 2023, Vol. 43
2023, Vol. 43文章信息
- 徐雪, 罗娅, 杨胜天, 陆晓辉, 周秋文, 刘茂, 石春茂, 廖梦垚
- XU Xue, LUO Ya, YANG Shengtian, LU Xiaohui, ZHOU Qiuwen, LIU Mao, SHI Chunmao, LIAO Mengyao
- 全球不同气候带陆地植被净初级生产力变化趋势与可持续性
- Change trend and sustainability of vegetation net primary productivity of terrestrial ecosystems in different global climatic zones
- 生态学报. 2023, 43(9): 3729-3743
- Acta Ecologica Sinica. 2023, 43(9): 3729-3743
- http://dx.doi.org/10.5846/stxb202205061255
-
文章历史
- 收稿日期: 2022-05-06
- 网络出版日期: 2022-09-05






2. 贵州省喀斯特山地生态环境国家重点实验室培育基地, 贵阳 550025;
3. 北京师范大学水科学研究院, 北京 100875
2. State Key Laboratory Incubation Base for Karst Mountain Ecology Environment of Guizhou Province, Guiyang 550025, China;
3. College of Water Sciences, Beijing Normal University, Beijing 100875, China
植被净初级生产力(NPP)作为陆地生态系统能量和质量转换的关键组成部分[1], 可有效量化陆地生态系统的生产能力[2], 是评估碳汇水平的重要指标[3]。宏观上, 气候是决定植被类型和空间分布的最主要的非生物因素[4—5]。不同的气候带具有不同的水热格局, 影响着植被的结构、功能和碳吸收能力[6]。对比研究全球不同气候带陆地植被NPP变化趋势与可持续性, 对于估算全球陆地生态系统的结构、功能和碳源(汇)具有十分重要的意义。
已有成果常用长时序列原位监测法[7]、模型模拟法[8—9]等获取NPP数据后揭示不同气候带的陆地NPP变化趋势。研究表明, 全球NPP在1981—2018年总体呈显著上升趋势, 但在2000—2009年有所减少[10]。热带地区, 亚洲和亚马逊热带雨林的平均NPP显著下降, 非洲热带雨林的平均NPP显著上升[11];泰国东部的热带季风区的红树林NPP没有明显变化[7]。中国的亚热带地区, 植被NPP呈显著上升趋势[12—13]。中国内蒙古温带荒漠草原区, 植被NPP有下降趋[14]。干旱的中亚五国, 其NPP有轻微上升趋势[15]。澳大利亚半干旱草地, 其NPP呈退化状态[16]。高寒的青藏高原区, 植被NPP呈显著增加趋势, 增加速率为1.67gCm-2a-1[17]。这些成果较好地揭示了不同气候区的NPP变化趋势。但是, 由于它们所研究的时空尺度不一致, 精度不一致, 可靠性不一致, 因此应用这些成果开展全球不同气候带陆地植被NPP变化趋势对比分析时面临一定的挑战, 不同气候带陆地生态系统的碳源(汇)功能还存在争议。
卫星遥感作为全球生态系统研究的重要手段, 可获得时空更连续、精度更高的NPP数据产品[18], 对于研究全球NPP变化具有较好的优势[19—21]。因此, 基于遥感方法获取全球陆地NPP数据, 运用Mann-Kendall趋势检验、Theil-Sen斜率估计和Hurst指数, 研究2001—2018年全球不同气候带陆地NPP变化趋势与可持续性, 主要解决三个问题:(1)阐明2001—2018年全球陆地平均NPP空间分布及变化趋势;(2)揭示不同气候带NPP变化趋势及突变点;(3)揭示不同气候带NPP变化的可持续性。希望通过研究, 为国内外同行对比分析全球不同气候带陆地生态系统的碳源(汇)功能提供参考。
1 数据与方法 1.1 研究方法(1) Mann-Kendall突变分析
运用Mann-Kendall突变检验分析法判断全球NPP变化的突变点[22—23], 计算公式如下:
第一, 对2001—2018年NPP均值构造一个秩序列为:
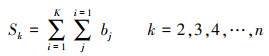
|
(1) |
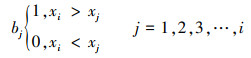
|
(2) |
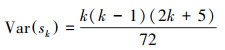
|
(3) |
第二, 定义统计变量:
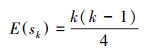
|
(4) |
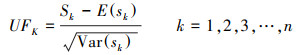
|
(5) |
式中, Sk为秩序列, k为时间序列长度, xi、xj分别为第i年和第j年的NPP值, Var(sk) 为Sk的方差, E(sk)为Sk的均值, UFK是标准正态分布。将时间序列x按照逆序列排序, 重复上述公式, 可计算出另一条UBK曲线。最后在置信水平α=0.05和两条曲线的临界值±1.96下进行解读。若UF>0, 则代表上升趋势;若UF>1.96或UF<-1.96, 代表增加或下降趋势显著;若UFK和UBK两条曲线相交, 并且交点位于临界值内, 该对应交点即为时间序列发生突变的点。
(2) Mann-Kendall趋势检验+Theil-Sen斜率估计
Mann-Kendall趋势检验+Theil-Sen斜率估计适合处理和分析长时间序列或有噪声序列数据, 被广泛应用于水文气象研究[24—25]。鉴于此, 运用该方法分析全球不同气候带的陆地植被NPP变化趋势。具体方法为:
斜率估计:
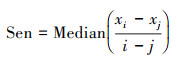
|
(6) |
式中, xi、xj分别为第i年和第j年的NPP值, Median表示所求时间序列的中位数。Sen估计值可以是+1、0或-1, 分别代表增加趋势、稳定不变、减少趋势。之后, 运用Mann-Kendall趋势检验分析法进一步判断Sen值计算结果的显著性, 公式如下:
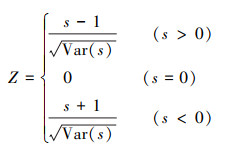
|
(7) |
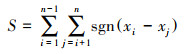
|
(8) |
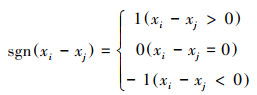
|
(9) |
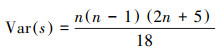
|
(10) |
式中, S为统计变量, xi、xj分别为第i年和第j年的NPP值, n为样本数量, sgn为符号函数。当|Z| >1.96时, NPP的变化趋势显著, 反之则为不显著。根据前人研究[26], 将NPP变化趋势划分为五类, 即:严重退化(Sen < -0.0005, Z≥1.96或Z≤-1.96)、轻微退化(Sen < -0.0005, -1.96 < Z < 1.96)、稳定不变(-0.0005 < Sen < 0.0005)、轻微改善(Sen>0.0005, -1.96 < Z < 1.96)、明显改善(Sen>0.0005, Z≥1.96或Z≤-1.96)。
(3) Hurst指数分析
Hurst指数最初由英国水文学家H.E.Hurst提出, 用于研究洪水形成周期。后来被广泛用于预测时间序列的未来趋势, 尤其在经济学、气候学等领域的运用更为常见[27]。目前, Hurst指数的估算方法有绝对值法、聚合方差法、周期图法、小波分析法、残差分析法和R/S分析法等[28]。其中, 重标极差R/S分析常被用于研究植被变化趋势, 并取得了良好的效果[29—30]。因此, 运用重标极差R/S分析法预测NPP变化的可持续性。方法为:
第一, 定义时间序列x(t):
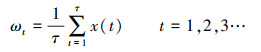
|
(11) |
第二, 计算累计偏差:
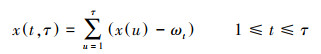
|
(12) |
第三, 计算极差序列:
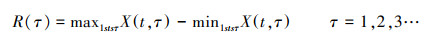
|
(13) |
第四, 计算标准差序列:
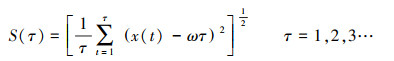
|
(14) |
第五, 取R(τ)和S(τ)之比:
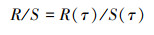
|
(15) |
若:
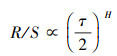
|
(16) |
则说明时间序列存在Hurst现象, 此时H被称为Hurst指数, 其范围为0—1。当H=0.5时, 数据无变化。当0.5 < H < 1时, 该过程具有连续特征, 说明未来趋势与过去一致。当0 < H < 0.5, 表明预计未来趋势将与过去趋势相反[31]。
最后, 根据Theil-Sen斜率估计和Hurst指数分析结果, 分五种情形揭示全球及不同气候带的NPP变化可持续性。即:持续增加(Sen>0, 0.5 < H < 1)、由增加变减少(Sen>0, 0 < H < 0.5)、持续减少(Sen < 0, 0.5 < H < 1)、由减少变为增加(Sen < 0, 0 < H < 0.5)、无法预测(H=0.5)。
1.2 数据来源(1) 植被净初级生产力(NPP)
研究所需的NPP数据来源于MuSyQ NPP产品(http://doi.org/10.5281/Zenodo.3996814), 该数据空间分辨率为0.05°, 时间分辨率为8d, 对估算全球NPP有较好的可靠性[11]。在分析之前, 将NPP的日数据用最大值合成法(MVC)[32]合成月数据集, 并利用均值法[33]将逐月数据合成逐年数据。
(2) 柯本气候类型数据
气候类型数据来自普林斯顿大学2018年发布的柯本气候分类数据集[34](http://www.gloh2o.org/koppen/)。依据气温和降水两个指标, 该数据集将全球共划为了五个气候带, 即:A(热带)、B(干旱气候带)、C(亚热带)、D(温带、寒温带)、E(极地带)(图 1)。为便于分析, 将气候类型数据采用双线性内插的方法, 重采样到和NPP数据一样的空间分辨率, 并将其坐标与投影方式与NPP数据相统一。

|
| 图 1 全球气候带的空间分布 Fig. 1 Spatial distribution of global climate zones |
如图 2所示, 2001—2018年间, 全球植被NPP年均总量为127.13gCm-2a-1, 呈现出明显的空间分异特征。其高值区集中分布在低纬度气候带, 以南美亚马逊流域、非洲刚果盆地及东南亚岛屿地区为代表;相较于高值区, 全球植被NPP低值区的分布范围明显较广, 具体分布在北美西南部和高纬地区、南美西部、非洲北部和南部、中亚和北亚高纬地区、澳大利亚的荒漠地区。全球不同气候带年均NPP值由高至低依次为:热带(307.70gCm-2a-1)>亚热带(206.95gCm-2a-1)>温带寒温带(123.19gCm-2a-1)>干旱气候带(58.17gCm-2a-1)>极地带(11.05gCm-2a-1)。

|
| 图 2 2001—2018年全球植被NPP的空间分布 Fig. 2 Spatial distribution of global vegetation NPP from 2001 to 2018 |
获取全球陆地NPP各年均值后, 运用线性拟合方法, 可看出全球陆地NPP的变化趋势。如图 3所示, 2001—2018年, 全球陆地NPP呈现波动上升趋势(R2=0.083, P=0.247), 波动范围为125.09—128.56gCm-2a-1, 线性增长速率为0.05gCm-2a-1, 其最大值出现在2001年, 为128.56gCm-2a-1, 最小值出现在2008年, 为125.09gCm-2a-1。运用Mann-Kendall突变分析, 检验全球陆地NPP的突变点。根据突变检验结果, UF和UB两线在2015年相交, 且交点在置信区间内, 表明2015年NPP变化趋势发生突变。

|
| 图 3 2001—2018年全球植被NPP变化趋势及突变检验 Fig. 3 Global vegetation NPP trend and mutation test from 2001 to 2018 |
综合Mann-Kendall趋势检验和Theil-Sen斜率估计结果, 可推测全球陆地NPP的变化类型。由图 4和表 1可看出, NPP显著改善的面积占全球陆地面积的8.45%, 主要分布在北美洲中部平原、亚马逊河流域西部、非洲刚果盆地、欧洲南部、印度半岛西北部、中国黄土高原。NPP轻微改善的面积最大, 占全球陆地面积的40.34%, 主要分布在北美洲北部、南美洲中南部、亚洲东部和澳大利亚大陆东部。NPP稳定不变的面积占全球陆地面积的2.6%, 零星分布于各地。NPP严重退化的面积占全球陆地面积的8.54%, 主要分布在亚马逊河流域东北部、非洲萨赫勒以及加里曼丹岛北部等。其他40.07%的区域则表现为轻微退化。

|
| 图 4 2001—2018年全球陆地NPP变化类型空间分布 Fig. 4 Spatial distribution of global terrestrial NPP change types from 2001 to 2018 |
| 变化趋势 Trends |
像元数 Number of pixels |
像元占比 Percentage of pixels/% |
变化趋势 Trends |
像元数 Number of pixels |
像元占比 Percentage of pixels/% |
|
| 明显改善Significant improvement | 456885 | 8.45 | 轻微退化Slight degradation | 2168007 | 40.07 | |
| 轻微改善Slight improvement | 2182688 | 40.34 | 严重退化Significant degradation | 462105 | 8.54 | |
| 稳定不变Stability | 140470 | 2.60 | ||||
运用线性趋势拟合方法, 分析全球五个气候带NPP在2001—2018年间的变化趋势。在此基础上, 运用Mann-Kendall趋势检验方法, 揭示各气候带NPP的突变年份。研究结果如下:
2.2.1 热带植被NPP变化趋势及突变点如图 5可见, 热带NPP呈不显著的下降趋势(R2=0.111, P=0.176), 下降速率约为0.189gCm-2a-1, NPP值波动范围为313.72—301.48gCm-2a-1。其最高值出现在2001年, 最低值出现在2016年。MK突变检验显示, 热带NPP的突变时间分别为2002年、2012年、2013年。根据滑动t检验进一步验证, 研究期间热带NPP并未存在显著突变年。

|
| 图 5 2001—2018年热带NPP变化趋势及突变检验 Fig. 5 rend and mutation test of NPP in tropical from 2001 to 2018 |
如图 6所示, 干旱气候带NPP呈不显著上升趋势(R2=0.036, P=0.450), 上升速率为0.046gCm-2a-1, NPP值波动范围为55.82—59.96gCm-2a-1。最大值出现2018年, 最小值出现在2002年。MK突变检验显示, 干旱气候带NPP在2004年、2009年、2011年发生突变, 根据滑动t检验进一步验证, 干旱气候带NPP的实际突变年为2009年。

|
| 图 6 2001—2018年干旱气候带NPP变化趋势及突变检验 Fig. 6 Trend and mutation test of NPP in arid climate zones from 2001 to 2018 |
如图 7所示, 亚热带NPP呈现不显著的下降趋势(R2=0.144, P=0.120), 下降速率为0.175gC m-2 a-1, NPP值的波动范围为212.90—202.63gC m-2 a-1。最大值出现在2004年, 最低值出现2018年。MK突变检验结果显示, 亚热带NPP分别在2002年、2004年、2005年、2015年和2017年发生突变。根据滑动t检验结果验证, 热带NPP实际突变年为2015年。

|
| 图 7 2001—2018年亚热带NPP变化趋势及突变检验 Fig. 7 Trend and mutation test of NPP In subtropical from 2001 to 2018 |
由图 8可见, 温带寒温带植被NPP呈显著的上升趋势(R2=0.533, P=0.001), 上升速率为0.297gCm-2a-1, NPP值的波动范围为:119.71—127.53gCm-2a-1。最大值出现在2016年, 最小值出现在2008年。根据MK突变检验结果, 温带寒温带NPP的突变年为2014年。

|
| 图 8 2001—2018年温带寒温带NPP变化趋势及突变检验 Fig. 8 Trend and mutation test of NPP in the temperate cold temperate zone from 2001 to 2018 |
如图 9所示, 极地带NPP变化整体呈不显著的下降趋势(R2=0.002, P=0.854), 下降速率为0.005gCm-2a-1, NPP值的波动范围为12.07—10.11gCm-2a-1。最大值出现在2010年, 最小值出现在2004年、2014年。MK突变检验结果显示, 极地带NPP的突变年为2005年。

|
| 图 9 2001—2018年极地带NPP变化趋势及突变检验 Fig. 9 Trend and mutation test of NPP in polar zones from 2001 to 2018 |
估算2001—2018年各气候带NPP分别与年降水量、年均气温的相关性, 解析不同气候带NPP变化与气候因子的关系。
2.3.1 不同气候带NPP变化与年降水量变化的相关性降水对NPP的作用在不同气候区会呈现差异。由图 10可见, 热带NPP变化与年降水量无明显相关性(R2=0.001, P=0.894), 干旱气候带NPP变化与年降水量存在显著的正相关关系(R2=0.269, P=0.027), 亚热带NPP变化与年降水量无明显相关性(R2=0.011, P=0.680), 温带寒温带NPP变化与年降水量存在不显著的正相关关系(R2=0.133, P=0.137), 极地带NPP变化与年降水量存在较显著的负相关关系(R2=0.257, P=0.032)。结果表明, 热带、亚热带和温带寒温带NPP受年降水量影响不显著。在干旱气候带, 年降水量增加可能会提高NPP和生态系统碳吸存。而在极地带, 由于其降水多为较寒冷的固态水, 固态水会对植被生产力产生抑制作用, 因此年降水量增加会显著降低NPP和生态系统碳吸存[35]。

|
| 图 10 不同气候带NPP变化与年降水量的相关性 Fig. 10 Correlation between NPP variation and annual precipitation in different climatic zones |
气温也影响着植被生长, 全球植被生产力的最佳适宜温度为(23±6)℃[36]。不同气候带的气温具有显著差异性, 因而其NPP的变化特征也各不相同。如图 11所示, 热带NPP变化与年均气温有显著的负相关性(R2=0.487, P=0.001), 干旱气候带NPP变化与年均气温无明显相关性(R2=0.036, P=0.449), 亚热带NPP变化与年均气温无明显相关性(R2=0.00002, P=0.985), 温带寒温带NPP变化与年均气温有显著的正相关性(R2=0.585, P < 0.001), 极地带NPP变化与年均气温有显著的正相关性(R2=0.419, P=0.04)。结果表明, 在热带, 气温升高抑制了NPP和生态系统碳吸收。干旱气候带和亚热带NPP变化受气温影响不明显。温带寒温带和极地带上升的气温大大提高了植被的固碳能力。

|
| 图 11 不同气候带NPP变化与年均气温变化的相关性 Fig. 11 Correlation of NPP changes with annual mean temperature changes in different climatic zones |
揭示全球及不同气候带NPP变化的可持续性, 有助于了解植被未来的发展动态, 从而为制定有效的应对策略提供参考。
2.4.1 全球NPP未来变化趋势的空间分布如图 12所示, 全球陆地生态系统的未来演进中, NPP呈减少的面积(51.31%)大于增加的面积(48.69%)。其中, NPP持续减少的区域主要分布在北美洲西南部、亚马逊河流域中部和东部、南美洲南部、非洲北部、亚洲西部、加里曼丹岛北部、新几内亚岛东南部、澳大利亚大陆中部等地区, 面积占比为22.02%;NPP由增变减的区域主要分布在北美洲中部和北部、亚洲北部、亚马逊河流域西部、非洲萨赫勒等地区, 面积占比为29.29%。总体上, 未来全球陆地生态系统NPP趋于退化, NPP变化以反持续性特征为主, 占比为58.45%。

|
| 图 12 2001—2018年全球陆地NPP变化可持续性的空间分布 Fig. 12 Spatial distribution of sustainability of global terrestrial NPP change from 2001 to 2018 |
图 13显示各气候带未来的NPP变化可持续性分类情形。可看出:(1)热带、极地带NPP趋于稳定, 热带、极地带NPP增加面积(分别为49.50%、48.80%)与减少面积(分别为50.1%、49.4%)基本相当。(2)干旱气候带、亚热带、温带寒温带NPP趋于退化, 干旱气候带、亚热带、温带寒温带NPP增加面积(分别为43.40%、46.40%、43.30%)明显小于减少面积(分别为46.5%、53.3%、56.3%)。结果表明, 干旱气候带、亚热带、温带寒温带三类地区是未来防止NPP退化工作的重点区域。

|
| 图 13 不同气候带NPP变化的可持续性统计 Fig. 13 Sustainability statistics of NPP changes in different climate zones |
不同气候带降水、气温有显著的差异, 植被NPP受降水、气温的作用机制不同。具体来看, 干旱气候带、极地带NPP主要为水分限制型, 而热带、温带寒温带NPP为热量限制型, 亚热带NPP受气温、降水影响不显著。干旱气候带受水分的正限制作用, 表现为植被对降水变化的敏感性大于温度[37], 随降水增加, 干旱得到缓解, 对植被生长越有利。极地带由于环境潮湿、气温极低, 过多降水加剧植被霜冻灾害的同时[38], 还不利于光合作用[39], 植被生长受到极大抑制, 因此该地区植被表现出受水分的负限制影响, 即随降水增加, NPP下降。对热带地区而言, 其平均气温足够高, 气温上升则会增加呼吸支出[40], 导致NPP降低。与之相反, 在温带寒温带地区, 气温变暖对植被具有显著的积极作用[41], 表现为植被光合作用增加, 植被固碳能力增强[42], 进而使得NPP上升。
3.1.2 降水、气温对NPP的组合影响降水和气温变化并非分别发生, 两者的综合还共同决定了一个地区的冷、暖、干、湿变化[43]。冷暖干湿通过改变植被的物候[44]、代谢[45]和功能性状[46], 进而直接影响陆地生态系统的植被生产力。并且不同水热组合类型对植被NPP的影响是不一样的, 主要表现为冷湿地区的NPP呈显著增加, 暖湿地区的NPP呈显著减少, 其他区域的NPP则无显著变化[47]。可见, NPP除了受降水、气温的单独影响外, 还受两者间协同作用的影响, 并且只有当降水和气温趋于平衡时, 对植被的生长才更有利。
3.2 研究成果适用边界条件及未来研究方向 3.2.1 研究成果适用边界本研究仅从宏观尺度上分析了陆地植被NPP的动态变化, 揭示了各气候带植被NPP的变化特征, 并阐明NPP变化与年均气温、年降水量的相关性, 研究结果为国内外同行对比分析全球不同气候带陆地生态系统的碳源(汇)功能提供了一定参考。然而, 除了气候因素的影响外, 植被NPP的生长还与人类活动[48—49]、土壤[50]、地形[51]、景观格局[52]等因素有关, 这些环境特征在不同区域情况不一, 因此, 研究结果仅代表宏观尺度的情况, 不能反映微观尺度上的具体变化情况。
3.2.2 未来研究方向不同气候带内部存有显著差异, 又分为不同气候类型, 不同气候类型的水热特征各不相同, 进而导致陆地植被NPP的变化及可持续性也会有差异性。而且, 植被自身由于物种组成、群落结构、自然演替规律等存有较大差异[53], 其对外界环境的响应和敏感性也均不相同。因此, 未来工作中将继续开展不同气候类型以及不同植被NPP变化差异的研究, 以期为丰富研究结论和相关应对策略的制定提供重要支撑。
此外, 本研究分析NPP变化的可持续性是基于现阶段植被变化情况预测得到, 随时间的推移, 陆地植被NPP可能会呈现不同的变化趋势。因此, 要准确掌握未来植被的动态变化情况需要不间断地进行监测。
4 结论全球陆地NPP有明显的地域分异规律, 呈现低纬高、高纬低, 沿海高、内陆低的分布特点, 并以每年0.05gC/m2的增速波动上升。各气候带的分布差异表现为热带(307.70gCm-2a-1)>亚热带(206.95gCm-2a-1)>温带寒温带(123.19gCm-2a-1)>干旱气候带(58.17gCm-2a-1)>极地带(11.05gCm-2a-1)。研究期间, NPP变化类型以轻微改善为主, 占全球陆地表面的40.34%, 分布在北美洲北部、南美洲中南部、亚洲东部和澳大利亚大陆东部。
不同气候带NPP变化趋势表现为热带总体表现为不显著下降趋势, 无明显突变年份;干旱气候带整体呈不显著上升趋势, 突变点发生在2009年;亚热带整体呈不显著的下降趋势, 突变点发生在2015年;温带寒温带整体呈显著上升趋势, 突变点发生在2014年;极地带整体呈不明显下降趋势, 突变点发生在2005年。
各气候带NPP与降水、气温的关系为:干旱气候带、极地带NPP主要为水分限制型, 而热带、温带寒温带NPP为热量限制型, 亚热带NPP受气温、降水影响不显著。
在未来, 热带、极地带的NPP趋于稳定, 干旱气候带、亚热带、温带寒温带的NPP趋于退化, 干旱气候带、亚热带、温带寒温带三类地区将是防止NPP退化工作的重点区域。
| [1] |
戴尔阜, 黄宇, 吴卓, 赵东升. 内蒙古草地生态系统碳源/汇时空格局及其与气候因子的关系. 地理学报, 2016, 71(1): 21-34. |
| [2] |
Wolf S, Keenan T F, Fisher J B, Baldocchi D D, Desai A R, Richardson A D, Scott R L, Law B E, Litvak M E, Brunsell N A, Peters W, Van Der Laan-Luijkx I T. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(21): 5880-5885. DOI:10.1073/pnas.1519620113 |
| [3] |
Sha Z Y, Bai Y F, Li R R, Lan H, Zhang X L, Li J, Liu X F, Chang S J, Xie Y C. The global carbon sink potential of terrestrial vegetation can be increased substantially by optimal land management. Communications Earth & Environment, 2022, 3(1): 8. |
| [4] |
Canadell J G, Pataki D E, Pitelka L F. Terrestrial ecosystems in a changing world. Berlin: Springer, 2007.
|
| [5] |
冯险峰, 孙庆龄, 林斌. 区域及全球尺度的NPP过程模型和NPP对全球变化的响应. 生态环境学报, 2014, 23(3): 496-503. DOI:10.3969/j.issn.1674-5906.2014.03.020 |
| [6] |
Del Grosso S, Parton W, Stohlgren T, Zheng D L, Bachelet D, Prince S, Hibbard K, Olson R. Global potential net primary production predicted from vegetation class, precipitation, and temperature. Ecology, 2008, 89(8): 2117-2126. DOI:10.1890/07-0850.1 |
| [7] |
Poungparn S, Komiyama A, Umnouysin S, Rodtassana C, Sangtiean T, Maknual C, Pravinvongvuthi T, Suchewaboripont V, Kato S. Ten-Year Estimation of Net Primary Productivity in a Mangrove Forest under a Tropical Monsoon Climate in Eastern Thailand: Significance of the Temperature Environment in the Dry Season. Forests, 2020, 11(9): 987. DOI:10.3390/f11090987 |
| [8] |
Field C B, Behrenfeld M J, Randerson J T, Falkowski P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science, 1998, 281(5374): 237-240. DOI:10.1126/science.281.5374.237 |
| [9] |
郭睿妍, 田佳, 杨志玲, 杨泽康, 苏文瑞, 刘文娟. 基于GEE平台的黄河流域森林NPP时空变化特征. 生态学报, 2022, (13): 1-9. (2022-03-18). http://kns.cnki.net/kcms/detail/11.2031.Q.20220316.1720.030.html.
|
| [10] |
Zhang Y J, Yu G R, Yang J, Wimberly M C, Zhang X Z, Tao J, Jiang Y B, Zhu J T. Climate-driven global changes in carbon use efficiency. Global Ecology and Biogeography, 2014, 23(2): 144-155. DOI:10.1111/geb.12086 |
| [11] |
Wang J M, Sun R, Zhang H L, Xiao Z Q, Zhu A R, Wang M J, Yu T, Xiang K L. New global MuSyQ GPP/NPP remote sensing products from 1981 to 2018. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing, 2021, 14: 5596-5612. DOI:10.1109/JSTARS.2021.3076075 |
| [12] |
Liu F H, Xu C Y, Yang X X, Ye X C. Controls of Climate and Land-Use Change on Terrestrial Net Primary Productivity Variation in a Subtropical Humid Basin. Remote Sensing, 2020, 12(21): 3525. DOI:10.3390/rs12213525 |
| [13] |
Ge W Y, Deng L Q, Wang F, Han J Q. Quantifying the contributions of human activities and climate change to vegetation net primary productivity dynamics in China from 2001 to 2016. Science of The Total Environment, 2021, 773: 145648. DOI:10.1016/j.scitotenv.2021.145648 |
| [14] |
Yu D S, Li Y H, Yin B L, Wu N, Ye R H, Liu G X. Spatiotemporal variation of net primary productivity and its response to drought in Inner Mongolian desert steppe. Global Ecology and Conservation, 2022, 33: e01991. DOI:10.1016/j.gecco.2021.e01991 |
| [15] |
Wu L Z, Ma X F, Dou X, Zhu J T, Zhao C Y. Impacts of climate change on vegetation phenology and net primary productivity in arid Central Asia. Science of the Total Environment, 2021, 796: 149055. DOI:10.1016/j.scitotenv.2021.149055 |
| [16] |
Jackson H, Prince S D. Degradation of net primary production in a semi-arid rangeland. Biogeosciences, 2016, 13(16): 4721-4734. DOI:10.5194/bg-13-4721-2016 |
| [17] |
刘杰, 汲玉河, 周广胜, 周莉, 吕晓敏, 周梦子. 2000-2020年青藏高原植被净初级生产力时空变化及其气候驱动作用. 应用生态学报, 2022, 33(6): 1533-1538. |
| [18] |
Xiao Z Q, Liang S L, Sun R, Wang J D, Jiang B. Estimating the fraction of absorbed photosynthetically active radiation from the MODIS data based GLASS leaf area index product. Remote Sensing of Environment, 2015, 171: 105-117. DOI:10.1016/j.rse.2015.10.016 |
| [19] |
Nemani R, Keeling C D, Hashimoto H, Jolly W M, Piper S C, Tucker C J, Myneni R B, Running S W. Climate-Driven Increases in Global Terrestrial Net Primary Production from 1982 to 1999. Science, 2003, 300(5625): 1560-1563. DOI:10.1126/science.1082750 |
| [20] |
Zhao M S, Running S W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 Through 2009. Science, 2010, 329(5994): 940-943. DOI:10.1126/science.1192666 |
| [21] |
Tum M, Zeidler J N, Günther K P, Esch T. Global NPP and straw bioenergy trends for 2000-2014. Biomass and Bioenergy, 2016, 90: 230-236. DOI:10.1016/j.biombioe.2016.03.040 |
| [22] |
Hubau W, Lewis S L, Phillips O L, Affum-Baffoe K, Beeckman H, Cuní-Sanchez A, Daniels A K, Ewango C E N, Fauset S, Mukinzi J M, Sheil D, Sonké B, Sullivan M J P, Sunderland T C H, Taedoumg H, Thomas S C, White L J T, Abernethy K A, Adu-Bredu S, Amani C A, Baker T R, Banin L F, Baya F, Begne S K, Bennett A C, Benedet F, Bitariho R, Bocko Y E, Boeckx P, Boundja P, Brienen R J W, Brncic T, Chezeaux E, Chuyong G B, Clark C J, Collins M, Comiskey J A, Coomes D A, Dargie G C, De Haulleville T, Kamdem M N D, Doucet J L, Esquivel-Muelbert A, Feldpausch T R, Fofanah A, Foli E G, Gilpin M, Gloor E, Gonmadje C, Gourlet-Fleury S, Hall J S, Hamilton A C, Harris D J, Hart T B, Hockemba M B N, Hladik A, Ifo S A, Jeffery K J, Jucker T, Yakusu E K, Kearsley E, Kenfack D, Koch A, Leal M E, Levesley A, Lindsell J A, Lisingo J, Lopez-Gonzalez G, Lovett J C, Makana J R, Malhi Y, Marshall A R, Martin J, Martin E H, Mbayu F M, Medjibe V P, Mihindou V, Mitchard E T A, Moore S, Munishi P K T, Bengone N N, Ojo L, Ondo F E, Peh K S H, Pickavance G C, Poulsen A D, Poulsen J R, Qie L, Reitsma J, Rovero F, Swaine M D, Talbot J, Taplin J, Taylor D M, Thomas D W, Toirambe B, Mukendi J T, Tuagben D, Umunay P M, Van Der heijden G M F, Verbeeck H, Vleminckx J, Willcock S, Wöll H, Woods J T, Zemagho L. Asynchronous carbon sink saturation in African and Amazonian tropical forests. Nature, 2020, 579(7797): 80-87. DOI:10.1038/s41586-020-2035-0 |
| [23] |
魏凤英. 现代气候统计诊断与预测技术(第二版). 北京: 气象出版社, 2007.
|
| [24] |
Yang Y J, Wang S J, Bai X Y, Tan Q, Li Q, Wu L H, Tian S Q, Hu Z Y, Li C J, Deng Y H. Factors Affecting Long-Term Trends in Global NDVI. Forests, 2019, 10(5): 372. DOI:10.3390/f10050372 |
| [25] |
Kendall M G. A New Measure of Rank Correlation. Biometrika, 1938, 30(1/2): 81-93. DOI:10.2307/2332226 |
| [26] |
袁丽华, 蒋卫国, 申文明, 刘颖慧, 王文杰, 陶亮亮, 郑华, 刘孝富. 2000-2010年黄河流域植被覆盖的时空变化. 生态学报, 2013, 33(24): 7798-7806. |
| [27] |
Granero M A S, Segovia J E T, Pérez J G. Some comments on Hurst exponent and the long memory processes on capital markets. Physica A: Statistical Mechanics and its Applications, 2008, 387(22): 5543-5551. DOI:10.1016/j.physa.2008.05.053 |
| [28] |
严恩萍, 林辉, 党永峰, 夏朝宗. 2000-2012年京津风沙源治理区植被覆盖时空演变特征. 生态学报, 2014, 34(17): 5007-5020. |
| [29] |
Peng J, Liu Z H, Liu Y H, Wu J S, Han Y N. Trend analysis of vegetation dynamics in Qinghai-Tibet Plateau using Hurst Exponent. Ecological Indicators, 2012, 14(1): 28-39. DOI:10.1016/j.ecolind.2011.08.011 |
| [30] |
王建邦, 赵军, 李传华, 朱钰, 康重阳, 高超. 2001-2015年中国植被覆盖人为影响的时空格局. 地理学报, 2019, 74(3): 504-519. |
| [31] |
Mandelbrot B B, Wallis J R. Robustness of the rescaled range R/S in the measurement of noncyclic long run statistical dependence. Water Resources Research, 1969, 5(5): 967-988. DOI:10.1029/WR005i005p00967 |
| [32] |
李美丽, 尹礼昌, 张园, 苏旭坤, 刘国华, 王晓峰, 奥勇, 伍星. 基于MODIS-EVI的西南地区植被覆盖时空变化及驱动因素研究. 生态学报, 2021, 41(3): 1138-1147. |
| [33] |
李茂华, 都金康, 李皖彤, 李闰洁, 吴森垚, 王珊珊. 1982-2015年全球植被变化及其与温度和降水的关系. 地理科学, 2020, 40(5): 823-832. |
| [34] |
Beck H E, Zimmermann N E, Mcvicar T R, Vergopolan N, Berg A, Wood E F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Scientific Data, 2018, 5(1): 180214. |
| [35] |
Gang C C, Wang Z N, You Y F, Liu Y, Xu R T, Bian Z H, Pan N Q, Gao X R, Chen M X, Zhang M. Divergent responses of terrestrial carbon use efficiency to climate variation from 2000 to 2018. Global and Planetary Change, 2022, 208: 103709. |
| [36] |
Huang M T, Piao S L, Ciais P, Peñuelas J, Wang X H, Keenan T F, Peng S S, Berry J A, Wang K, Mao J F, Alkama A, Cescatti A, Cuntz M, De Deurwaerder H, Gao M D, He H, Liu Y W, Luo Y Q, Myneni R B, Niu S L, Shi X Y, Yuan W P, Verbeeck H, Wang T, Wu J, Janssens I A. Air temperature optima of vegetation productivity across global biomes. Nature Ecology & Evolution, 2019, 3(5): 772-779. |
| [37] |
Shen M G, Piao S L, Cong N, Zhang G X, Jassens I A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Global Change Biology, 2015, 21(10): 3647-3656. |
| [38] |
Väisänen M, Krab E J, Dorrepaal E. Carbon dynamics at frost-patterned tundra driven by long-term vegetation change rather than by short-term non-growing season warming. Biogeochemistry, 2017, 136(1): 103-117. |
| [39] |
Thomey M L, Collins S L, Vargas R, Johnson J E, Brown R F, Natvig D O, Friggens M T. Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan Desert grassland. Global Change Biology, 2011, 17(4): 1505-1515. |
| [40] |
Corlett R T. Impacts of warming on tropical low land rainforests. Trends in Ecology & Evolution, 2011, 26(11): 606-613. |
| [41] |
Keenan T F, Riley W J. Greening of the land surface in the world's cold regions consistent with recent warming. Nature Climate Change, 2018, 8(9): 825-828. |
| [42] |
Wu Z T, Dijkstra P, Koch G W, Peñuelas J, Hungate B A. Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation. Global Change Biology, 2011, 17(2): 927-942. |
| [43] |
Wang J S, Chen F H, Jin L Y, Bai H Z. Characteristics of the dry/wet trend over arid central Asia over the past 100 years. Climate Research, 2010, 41(1): 51-59. |
| [44] |
Gill A L, Gallinat A S, Sanders-DeMott R, Rigden A J, Gianotti D J S, Mantooth J A, Templer P H. Changes in autumn senescence in northern hemisphere deciduous trees: a meta-analysis of autumn phenology studies. Annals of Botany, 2015, 116(6): 875-888. |
| [45] |
Grossiord C, Buckley T N, Cernusak L A, Novick K A, Poulter B, Siegwolf R T W, Sperry J S, McDowell N G. Plant responses to rising vapor pressure deficit. New Phytologist, 2020, 226(6): 1550-1566. |
| [46] |
Anderegg W R L. Spatial and temporal variation in plant hydraulic traits and their relevance for climate change impacts on vegetation. New Phytologist, 2015, 205(3): 1008-1014. |
| [47] |
Li C H, Zhou M, Dou T B, Zhu T B, Yin H H, Liu L H. Convergence of global hydrothermal pattern leads to an increase in vegetation net primary productivity. Ecological Indicators, 2021, 132: 108282. |
| [48] |
Milesi C, Hashimoto H, Running S W, Nemani R R. Climate variability, vegetation productivity and people at risk. Global and Planetary Change, 2005, 47(2/4): 221-231. |
| [49] |
茆杨, 蒋勇军, 张彩云, 乔伊娜, 吕同汝, 邱菊. 近20年来西南地区植被净初级生产力时空变化与影响因素及其对生态工程响应. 生态学报, 2022, 42(7): 2878-2890. |
| [50] |
Ugbaje S U, Odeh I O A, Bishop T F A, Li J L. Assessing the spatio-temporal variability of vegetation productivity in Africa: quantifying the relative roles of climate variability and human activities. International Journal of Digital Earth, 2017, 10(9): 879-900. |
| [51] |
Li H, Zhang H Y, Li Q X, Zhao J J, Guo X Y, Ying H, Deng G R, Wu R H, Wang S L. Vegetation productivity dynamics in response to climate change and human activities under different topography and land cover in northeast China. Remote Sensing, 2021, 13(5): 975. |
| [52] |
安秀江, 张凌云, 范泽熙, 杨广斌, 肖钊富. 贵阳市植被净初级生产力与景观格局时空分布及相关性研究. 贵州师范大学学报: 自然科学版, 2021, 39(4): 30-38, 69-69. |
| [53] |
刘静, 温仲明, 刚成诚. 黄土高原不同植被覆被类型NDVI对气候变化的响应. 生态学报, 2020, 40(2): 678-691. |