 2023, Vol. 43
2023, Vol. 43文章信息
- 张杰京, 陈飞, 谢菲, 张鑫, 尹文萍, 樊辉
- ZHANG Jiejing, CHEN Fei, XIE Fei, ZHANG Xin, YIN Wenping, FAN Hui
- 亚洲象生境长时序变化及其对人象冲突的影响——基于融合MaxEnt与HSI模型的生境适宜性评价方法
- Long time series changes of Asian elephant habitat and impacts on human-elephant conflict: based on habitat suitability evaluation method by combining MaxEnt and HSI models
- 生态学报. 2023, 43(9): 3807-3818
- Acta Ecologica Sinica. 2023, 43(9): 3807-3818
- http://dx.doi.org/10.5846/stxb202205101297
-
文章历史
- 收稿日期: 2022-05-10
- 网络出版日期: 2022-09-19






2. 云南省国际河流与跨境生态安全重点实验室, 昆明 650091;
3. 国家林业和草原局亚洲象研究中心, 昆明 650216;
4. 国家林业和草原局西南调查规划院, 昆明 650216;
5. 云南亚洲象教育部野外科学观测研究站, 昆明 650504
2. Yunnan Key Laboratory of International Rivers and Transboundary Eco-Security, Yunnan University, Kunming 650091, China;
3. Asian Elephant Research Center of National Forestry and Grassland Administration, Kunming 650216, China;
4. Southwest Survey and Planning Institute of National Forestry and Grassland Administration, Kunming 650216, China;
5. Yunnan Asian Elephant Field Scientific Observation and Research Station of the Ministry of Education, Kunming 650504, China
人类活动及全球气候变化导致的生境丧失与破碎化正严重威胁着生物多样性[1—3], 生境作为物种生存、繁衍的重要场所, 对物种生存具有重要意义[4]。物种保护不仅要考虑物种本身, 还要考虑其所在生境景观的变化特征[5]。因而, 悉知物种生境景观格局的历史变化动态及物种对生境变化的响应行为对物种的保护尤为重要。物种分布模型(Species distribution models, SDMs)是当前较为常用的生境适宜性评价方法, 其基于物种的活动点数据和环境因子数据构建数学模型, 达到量化物种生境需求和确定其潜在适宜生境范围的目的[6];目前常用的物种分布模型包括生态位因子分析(Ecological Niche Factor Analysis, ENFA)模型[7], 最大熵(Maximum Eentropy, MaxEnt)模型[8], 贝叶斯网络(Bayesian network, BN)模型[9]等, 但受物种活动点数据在时间空间上的限制, 这类模型不能进行长时序的物种生境评价[10—11]。生境适宜性指数(Habitat suitability index, HSI)模型是基于地理信息系统(GIS)建立的生境评价方法, 因不依赖于物种活动点数据即可实现长时序的生境适宜性评价, 并且被广泛应用于物种活动点数据稀缺地区[12], 但是该模型缺乏生态位理论知识支撑[13]。受物种活动点数据的时间和空间定位精度限制, 定量获取物种历史生境偏好较为困难, 导致以往的生境适宜性评价多集中于某单一时间节点[7, 14—15], 缺乏对物种生境长时序变化的研究。而单一时间节点的生境适宜性评价已无法满足全球环境变化背景下的物种生境修复、生境变化归因和人兽冲突缓解等方面的应用需求[16]。物种生境偏好具有较强的延续性, 极少发生突变。因此, 可假设物种生境偏好在时间上具有一致性, 并以此进行生境评价。这为物种长时序生境制图提供了契机, 同时还避免了以往研究中采用历史物种活动点数据存在的严重空间误差对亚洲象生境偏好以及生境适宜性评价的影响[17—18]。近年来, 随着地球观测技术的进步, 长时序地理空间数据为物种长时序生境制图提供了坚实的数据基础[19]。因此, 基于长时序的遥感与地理空间数据, 构建一套适用于物种长时序生境评价的方法, 服务于物种生境的保护、管理及未来规划等, 具有重要意义。
亚洲象(Elephas maximus)是亚洲热带森林生态系统的重要物种[20—21], 对维持区域生物多样性具有重要意义[22]。我国野生亚洲象主要分布于云南南部西双版纳、普洱、临沧等地区[15], 数量约300头[23]。西双版纳勐海—普洱澜沧地区亚洲象种群(勐海—澜沧象群)的活动范围涉及西双版纳勐海县的勐阿镇、勐往镇、勐满镇和普洱市澜沧县的发展河乡、惠民镇等地。该象群最早记录于20世纪80年代(4头)由勐养子保护区迁移至此, 随后因景洪水电站蓄水, 象群渡江后无法返回原栖息地[24], 而频繁往返于勐海县和澜沧县交界地带, 截止到2018年象群个体数量已增至19头[14, 24—25]。随象群个体数量增长, 其活动范围不断扩大, 并与人类生产、生活空间重叠, 导致人象冲突频发。2011—2019年间, 共造成27人死亡, 多人受伤[24], 严重影响了当地居民生产生活。目前对象群活动区内生境质量与格局的认知极为有限, 严重掣肘了亚洲象生境保护与恢复以及人象冲突的缓解。近几十年来, 受经济作物种植扩张影响, 该区域亚洲象生境变化剧烈, 且近年来人象冲突愈发频繁, 亟需探究亚洲象生境景观的长期变化特征及其对人象冲突的影响, 揭示生境景观变化与亚洲象肇事间的作用机制。因此, 本研究假设研究时段内亚洲象生境偏好保持不变, 定量研究环境变化对亚洲象生境的影响。研究基于2020—2021年亚洲象活动点监测数据与研究区高精度逐年土地利用/覆被数据集(1988—2020年), 将MaxEnt模型获取的生境因子重要性融入HSI模型, 构建融合MaxEnt与HSI模型的生境适宜性评价方法, 并以勐海—澜沧象群活动区为例, 制作1988—2020年逐年亚洲象生境适宜性图, 分析亚洲象生境的时空变化过程, 定量揭示环境变化对物种生境景观的影响;在此基础上, 结合2020年亚洲象肇事公众责任保险理赔数据, 探究生境景观变化与人象冲突间的内在关系, 以期为亚洲象生境管理与保护和人象冲突缓解提供科学支撑。
1 研究区概况研究区(图 1)位于云南省南部(99°41′—100°57′E, 21°28′—22°47′N), 总面积约12000 km2。该区属于怒山山脉向南延伸余脉部, 区内地形复杂, 地势呈现中部平缓, 四周隆起, 海拔535—2521 m。研究区位于热带与亚热带的过渡带, 属亚热带季风气候, 年平均降水量1300—1500 mm, 年平均温度17—20℃[26]。西双版纳勐海—普洱澜沧亚洲象种群自2005年后定栖于此区域[24—25, 27], 种群活动区变化大致经历了三个阶段:1992—2005年北上阶段, 象群从西双版纳勐养保护区向北迁移, 后渡过澜沧江进入澜沧县糯扎渡镇, 并常迁移于澜沧江东、西岸;2005—2011年南下阶段, 象群滞留澜沧江西岸, 后进入澜沧县发展河乡, 主要活动于糯扎渡镇与发展河乡交界区域;2011年之后为外扩阶段, 象群由发展河乡进入勐海县勐往乡、勐阿镇一带活动, 并以发展河乡、勐往乡、勐阿镇为中心向外扩张, 活动范围包括澜沧县酒井乡、惠民镇和勐海县勐满镇、勐遮镇等[25](图 1)。

|
| 图 1 研究区地理位置及亚洲象迁移路线 Fig. 1 Geographical location of the study area and migration routes of the Asian elephant herd |
使用数据包括:1988—2020年土地利用/土地覆被数据、地形、道路、河流、降水、归一化差异植被指数(NDVI)等数据。1988—2020年土地利用/土地覆被数据基于Landsat TM/ETM+/OLI时序影像, 结合野外调查情况绘制分类样本, 利用随机森林分类器进行监督分类, 得到1988—2020年时序土地利用/土地覆被分类图, 总体精度大于85%(未发表数据);地形数据来自美国航天雷达地形测绘任务的数字高程模型(SRTM DEM) (https://earthexplorer.usgs.gov/);1988—2020年时序NDVI数据来自谷歌云平台以Landsat 5 TM、Landsat 7 ETM+、Landsat 8 OLI影像为基础计算的年度NDVI数据集(https://developers.google.com/earth-engine/datasets/catalog);降水数据由中国科学院水利部成都山地灾害与环境研究所数字山地与遥感应用中心根据CRU(Climatic Research Unit)数据集(https://sites.uea.ac.uk/cru/)进行降尺度与空间插值得到, 为1991—2020年多年平均降水数据;距河流距离数据根据云南省1:25万地形图河流矢量数据计算得到;距道路距离数据根据全国地理信息资源目录服务系统提供的道路矢量数据(https://www.webmap.cn/main.do?method=index)计算得到;上述数据统一投影至WGS_1984_UTM_Zone_47N坐标系, 并利用最邻近法重采样至30 m。亚洲象活动点数据来源于2020年6月—2021年1月实地调查和国家林业和草原局亚洲象研究中心野外监测, 共计活动点定位记录213个;亚洲象肇事公众责任保险理赔数据来自中国太平洋保险股份有限公司云南分公司(数据使用严格遵守保密协议), 对其空间化后计算亚洲象肇事频次[28]。
2.2 研究方法 2.2.1 生境评价因子基于文献数据库对亚洲象生境适宜性研究相关文献进行检索并统计生境评价因子, 选取出现频率较高的生境评价因子, 采用Pearson相关系数(Pearson Correlation Coefficient)和方差膨胀因子(Variance Inflation Factor, VIF)独立性检验对生境评价因子进行两次筛选, 共获得15个生境评价因子[19]。
2.2.2 生境适宜性模型MaxEnt模型基于最大熵模型原理依据物种活动点和生境评价因子来预测物种的潜在分布[29]。该模型根据物种活动点的环境变量特征计算约束条件, 以此探寻物种最大熵的可能分布, 预测目标物种的适宜生境[30]。模型利用AUC(Area Under Curve)值对预测结果进行精度评价, 数值越大表示预测结果越好。AUC值评价标准为:0.5—0.6不合格, 0.6—0.7较差, 0.7—0.8一般, 0.8—0.9较好, 0.9—1.0非常好[31]。MaxEnt模型在动植物的潜在分布预测中具有良好的预测能力[8, 32], 与其它物种分布模型相比具有较高的可信度[33]。将选取的15个生境评价因子和亚洲象活动点输入MaxEnt模型, 从活动点中随机选择75%用于模型构建, 25%用于模型验证;模型运行次数设置为10次, 模型参数设置参考相关文献[8, 34], 计算结果以Logistic格式输出, 采用AUC值衡量模型预测效果。
由于2020年之前的亚洲象活动点观测数据较少, 故本研究假设研究时段内亚洲象生境偏好不变, 利用MaxEnt模型获取的15个生境评价因子贡献率, 结合HSI模型计算1988—2020年逐年的亚洲象生境适宜性指数[7], 其计算公式为[35]:

|
(1) |
式中, fi为第i个评价因子, wi为第i个因子的权重, n为评价因子数量。
参照已有研究, 结合野外监测的亚洲象活动点位置, 将其与生境适宜性评价结果进行匹配, 依据活动点位置调整分类标准, 将生境适宜性划分为非生境(0—0.58)、边际生境(0.58—0.65)、较适生境(0.65—0.70)、最适生境(0.70—1.0)4个等级[7, 15, 36]。
2.2.3 生境景观格局分析运用FRAGSTATS4.2[19]计算研究区亚洲象生境景观的斑块数(Number of Patches, NP)、斑块密度(Patch Density, PD)、最大斑块指数(Largest Patch Index, LPI)、平均斑块面积(Mean Patch Area, AREA_MN)、斑块分维数(Fractal Dimension Index, FRAC)、斑块连通度指数(Connectance Index, CONNECT)、景观形状指数(Landscape Shape Index, LSI)、Shannon多样性指数(Shannon′s Diversity Index, SHDI)8个景观指数, 以此分析生境景观格局特征及其变化过程。其中, NP表示生境景观中各类斑块的数量;PD表示生境景观的破碎化程度;LPI表示最大生境斑块的面积占比, 取值范围:1 < LPI≤100;AREA_MN表示生境斑块的平均面积大小;FRAC取值范围[1, 2], 值越接近1, 表示生境受人类活动影响越大, 反之越小[7];CONNECT反映生境间的连接程度, 取值范围[0, 1], 数值越小表示生境间连通度越低, 生境破碎化程度越高;LSI反映生境斑块形状的复杂化趋势, 值越大表明生境景观的结构愈复杂, 受人类活动影响越强, 取值范围:LSI≥1, 无上限;SHDI可衡量景观结构的复杂度, 其值越大表明各斑块类型在景观中呈均衡化趋势分布, 景观中的土地利用类型越丰富, 破碎化程度越高, 取值范围:SHDI≥0, 无上限。
2.2.4 Mann-Kendall检验Mann-Kendall检验(简称MK检验)是非参数统计显著性检验, 不需要数据符合特定分布规律, 可用来判断长时间序列数据的变化趋势[37]。计算公式如下:
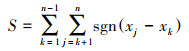
|
(2) |
式中, n为样本总量, j、k=1, 2, …, n; xj、xk分别为第j、k时刻的样本值。sgn为符号函数, 公式如下:

|
(3) |
S为正态分布, 其均值为0, 方差var(S)=n(n-1)(2n+5)/18, 当n>10时, 正态分布统计量计算如下:

|
(4) |
本文检验选取显著水平α=0.05, Z=1.96进行显著性检验, 当|Z|>1.96时表明通过了显著性检验, Z>1.96表明生境质量显著改善, Z < -1.96表明生境质量显著退化, -1.96 < Z < 1.96表明无显著变化[38]。
2.2.5 时序生境与人象冲突的分析地理探测器模型是探测某类现象空间分异并揭示其主要驱动力的方法[39], 可用于研究不同影响因子与目标要素的作用机制。研究应用探测器的因子探测模块探测生境适宜性和景观变化对亚洲象肇事频率的影响, 各因子对亚洲象肇事频率的影响用q值衡量, 公式如下:

|
(5) |
式中, L为自变量X的分层, Nh和σh2为h层内单元数和方差, N和σ为研究区单元数和方差。q取值[0, 1], q值越大表示因子解释力越强, 反之减弱。基于1988—2020年时序生境适宜性指数计算33年生境的最大值、中值、均值等指标, 用以代表历史生境适宜性;计算香农多样性指数、景观形状指数和斑块数量, 并取其中值用以代表历史生境景观状况;将历史生境适宜性指标、历史景观指数中值、现状生境适宜性及现状生境景观指数(2020年生境及景观指数)作为自变量因子, 亚洲象肇事频率作为因变量, 离散化后输入地理探测器, 依据各指标的q值分析生境适宜性和生境景观变化对亚洲象肇事的影响。
3 研究结果 3.1 生境空间格局变化 3.1.1 生境评价因子权重MaxEnt模型运行10次后, AUC平均值为0.841±0.020。基于MaxEnt模型获得各生境评价因子权重, 结果如图 2。

|
| 图 2 生境评价因子权重 Fig. 2 Weights of habitat evaluation factors |
由图 3、图 4可知, 1988—2020年间, 亚洲象最适生境主要分布于研究区东北部、西部及南部;较适生境则呈镶嵌式分布在最适生境外围;边际生境主要分布于较适生境周边, 呈扩张趋势;非生境位于边际生境外围主要分布在研究区中部及北部。亚洲象三类生境中, 边际生境面积最多, 较适生境面积次之, 最适生境面积最少。近30年来, 研究区内边际生境面积显著增加, 较适生境和最适生境面积显著减少;其中, 2000—2013年亚洲象适宜生境大面积减少, 而2013年之后适宜生境变动较小。1988—2020年亚洲象生境由7757.12 km2减少至7373.18 km2, 减少383.94 km2(4.9%)。其中1988—2020年间最适生境面积持续减少, 2020年最适生境面积与1988年相比最适生境减少568.48 km2 (25.4%), 较适生境面积减少187.95 km2(7.3%), 边际生境面积增加372.49 km2(12.6%)。

|
| 图 3 西双版纳勐海—普洱澜沧亚洲象活动区生境分布 Fig. 3 Spatial patterns of Asian elephant habitats in the range of Menghai-Lancang elephant herd |

|
| 图 4 1988—2020年亚洲象生境面积变化和各类生境间转移 Fig. 4 Area changes of Asian elephant habitat and areas conversion among different habitat types from 1988 to 2020 |
由图 5可知, 研究区亚洲象生境类型的NP、PD和FRAC表现为边际生境>较适生境>最适生境, 而CONNECT则是最适生境>较适生境>边际生境。2000—2013年最适生境AREA_MN急剧减小, 同时最适生境FRAC明显升高。与1988年相比, 2020年亚洲象各类生境的LPI、AREA_MN、CONNECT均表现为下降。其中, 最适生境AREA_MN从14.89降至10.05, 降幅达32.5%;最适生境LPI从7.80降至1.64, 降幅达78.97%。

|
| 图 5 西双版纳勐海-普洱澜沧亚洲象活动区生境景观指数变化 Fig. 5 Landscape indices changes of Asian elephant habitat in the range of Menghai-Lancang elephant herd |
图 6表明, 亚洲象肇事区主要位于研究区中部。发展河乡、勐阿镇、勐往乡和勐满镇为亚洲象肇事频发区域, 尤以发展河乡、勐往乡、勐阿镇三个乡镇交界处为甚。研究区生境退化面积占比为9.11%, 生境改善面积占比为2.93%;其中, 亚洲象肇事区生境退化面积占比为10.47%, 生境改善面积占比为1.32%;无肇事区生境退化面积占比为8.11%, 生境改善面积占比为2.89%;亚洲象肇事区生境退化面积占比大于无肇事区。

|
| 图 6 研究区生境退化与改善空间分布和亚洲象肇事区肇事频率 Fig. 6 Spatial pattern of habitat degradation and improvement and frequency of Asian elephant accidents |
除现状生境外, 时序生境、现状与时序生境的景观指数均与亚洲象肇事频率存在显著相关(P < 0.05), 且景观指数的解释力高于生境适宜性的解释力, 尤以时序生境景观指数的解释力最高(图 7);在各景观指数中, 斑块数和景观形状指数对亚洲象肇事频率的解释力较强。

|
| 图 7 时序生境、现状生境、时序景观指数及现状景观指数对亚洲象肇事影响的地理探测器q值 Fig. 7 Geographical detector′s q value between the frequency of Asian elephant accidents and time-series and current habitat suitability and the landscape indices |
(1) 基于有限物种活动点数据的长时序生境制图方法:研究利用MaxEnt模型结合亚洲象活动点计算得到亚洲象生境评价因子权重, 具有明确的生态学意义, 且不依赖主观经验知识;将计算得到的因子权重用于构建HSI模型, 并量化研究区亚洲象的历史生境分布, 突破了物种历史活动点的限制[7, 12], 客观地表征了亚洲象长时序历史生境的变化过程。以往基于生态位模型的生境评价多局限于某单一时间节点, 鲜有长时序生境评价研究;采用HSI模型的生境适宜性评价, 主要通过专家打分法、层次分析法和主成分分析法等确定评价因子权重[19, 40]。层次分析法和专家打分法依赖经验知识来确定因子重要性, 受主观因素影响较大[35];而主成分分析法虽可避免主观因素的影响, 但各因子权重的确定缺乏生态学解释力[41]。本研究基于亚洲象活动点数据, 利用MaxEnt模型定量获取亚洲象的生境偏好, 但未考虑亚洲象生境偏好变化对长时序生境评价的潜在影响。
(2) 勐海—澜沧象群活动区生境时空变化:研究区内亚洲象生境主要沿河流干流分布, 受地形影响明显;研究期内亚洲象适宜生境面积明显减少, 生境斑块破碎化日趋严重, 斑块间连通性显著降低, 这与之前众多学者的研究结果一致[7, 14—15, 42], 而本文首次揭示了该区域亚洲象生境的年尺度变化特征与过程。研究结果表明, 亚洲象生境主要沿河流干流向两岸依次为最适生境、较适生境和边际生境, 该生境格局反映了亚洲象对生境的基本需求, 如河岸的低海拔森林区可提供隐蔽场所[15]、水源、充足食物以及必要的矿物质盐份[42—43]。亚洲象适宜生境大部分位于研究区东北部及西部沿边地区, 因受水电站建成后澜沧江水位上升与海拔阻隔影响[25], 这些生境尚未得到有效利用。目前象群主要活动区内适宜生境少, 且不连续, 导致象群活动持续向外扩张[25, 27]。20世纪80年代初期以来, 土地承包政策的施行, 加之当地经济作物价格的持续升温, 促进了该区域茶叶、橡胶等经济作物种植的扩张[44], 亚洲象适宜生境被人类生产和生活空间所侵占, 使2000—2013年间适宜生境面积呈现锐减;2013年后, 受世界经济环境及生态环境保护政策的综合影响[45—46], 区域内甘蔗等经济作物种植扩张减缓[47], 生境受扰减弱, 亚洲象生境面积也逐渐趋于稳定。研究发现, 1988—2020年间, 亚洲象生境的AREA_MN、CONNECT显著减小, 而NP、PD、FRAC显著升高, 表明亚洲象生境长期遭受人类活动干扰, 生境破碎化现象逐年加重[48]。因此, 研究认为, 为实现该亚洲象群的就地保护, 应加强研究区中部地区适宜生境的修复, 提升斑块间的连通性。
(3) 勐海—澜沧象群活动区时序生境变化与人象冲突的关系:研究发现, 亚洲象肇事主要集中在研究区中部, 且亚洲象肇事区生境退化较无肇事区严重。该象群目前主要在研究区中部活动, 但该区域亚洲象适宜生境少且分布不连续, 客观上加剧了人象冲突, 成为亚洲象肇事的热点地区。相关性分析表明:人象冲突与生境景观格局变化存在显著相关;研究区1988—2020年生境景观破碎加剧, NP显著增大, 最大生境斑块面积由174.72 km2剧减至27.44 km2, 远小于亚洲象最小的家域面积需求(105 km2)[49];人类活动干扰使亚洲象生境斑块被农田、茶园等分割, 生境斑块内地类数量增多, LSI和SHDI变大。亚洲象是一种移栖性动物[42], 生境破碎化切断了亚洲象的天然迁移通道[50], 影响了亚洲象的活动行为, 导致象群频繁穿梭于农田、村寨等地[36]。另外, 农田、茶园等人类生产生活空间与亚洲象所依赖的生态空间交错分布、互相重叠[51], 客观上增加了人象相遇的概率, 增加了人象冲突的风险。生境适宜性对亚洲象的生存、繁衍十分重要。已有研究表明[14, 42], 人类活动及环境变化导致亚洲象生境退化、亚洲象天然食物的减少或出现短缺, 迫使亚洲象取食农作物, 继而引发人象冲突。由此推断, 人类活动与自然环境变化造成的生境退化和生境景观格局变化是影响区域亚洲象肇事频发的主要因素。因此, 亚洲象生境斑块间连通性和破损生境的修复, 对缓解人象冲突具有重要意义。此外, 研究结果表明, 生境丧失和破碎化会使亚洲象不断寻求新的生境, 以维持种群的生存[52—53]。2021年北移象群从勐养子保护区出走, 北迁至昆明城郊活动, 引起了国内外的广泛关注, 有关研究发现北移象群的原生生境适宜性高于新迁入地, 且新迁入地难以支撑象群的长期生存, 而南返则更有利于其生存[19, 54]。从本文的研究来看, 亚洲象北迁可能是亚洲象应对原生生境变化的一种探索行为。
4.2 结论整合MaxEnt与HSI模型, 结合高精度土地利用/覆被时序数据和亚洲象肇事公众责任保险理赔数据, 评价了1988—2020年西双版纳勐海—普洱澜沧亚洲象种群活动区的年尺度生境适宜性, 并分析了亚洲象生境长时序时空变化过程及其与人象冲突的关系, 得出以下主要结论:
(1) 基于物种生境偏好不变的前提, 利用高精度监测获取的物种活动点数据, 融合MaxEnt模型与HSI模型的生境适宜性评价方法可用于物种长时序生境评价。
(2) 西双版纳勐海—普洱澜沧亚洲象种群活动区亚洲象适宜生境主要沿河流干流分布, 按距干流由近到远依次为最适生境、较适生境、边际生境。
(3) 研究期内, 因受橡胶和茶叶等经济作物种植的影响, 亚洲象适宜生境显著减少, 尤以2000—2013年最为严重;同时斑块间连通性显著降低, 生境破碎加重。
(4) 研究区人象冲突主要集中于生境退化区, 人类活动与自然环境变化导致的生境退化和生境景观格局变化是影响亚洲象肇事的重要因素。
| [1] |
Chase J M, Blowes S A, Knight T M, Gerstner K, May F. Ecosystem decay exacerbates biodiversity loss with habitat loss. Nature, 2020, 584(7820): 238-243. DOI:10.1038/s41586-020-2531-2 |
| [2] |
Powers R P, Jetz W. Global habitat loss and extinction risk of terrestrial vertebrates under future land-use-change scenarios. Nature Climate Change, 2019, 9(4): 323-329. DOI:10.1038/s41558-019-0406-z |
| [3] |
Richards D R, Thompson B S, Wijedasa L. Quantifying net loss of global mangrove carbon stocks from 20 years of land cover change. Nature Communications, 2020, 11(1): 4260. DOI:10.1038/s41467-020-18118-z |
| [4] |
颜忠诚, 陈永林. 动物的生境选择. 生态学杂志, 1998, 17(2): 43-49. DOI:10.3321/j.issn:1000-4890.1998.02.009 |
| [5] |
Rattis L, Dobrovolski R, Talebi M, Loyola R. Geographic range-scale assessment of species conservation status: a framework linking species and landscape features. Perspectives in Ecology and Conservation, 2018, 16(2): 97-104. DOI:10.1016/j.pecon.2018.01.001 |
| [6] |
郭彦龙, 赵泽芳, 乔慧捷, 王然, 卫海燕, 王璐坤, 顾蔚, 李新. 物种分布模型面临的挑战与发展趋势. 地球科学进展, 2020, 35(12): 1292-1305. DOI:10.11867/j.issn.1001-8166.2020.110 |
| [7] |
吴得卿, 魏建华, 樊辉. 南滚河流域土地利用变化对亚洲象生境的影响. 生态学报, 2021, 41(16): 6476-6485. |
| [8] |
唐书培, 穆丽光, 王晓玲, 张静, 刘波, 孟和达来, 鲍伟东. 基于MaxEnt模型的赛罕乌拉国家级自然保护区斑羚生境适宜性评价. 北京林业大学学报, 2019, 41(1): 102-108. |
| [9] |
Tantipisanuh N, Gale G A, Pollino C. Bayesian networks for habitat suitability modeling: a potential tool for conservation planning with scarce resources. Ecological Applications, 2014, 24(7): 1705-1718. DOI:10.1890/13-1882.1 |
| [10] |
Schaefer H C, Jetz W, Böhning-Gaese K. Impact of climate change on migratory birds: community reassembly versus adaptation. Global Ecology and Biogeography, 2008, 17(1): 38-49. |
| [11] |
Richmond O M W, McEntee J P, Hijmans R J, Brashares J S. Is the climate right for pleistocene rewilding? Using species distribution models to extrapolate climatic suitability for mammals across continents. PLoS One, 2010, 5(9): e12899. DOI:10.1371/journal.pone.0012899 |
| [12] |
廖颖, 王心源, 周俊明. 基于地理探测器的大熊猫生境适宜度评价模型及验证. 地球信息科学学报, 2016, 18(6): 767-778. |
| [13] |
Roloff G J, Kernohan B J. Evaluating reliability of habitat suitability index models. Wildlife Society Bulletin, 1999, 27(4): 973-985. |
| [14] |
林柳, 金延飞, 陈德坤, 郭贤明, 罗爱东, 赵建伟, 王巧燕, 张立. 西双版纳国家级自然保护区勐腊子保护区亚洲象种群和栖息地评价. 生态学报, 2014, 34(7): 1725-1735. |
| [15] |
刘鹏, 代娟, 曹大藩, 李志宏, 张立. 普洱市亚洲象栖息地适宜度评价. 生态学报, 2016, 36(13): 4163-4170. |
| [16] |
Harrity E J, Stevens B S, Conway C J. Keeping up with the times: mapping range-wide habitat suitability for endangered species in a changing environment. Biological Conservation, 2020, 250: 108734. DOI:10.1016/j.biocon.2020.108734 |
| [17] |
Store R, Kangas J. Integrating spatial multi-criteria evaluation and expert knowledge for GIS-based habitat suitability modelling. Landscape and Urban Planning, 2001, 55(2): 79-93. DOI:10.1016/S0169-2046(01)00120-7 |
| [18] |
Vaughan I P, Ormerod S J. Improving the quality of distribution models for conservation by addressing shortcomings in the field collection of training data. Conservation Biology, 2003, 17(6): 1601-1611. DOI:10.1111/j.1523-1739.2003.00359.x |
| [19] |
张鑫, 尹文萍, 谢菲, 樊辉, 陈飞. 元江-李仙江流域亚洲象生境适宜性评价——基于荟萃分析和遥感大数据分析. 生态学报, 2022, 42(12): 5067-5078. |
| [20] |
Campos-Arceiz A, Blake S. Megagardeners of the forest-the role of elephants in seed dispersal. Acta Oecologica, 2011, 37(6): 542-553. DOI:10.1016/j.actao.2011.01.014 |
| [21] |
De R, Sharma R, Davidar P, Arumugam N, Sedhupathy A, Puyravaud J P, Selvan K M, Rahim P P A, Udayraj S, Parida J, Digal D K, Kanagaraj R, Kakati K, Nigam P, Williams A C, Habib B, Goyal S P. Pan-India population genetics signifies the importance of habitat connectivity for wild Asian elephant conservation. Global Ecology and Conservation, 2021, 32: e01888. DOI:10.1016/j.gecco.2021.e01888 |
| [22] |
林柳, 张立. 象科动物对当地森林生态系统的影响. 兽类学报, 2018, 38(4): 411-419. |
| [23] |
赵宇, 金崑. 亚洲象分布、数量、栖息地状况及种群管理. 世界林业研究, 2018, 31(2): 25-30. |
| [24] |
Wang Z H, Li Z L, Tang Y J, Yao C X, Liu Y, Jiang G L, Wang F, Liang L, Zhao W L, Zhu G F, Chen M Y. China's dams isolate Asian elephants. Science, 2020, 367(6476): 373-374. |
| [25] |
王智红, 张恒振, 赵文蓝, 郑璇, 岩温扁, 李正玲, 陈明勇. 西双版纳勐海-普洱澜沧亚洲象种群动态分析. 林业建设, 2019(6): 79-84. |
| [26] |
Liu S L, Yin Y J, Li J R, Cheng F Y, Dong S K, Zhang Y Q. Using cross-scale landscape connectivity indices to identify key habitat resource patches for Asian elephants in Xishuangbanna, China. Landscape and Urban Planning, 2018, 171: 80-87. |
| [27] |
王巧燕, 陶永祥, 李劲松, 董忠, 岩丙, 杨鸿培. 西双版纳地区野生亚洲象种群分布及变迁原因分析. 林业调查规划, 2017, 42(4): 113-118. |
| [28] |
尹文萍, 高宸, 樊辉, 谢菲, 张鑫. 一种融合文本中地理位置和土地利用/覆被信息的野生动物活动细粒度定位方法. 地球信息科学学报, 2022, 24(7): 1363-1374. |
| [29] |
Pearson R G, Raxworthy C J, Nakamura M, Peterson A T. ORIGINAL ARTICLE: predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. Journal of Biogeography, 2007, 34(1): 102-117. |
| [30] |
Phillips S J, Dudik M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 2008, 31(2): 161-175. |
| [31] |
Swets J A. Measuring the accuracy of diagnostic systems. Science, 1988, 240(4857): 1285-1293. |
| [32] |
刘振生, 高惠, 滕丽微, 苏云, 王晓勤, 孔芳毅. 基于MAXENT模型的贺兰山岩羊生境适宜性评价. 生态学报, 2013, 33(22): 7243-7249. |
| [33] |
王运生, 谢丙炎, 万方浩, 肖启明, 戴良英. ROC曲线分析在评价入侵物种分布模型中的应用. 生物多样性, 2007, 15(4): 365-372. |
| [34] |
Phillips S J, Anderson R P, Schapire R E. Maximum entropy modeling of species geographic distributions. Ecological Modelling, 2006, 190(3): 231-259. |
| [35] |
Nyimbili P H, Erden T. A hybrid approach integrating entropy-AHP and GIS for suitability assessment of urban emergency facilities. ISPRS International Journal of Geo-Information, 2020, 9(7): 419. |
| [36] |
林柳, 金延飞, 杨鸿培, 罗爱东, 郭贤明, 王利繁, 张立. 西双版纳亚洲象的栖息地评价. 兽类学报, 2015, 35(1): 1-13. |
| [37] |
Gocic M, Trajkovic S. Analysis of changes in meteorological variables using Mann-Kendall and Sen's slope estimator statistical tests in Serbia. Global and Planetary Change, 2013, 100: 172-182. |
| [38] |
Liu Y, Li L H, Chen X, Zhang R, Yang J M. Temporal-spatial variations and influencing factors of vegetation cover in Xinjiang from 1982 to 2013 based on GIMMS-NDVI3g. Global and Planetary Change, 2018, 169: 145-155. |
| [39] |
王劲峰, 徐成东. 地理探测器: 原理与展望. 地理学报, 2017, 72(1): 116-134. |
| [40] |
林玲, 王卫红, 冉茂莹, 耿诗画, 杨元继. 王朗自然保护区大熊猫生境适宜性及潜在活动区域. 东北林业大学学报, 2022, 50(1): 87-92. |
| [41] |
Hirzel A H, Le Lay G. Habitat suitability modelling and niche theory. Journal of Applied Ecology, 2008, 45(5): 1372-1381. |
| [42] |
张立, 王宁, 王宇宁, 马利超. 云南思茅亚洲象对栖息地的选择与利用. 兽类学报, 2003, 23(3): 185-192. |
| [43] |
Neupane D, Kwon Y, Risch T S, Williams A C, Johnson R L. Habitat use by Asian elephants: context matters. Global Ecology and Conservation, 2019, 17: e00570. |
| [44] |
Su S L, Zhou X C, Wan C, Li Y K, Kong W H. Land use changes to cash crop plantations: crop types, multilevel determinants and policy implications. Land Use Policy, 2016, 50: 379-389. |
| [45] |
Li Y C, Liu C L, Zhang J, Zhang P, Xue Y F. Monitoring spatial and temporal patterns of rubber plantation dynamics using time-series landsat images and google earth engine. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing, 2021, 14: 9450-9461. |
| [46] |
中华人民共和国环境保护部. 全国生态保护"十二五"规划. (2013-01-25)[2022-01-21]. http://www.gov.cn/gongbao/content/2013/content_2396624.htm.
|
| [47] |
勐海县人民政府. 勐海县"十三五"蔗糖产业发展规划. (2018-03-06)[2022-01-28]. https://www.ynmh.gov.cn/cjj/21785.news.detail.dhtml?news_id=110807.
|
| [48] |
An Y, Liu S L, Sun Y X, Shi F N, Zhao S. Negative effects of farmland expansion on multi-species landscape connectivity in a tropical region in Southwest China. Agricultural Systems, 2020, 179: 102766. |
| [49] |
Jathanna D, Karanth K U, Kumar N S, Karanth K K, Goswami V R. Patterns and determinants of habitat occupancy by the Asian elephant in the western ghats of karnataka, India. PLoS One, 2015, 10(7): e0133233. |
| [50] |
Li W W, Liu P, Guo X M, Wang L X, Wang Q Y, Yu Y, Dai Y C, Li L, Zhang L. Human-elephant conflict in Xishuangbanna Prefecture, China: distribution, diffusion, and mitigation. Global Ecology and Conservation, 2018, 16: e00462. |
| [51] |
Chan A N, Wittemyer G, McEvoy J, Williams A C, Cox N, Soe P, Grindley M, Shwe N M, Chit A M, Oo Z M, Leimgruber P. Landscape characteristics influence ranging behavior of Asian elephants at the human-wildlands interface in Myanmar. Movement Ecology, 2022, 10(1): 6. |
| [52] |
Ram A K, Yadav N K, Kandel P N, Mondol S, Pandav B, Natarajan L, Subedi N, Naha D, Reddy C S, Lamichhane B R. Tracking forest loss and fragmentation between 1930 and 2020 in Asian elephant (Elephas maximus) range in Nepal. Scientific Reports, 2021, 11(1): 19514. |
| [53] |
Wilson S, Davies T E, Hazarika N, Zimmermann A. Understanding spatial and temporal patterns of human-elephant conflict in Assam, India. Oryx, 2015, 49(1): 140-149. |
| [54] |
陈飞, 杨子诚, 杨聪, 赵明旭, 徐卫平, 汤永晶, 王智红, 张光元. 云南亚洲象北移区域生境适宜性评价. 自然保护地, 2021, 1(3): 9-15. |