.3, 表明树轮宽度变异性相对较低; 信噪比分别为12.395、4.386、7.959, 达到了较高水平, 表明包含丰富的气候信号; 样本总体代表性分别为0.925、0.814、0.888, 巴青县、囊谦县高山柏样本总体代表性大于0.85, 表明其年表质量较杂多县高山柏年表质量高。
综上所述, 澜沧江源区高山柏年表质量相对较好, 符合树轮气候学分析要求。以下运用标准年表来进行高山柏径向生长对气候变化的响应分析。高山柏标准年表的起始年份样本量大于5根, 可以较好地反映径向生长年际波动, 巴青县、杂多县和囊谦县高山柏的年表变化特征曲线如图 4所示。对比发现, 囊谦县高山柏年表序列时间跨度最长(1880—2019), 有效年表长度为120年(1900—2019);巴青县和杂多县高山柏年表序列相对较短, 其中巴青县高山柏有效年表长34年(1986—2019), 杂多县高山柏有效年表长41年(1979—2019)。在公共区间(1986—2019年)内, 巴青县和杂多县高山柏树轮宽度指数上升趋势较为显著, 囊谦县高山柏年表上升趋势稍显平缓, 表明巴青县和杂多县高山柏径向生长对气候因子的敏感性较囊谦县高。

|
| 图 4 澜沧江源区高山柏标准年表 Fig. 4 Standard chronology of Sabina squamata in the Source Region of Lancang River SSS为子样本信号强度Subsample signal strength |
气候变暖背景下, 澜沧江源区高山柏树轮宽度呈现显著上升趋势(P < 0.01, 图 5)。其中囊谦县采样点所处海拔较高(4309 m), 且高山柏树龄较大(120 a), 其生长比较慢, 1986—1999年以(0.11±0.01) mm/a的速度缓慢生长, 2000—2015年的生长速率略微上升((0.17±0.03) mm/a), 相对前期增长了54%。巴青县和杂多县采样点的海拔更为相近(分别为4121 m和4080 m), 且高山柏树龄较小(34 a、41 a), 二者的树轮宽度变化趋势相似, 都经历了显著上涨过程, 2000—2015年二者的平均生长速率((0.46±0.09) mm/a)较1986—1999年的生长速率((0.25±0.07) mm/a)增长了84%。2016年后, 三个样地的高山柏树轮宽度均呈下降趋势。

|
| 图 5 澜沧江源区高山柏树轮宽度和胸高断面积增量(BAI)年际变化 Fig. 5 Interannual variation of Sabina squamata tree-ring width and BAI in the Source Region of Lancang River |
BAI是每年树轮增加的面积, 相比年轮宽度, 它能够更好地反应树木生长速率和生长趋势[35]。囊谦县高山柏BAI在1986—1999年呈现稳定生长状态((6.65±0.70) mm2/a), 2000—2015年在逐步上升((11.89±3.20) mm2/a)。巴青县和杂多县高山柏的BAI在1986—1999年呈逐步上升趋势, 平均以(14.92±4.11) mm2/a的速度生长, 2000—2015年处于快速稳定生长的特征, 其生长速率平均为(27.49±5.33) mm2/a, 在此期间, 澜沧江源区高山柏BAI急速上升, 表明当地高山柏生产力进入旺盛生长阶段, 尚未出现衰退现象。2016年后, 三个样地的高山柏BAI均呈下降趋势(图 5)。
2.4 径向生长与月气候因子的相关性标准年表与气候因素相关性分析表明:澜沧江源区高山柏径向生长与多数月份的温度呈显著正相关, 与多数月份降水呈不显著的正相关(图 6)。三个样地高山柏径向生长均与上年及当年生长季(6—8月)、当年生长初期(4、5月)、生长末期(9—10月)和冬季(12—次年2月)的温度呈显著正相关, 特别是与月平均最低温度关系最为密切(P < 0.01), 只与春季(1、2月)和生长季(7月)的降水量呈不显著负相关(P>0.05, 图 6)。帕默尔干旱指数在干旱区的研究中广泛应用, 它是根据降水、温度以及土壤含水量数据进行计算, 代表土壤水分的有效状况, 其值越小表明有效土壤水分越低[36]。巴青县和杂多县高山柏径向生长与上年6月—当年10月的PDSI正相关, 囊谦县高山柏径向生长与之负相关, 但均未达到显著水平(图 6), 说明澜沧江源区有效土壤水分较高, 高山柏径向生长并未明显地遭受干旱胁迫。

|
| 图 6 澜沧江源区高山柏标准年表与月平均温度、月平均最低温度、月平均最高温度、月平均降水量、PDSI的相关系数 Fig. 6 Correlation coefficients between the standard chronology of Sabina squamata and monthly mean temperature, monthly mean minimum temperature, monthly mean maximum temperature, monthly mean precipitation and PDSI in the Source Region of Lancang River 虚线表示0.05水平的显著, 实线表示0.01水平的显著; 月份前的“-”表示上一年, 如-6表示上一年6月份 |
研究区巴青县、杂多县和囊谦县具有明显的气候相似性, 通过分析三个样地高山柏径向生长特征, 对澜沧江源区高山柏生长的气候适宜性研究有重要意义。由于囊谦县高山柏的采样点所处海拔高, 树龄大; 巴青县和杂多县高山柏采样点所处海拔相近, 较囊谦县低, 且树龄小。研究发现巴青县和杂多县高山柏树轮宽度变化趋势相似, 都经历了显著上涨过程, 二者在2000—2015年的平均生长速率((0.46±0.09) mm/a)较1986—1999年的生长速率((0.25±0.07) mm/a)增长了84%, 而囊谦县高山柏树轮在1986—2015年间呈平稳生长状态((0.14±0.04) mm/a), 说明气候变化对树龄较小的高山柏影响较为显著, 高山柏在生长过程中对温度和水分的敏感性更高, 囊谦县的高山柏随着树龄的增大, 其径向生长速率趋于稳定, 独特的低温高湿环境使高山柏的生长波动较小, 雷帅等[37]在毛乌素和塞罕坝对樟子松(Pinus sylvestris var. mongolica)也有相似的研究结果。此外, 朱明阳等[38]在云南桥子山高山树线交错带对不同海拔树线树种急尖长苞冷杉(Abies georgei var. smithii)的径向生长监测表明, 随海拔升高, 急尖长苞冷杉径向生长的开始时间推迟、结束时间提前, 生长季缩短, 从而很好地解释了囊谦县高山柏径向生长较慢的原因。BAI也快速上升, 2000—2015年处于平稳的快速增长阶段((22.29±8.74) mm2/a), 说明高山柏生产力稳定且旺盛, 该区气候条件适宜高山柏的径向生长, 刘亚玲等[39]在河北坝上对樟子松也有相似的研究结果。2016年之后, 三个样地的高山柏树轮宽度和BAI均呈下降趋势, 这可能与澜沧江源区的气候有关, 该区域的温度同样在2016年有所降低(图 3)。此外, 许学莲[34]等在三江源地区对气候的研究中发现, 近几年温度也出现了下降趋势。从三个样地高山柏逐年BAI增长量分析, 由于本文章中巴青县和杂多县高山柏的树龄较小, 仅有30—40 a, 而理论上高山柏的最大树龄可达千年, 所以该样地高山柏BAI年际增长量变化趋势不完全反映当地高山柏生产力是否出现下降现象, 还需要对研究区高山柏进行连续观测。
3.2 高山柏径向生长对气候因子的响应标准年表与月气象因子的相关性分析表明:澜沧江源区高山柏径向生长对温度的响应明显强于降水。三个样地高山柏径向生长均与多数月份(上年6月—当年10月)的温度显著正相关, 特别是月平均最低温度。高山柏径向生长与生长初期(4、5月)最低温度显著正相关, 朱良军等[40]在小兴安岭对阔叶红松(Pinus koraiensis)和臭冷杉(Abies nephrolepis)及李广起等[41]在长白山自然保护区对红松和鱼鳞云杉(Picea jezoensis var. komarovii)都有相似的研究结果, 生长季前充足的热量是树木形成宽轮的主要原因之一。因为3、4月即初春是树木结束休眠转入生长季的重要时期, 平均温度回升有利于树木打破休眠, 对高山柏的初期形成层细胞的分裂有显著促进作用[42]。澜沧江源区春季开始时间较晚, 故4月最低温度对高山柏径向生长的促进作用较为合理; 高山柏径向生长与生长季(6—8月)、生长末期(9—10月)最低温度显著正相关, 因为6—9月即生长季初夏到初秋是树木生长最旺盛的时期, 也是季风降水的高峰期, 最低温度的回升有利于树木的光合作用, 从而产生足够的光合产物供树木生长, 而温度过低则限制其光合[43]。10月份即生长季后期形成层依然在活动, 温度和降水继续增加有利于树木进行光合作用, 累积养分[44]。高山柏径向生长与冬季(12—次年1月)最低温度显著正相关, 冬季温度偏高时, 一方面可以避免破坏叶肉组织, 保证来年高山柏的正常代谢活动; 另一方面可降低树木休眠时维持生理活动所需能量, 减少能量消耗, 促进树木来年生长[45—46]。对比发现, 巴青县和杂多县高山柏径向生长对温度的响应较囊谦县略强, 说明气温对树木生长的早期影响较大, 树龄较小的高山柏在生长过程中需要更多的养分和能量, 宋来萍等[47]在呼伦贝尔沙地对不同树龄樟子松也有相似的研究结果。此外, 海拔的升高伴随着温度的下降和降水的增加, 再加上立体条件、个体区别、人为影响等差异, 温度、降水对树轮宽度的影响是综合及复杂的。当降水量低于一定值时, 温度的高低和树轮宽度的生长呈现明显的负相关, 而且降水量越少这种负相关性就越显著; 在降水适宜的情况下, 温度对树木生长的影响不明显[15]。囊谦县采样点所处海拔虽然较巴青县和杂多县高, 但三个样地年降水量均在500 mm以上(图 3), 且当地高山柏径向生长与多数月份(上年6月—当年10月)的降水量呈弱的正相关, 表明该区域降水量丰沛, 降水不是高山柏生长的主要限制性气候因素, 所以澜沧江源区高山柏径向生长对气候的响应受海拔因素的影响较小。
澜沧江源区平均海拔在4200 m以上, 常年处于低温高湿环境, 热量不足是高山柏生长的限制性因素, 最低温度的变化会显著影响高山柏的光合作用, 进而影响径向生长速率。因此, 在气候暖湿化背景下, 植被生长加速, 植被覆盖条件得到改善。从气候条件来看, 当地更加适宜人类居住。
4 结论本文以澜沧江源区的巴青县、杂多县和囊谦县高山柏为研究对象, 利用树木年轮学方法建立三个样地高山柏年轮宽度标准年表, 探讨了高山柏树轮宽度和胸高断面积增量的年际变化特征及对温度、降水等气候因素的响应。研究发现1986—2019年澜沧江源区年平均气温呈快速上升趋势(0.43℃/10a, P < 0.01), 而年降水量呈弱的上升状态。在气候变暖背景下, 澜沧江源区高山柏树轮宽度呈现上升趋势, 其中2000—2019年的径向生长速率为(0.37±0.16) mm/a, 较1986—1999年的生长速率((0.20±0.09) mm/a)增长了85%, 而BAI在前期呈逐步上升趋势, 在2000—2019年呈平稳的快速生长特征((22.49±8.84) mm2/a)。澜沧江源区高山柏径向生长对温度的响应明显强于降水, 特别是与月平均最低温度的相关性最为显著。澜沧江源区气候变暖驱动下高山柏进入了生长适应期, 树轮宽度和胸高断面积增量均呈快速生长特征, 表明该区域气候条件更加适宜高山柏生长。
| [1] |
Landman W. Climate change 2007: the physical science basis. South African Geographical Journal, 2010, 92(1): 86-87. DOI:10.1080/03736245.2010.480842 |
| [2] |
刘国华, 傅伯杰. 全球气候变化对森林生态系统的影响. 自然资源学报, 2001, 16(1): 71-78. DOI:10.3321/j.issn:1000-3037.2001.01.013 |
| [3] |
Zhu L J, Wang X C, Pederson N, Chen Z J, Cooper D J, Zhang Y D, Li Z S. Spatial variability in growth-climate relationships of Amur cork tree (Phellodendron amurense) and their connections WithPDOin northeast China. Journal of Geophysical Research: Biogeosciences, 2018, 123(5): 1625-1636. DOI:10.1029/2017JG004292 |
| [4] |
张慧, 邵雪梅, 张永. 不同海拔高度树木径向生长对气候要素响应的研究进展. 地球环境学报, 2012, 3(3): 845-854. |
| [5] |
Piao S L, Ciais P, Huang Y, Shen Z H, Peng S S, Li J S, Zhou L P, Liu H Y, Ma Y C, Ding Y H, Friedlingstein P, Liu C Z, Tan K, Yu Y Q, Zhang T Y, Fang J Y. The impacts of climate change on water resources and agriculture in China. Nature, 2010, 467(7311): 43-51. DOI:10.1038/nature09364 |
| [6] |
杨耀先, 胡泽勇, 路富全, 蔡英, 于海鹏, 郭瑞霞, 付春伟, 樊威伟, 吴笛. 青藏高原近60年来气候变化及其环境影响研究进展. 高原气象, 2022, 41(1): 1-10. |
| [7] |
冯松, 姚檀栋, 江灏, 康兴成, 汤懋苍. 青藏高原近600年的温度变化. 高原气象, 2001, 20(1): 105-108. DOI:10.3321/j.issn:1000-0534.2001.01.018 |
| [8] |
李跃清. 近40年青藏高原东侧地区云、日照、温度及日较差的分析. 高原气象, 2002, 21(3): 327-332. DOI:10.3321/j.issn:1000-0534.2002.03.016 |
| [9] |
吴祥定. 树木年轮与气候变化. 北京: 气象出版社, 1990.
|
| [10] |
Bräuning A. Dendrochronology for the last 1400 years in eastern Tibet. GeoJournal, 1994, 34(1): 75-95. DOI:10.1007/BF00813972 |
| [11] |
王晓明, 赵秀海, 高露双, 姜庆彪. 长白山北坡不同年龄红松年表及其对气候的响应. 生态学报, 2011, 31(21): 6378-6387. |
| [12] |
石松林, 靳甜甜, 刘国华, 王东波, 王景升, 李魁. 气候变暖抑制西藏拉萨河大果圆柏树木生长. 生态学报, 2018, 38(24): 8964-8972. DOI:10.5846/stxb201807131522 |
| [13] |
刘晶晶, 杨保. 青藏高原南部南木林地区树木径向生长对气候因子的响应. 中国沙漠, 2011, 31(6): 1527-1534. |
| [14] |
朱海峰, 郑永宏, 邵雪梅, 刘晓宏, 徐岩, 梁尔源. 树木年轮记录的青海乌兰地区近千年温度变化. 科学通报, 2008, 53(15): 1835-1841. DOI:10.3321/j.issn:0023-074X.2008.15.014 |
| [15] |
王晓嘉, 张占芳, 郑维列. 不同海拔方枝柏侧枝年轮对温度和降水响应的初步研究. 西藏科技, 2012(10): 53-57. DOI:10.3969/j.issn.1004-3403.2012.10.025 |
| [16] |
汪青春, 秦宁生, 李栋梁, 时兴合, 王振宇, 李林, 朱西德. 利用多条树轮资料重建青海高原近250年年平均气温序列. 高原气象, 2005, 24(3): 320-325. DOI:10.3321/j.issn:1000-0534.2005.03.006 |
| [17] |
田育红, 刘怡青, 吴秀臣, 颜孟萱, 刘凤花, 栾静雅. 内蒙古东北段森林衰退现状及种群竞争对其生长的影响. 生态学报, 2022, 42(17): 7222-7233. DOI:10.5846/stxb202106101549 |
| [18] |
潘磊. 华北落叶松径向生长对水热条件及竞争的响应[D]. 哈尔滨: 东北林业大学.
|
| [19] |
肖生春, 彭小梅, 丁爱军, 田全彦, 韩超. 中国寒旱区灌木年轮学研究进展. 地球科学进展, 2020, 35(6): 561-567. |
| [20] |
芦晓明, 梁尔源. 灌木年轮学研究进展. 生态学报, 2013, 33(5): 1367-1374. DOI:10.5846/stxb201208031105 |
| [21] |
Mushi C A, Ndomba P M, Trigg M A, Tshimanga R M, Mtalo F. Assessment of basin-scale soil erosion within the Congo River Basin: a review. CATENA, 2019, 178: 64-76. DOI:10.1016/j.catena.2019.02.030 |
| [22] |
梅志宏. 走进湄公河. 中国三峡, 2017(9): 58-71. DOI:10.3969/j.issn.1006-6349.2017.09.008 |
| [23] |
吴玉虎. 澜沧江源区种子植物区系研究. 武汉植物学研究, 2009, 27(3): 277-289. |
| [24] |
Stokes M A, Smiley T L. An introduction to tree-ring dating. Chicago: University of Chicago Press, 1968.
|
| [25] |
Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43: 51-67. |
| [26] |
Brubaker L. Tree rings and climate. Ecology, 1977, 58(6): 1400-1401. DOI:10.2307/1935092 |
| [27] |
贾飞飞, 孙翠洋, 孙红月, 李鑫. 昌岭山两个优势树种径向生长对气候变化的响应. 生态学报, 2019, 39(17): 6332-6340. DOI:10.5846/stxb201805181095 |
| [28] |
Harris I, Jones P D, Osborn T J, Lister D H. Updated high-resolution grids of monthly climatic observations-the CRU TS3.10 Dataset. International Journal of Climatology, 2014, 34(3): 623-642. DOI:10.1002/joc.3711 |
| [29] |
Biondi F, Waikul K. DENDROCLIM2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers & Geosciences, 2004, 30(3): 303-311. |
| [30] |
Fritts H C. Tree rings and climate. London: Academic Press, 1976.
|
| [31] |
孙铂, 雷莺, 郭玲霞, 刘宇峰, 王倩. 秦岭通天河地区树轮年表的建立及特征分析. 咸阳师范学院学报, 2019, 34(4): 58-60. DOI:10.3969/j.issn.1672-2914.2019.04.015 |
| [32] |
Savva Y, Oleksyn J, Reich P B, Tjoelker M G, Vaganov E A, Modrzynski J. Interannual growth response of Norway spruce to climate along an altitudinal gradient in the Tatra Mountains, Poland. Trees, 2006, 20(6): 735-746. DOI:10.1007/s00468-006-0088-9 |
| [33] |
Wigley T M L, Briffa K R, Jones P D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. Journal of Climate and Applied Meteorology, 1984, 23(2): 201-213. DOI:10.1175/1520-0450(1984)023<0201:OTAVOC>2.0.CO;2 |
| [34] |
许学莲, 李积芳, 李金凤, 文生祥, 李春晖. 三江源地区气温和降水变化对气候生产潜力的影响. 青海草业, 2022, 31(4): 45-49. |
| [35] |
刘敏. 中国东北红松生长对气候变化的响应及其动态研究[D]. 哈尔滨: 东北林业大学.
|
| [36] |
卫捷, 陶诗言, 张庆云. Palmer干旱指数在华北干旱分析中的应用. 地理学报, 2003, 58(S1): 91-99. |
| [37] |
雷帅, 张劲松, 孟平, 何子兵, 程顺, 孙守家. 中国北部不同地点樟子松人工林径向生长对气候响应的差异. 生态学报, 2020, 40(13): 4479-4492. DOI:10.5846/stxb201911152437 |
| [38] |
朱明阳, 林琳, 佘雨龙, 肖城材, 赵通兴, 胡春相, 赵昌佑, 王文礼. 云南轿子山不同海拔急尖长苞冷杉径向生长动态及其低温阈值. 植物生态学报, 2022, 46(9): 1038-1049. |
| [39] |
刘亚玲, 信忠保, 李宗善, 买尔当·克依木, 闫腾飞. 河北坝上樟子松人工林径向生长及其对气候因素的响应. 生态学报, 2022, 42(5): 1830-1840. DOI:10.5846/stxb202010272750 |
| [40] |
朱良军, 杨婧雯, 朱辰, 王晓春. 林隙干扰和升温对小兴安岭红松和臭冷杉径向生长的影响. 生态学杂志, 2015, 34(8): 2085-2095. |
| [41] |
李广起, 白帆, 桑卫国. 长白山红松和鱼鳞云杉在分布上限的径向生长对气候变暖的不同响应. 植物生态学报, 2011, 35(5): 500-511. |
| [42] |
常永兴, 陈振举, 张先亮, 白学平, 赵学鹏, 李俊霞, 陆旭. 气候变暖下大兴安岭落叶松径向生长对温度的响应. 植物生态学报, 2017, 41(3): 279-289. |
| [43] |
贺敏, 魏江生, 石亮, 周梅, 赵鹏武. 大兴安岭南段山杨径向生长和死亡对区域气候变化的响应. 生态学杂志, 2018, 37(11): 3237-3244. |
| [44] |
韦景树, 李宗善, 焦磊, 陈维梁, 伍星, 王晓春, 王帅. 黄土高原羊圈沟小流域人工物种和自然物种径向生长对气候变化的响应差异. 生态学报, 2018, 38(22): 8040-8050. DOI:10.5846/stxb201801100079 |
| [45] |
朱良军, 金光泽, 王晓春. 典型阔叶红松林干扰历史重建及干扰形成机制. 植物生态学报, 2015, 39(2): 125-139. |
| [46] |
Wang H, Shao X M, Fang X Q, Jiang Y, Liu C L, Qiao Q. Relationships between tree-ring cell features of Pinus koraiensis and climate factors in the Changbai Mountains, Northeastern China. Journal of Forestry Research, 2017, 28(1): 105-114. |
| [47] |
宋来萍, 刘礴霏, 王玉华, 高敬泽. 呼伦贝尔沙地不同树龄樟子松对气候的响应. 南京林业大学学报: 自然科学版, 2020, 44(2): 159-164. |