 2023, Vol. 43
2023, Vol. 43文章信息
- 朱玉, 张文广, 管强, 田雪, 赵予熙, 姜明, 肖盈丽, 王瑶, 赵琬婧, 王瑜, 原卉
- ZHU Yu, ZHANG Wenguang, GUAN Qiang, TIAN Xue, ZHAO Yuxi, JIANG Ming, XIAO Yingli, WANG Yao, ZHAO Wanjing, WANG Yu, YUAN Hui
- 三江自然保护区自然和干扰湿地节肢动物群落组成和生态指示
- Composition and indicative characteristics of arthropod communities in natural and disturbed wetlands of the Sanjiang Nature Reserve
- 生态学报. 2023, 43(9): 3819-3827
- Acta Ecologica Sinica. 2023, 43(9): 3819-3827
- http://dx.doi.org/10.5846/stxb202204251154
-
文章历史
- 收稿日期: 2022-04-25
- 网络出版日期: 2023-02-21






2. 东北师范大学草地科学研究所/植被生态科学教育部重点实验室, 长春 130024;
3. 黑龙江三江国家级自然保护区管理局, 佳木斯 156500
2. Institute of Grassland Science, Northeast Normal University, Key Laboratory of Vegetation Ecology, Ministry of Education, Chanchun 130024, China;
3. Bureau of Heilongjiang Sanjiang National Nature Reserve, Jiamusi 156500, China
湿地生态系统具有重要的水文功能、生物地球化学功能和生态功能, 同时湿地具有丰富的物种多样性, 为许多生物提供了良好的栖息地和生存场所。然而长期以来, 人为不合理利用导致了湿地生态系统发生退化[1—2]。据《全球湿地展望2021》报导, 湿地是当前全球最受威胁的生态系统:从1970到2015年期间, 全球湿地面积下降了35%, 是森林消失速度的三倍[3]。湿地的退化进一步导致全球生物多样性的丧失, 约25%依赖湿地环境的物种正濒临灭绝的威胁[4—5]。
在湿地生物群落中, 节肢动物(主要是昆虫)种类丰富、分布广泛, 是湿地生物多样性的重要组成部分[6]。节肢动物是湿地植物主要的采食者, 是珍禽和其他生物的重要食源, 也是高效的传粉者以及分解者, 在调控湿地养分循环和温室气体排放中发挥着关键作用[6—8]。同时, 节肢动物由于时代周期短, 易于采集, 对环境变化敏感, 近年来被认为是淡水湿地生态系统的良好指示生物[9—10]。探明节肢动物的指示作用对于早期预警湿地退化具有重要意义。
黑龙江三江国家级自然保护区(简称“三江保护区”)位于黑龙江省东北部的三江平原, 东临乌苏里江、北隔黑龙江, 与俄罗斯毗邻, 是黑乌两江汇合的三角地带。三江保护区是三江平原面积最大的湿地保护区, 以保护东方白鹳、丹顶鹤、大天鹅等珍贵水禽及其所依赖的沼泽湿地为主要保护对象[11—12]。然而受气候变化和人类活动的频繁干扰, 三江保护区自然湿地面积不断减少和退化, 湿地被大量开垦为水田和旱地, 垦殖和沟渠化造成自然栖息地和生物多样性受损, 湿地生态系统结构功能面临严峻挑战和威胁[13—14]。
三江平原沼泽湿地长期受农业开发等人为活动影响胁迫, 目前对此区域的研究多集中在植物群落演替以及植物种子库方面[11, 15]。相较鸟类、鱼类和其它底栖无脊椎动物[12, 16—17], 尚缺乏关于人类活动影响下湿地陆生无脊椎动物群落变化的研究。人类干扰会造成湿地植被和微环境条件等的改变, 这种变化是否会对节肢动物群落组成和多样性产生影响还未见报道。本研究通过对三江保护区自然和干扰湿地的节肢动物群落进行调查, 探究人类活动影响下节肢动物群落组成和数量差异, 研究节肢动物对湿地环境变化的生态指示功能, 以期为该区域湿地生物多样性保护和湿地生态健康评价提供参考。
1 材料与方法 1.1 研究区和采样点概况研究区位于黑龙江省东北部的三江保护区(47° 44′ 40″ N—48° 8′ 20″ N, 134° 36′ 12″ E—134° 4′ 38″ E), 面积为198089 hm2。该区域属温带湿润大陆性季风气候, 年平均气温为2.5 ℃, 年降水量为558 mm。保护区内分布着57条河流和200多个湖泡, 属于低冲积平原沼泽湿地, 是典型的低地高寒湿地[4—5]。保护区湿地植被类型多样, 优势种包括灰脉薹草(Carex appendiculata)、瘤囊薹草(Carex schmidtii)、漂筏薹草(Carex pseudocuraica)、毛薹草(Carex lasiocarpa)、小叶章(Calamagrostis angustifolia)、芦苇(Phragmites australis)、菰(Zizania latifolia)、香蒲(Typha orientalis)和狭叶甜茅(Glyceria spiculosa)等。我们在三江保护区内沿江共布设23个采样点(其中自然湿地8处, 干扰湿地15处), 各采样点的基本信息见表 1。在2020年夏季(7月17日—26日)对各采样点的节肢动物群落进行采集。
| 样点编号 Sample ID |
样点名 Location |
经纬度 Geographical position |
湿地类型 Wetland type |
样点描述 Sample description |
| S1 | 抓吉 | 48°10′4.30″N 134°38′22.55″E | 自然湿地 | 薹草群落, 保护区长期定位站, 无人为影响 |
| S2 | 抓吉门前1 | 48°9′49.20″N 134°38′56.73″E | 自然湿地 | 薹草群落, 伴生植物丰富, 无人为影响 |
| S3 | 姜明礼岗1 | 47°57′6.25″N 134°31′19.85″E | 自然湿地 | 小叶章-薹草群落, 无人为影响 |
| S4 | 海清 | 47°51′56.09″N 134°39′43.20″E | 自然湿地 | 薹草-芦苇群落, 无人为影响 |
| S5 | 永发2 | 47°40′5.80″N 134°40′1.43″E | 自然湿地 | 薹草-菖蒲群落, 无人为影响 |
| S6 | 鸭绿河2 | 48°3′49.464″N 134°0′41.198″E | 自然湿地 | 薹草群落, 伴生植物丰富, 无人为影响 |
| S7 | 鸭绿河5 | 48°0′41.569″N 133°52′04.588″E | 自然湿地 | 薹草群落, 伴生植物丰富, 无人为影响 |
| S8 | 鸭绿河6 | 47°55′29.208″N 133°52′04.487″E | 自然湿地 | 塔头薹草-芦苇群落, 无人为影响 |
| S9 | 抓吉门前2 | 48°8′0.40″N 134°36′22.35″E | 干扰湿地 | 芦苇群落, 人为影响较小, 有旱化现象 |
| S10 | 抓吉门前3 | 48°7′46.79″N 134°35′44.65″E | 干扰湿地 | 芦苇群落, 周围存在沟渠化现象 |
| S11 | 姜明礼岗2 | 47°54′50.34″N 134°33′31.63″E | 干扰湿地 | 小叶章-薹草群落, 沟渠化, 临近水稻田 |
| S12 | 海兴 | 47°53′24.27″N 134°35′42.49″E | 干扰湿地 | 塔头薹草群落, 旱化现象 |
| S13 | 大甸子 | 47°45′41.96″N 134°40′1.12″E | 干扰湿地 | 小叶章-薹草群落, 有灌木侵入, 旱化 |
| S14 | 永发1 | 47°38′17.57″N 134°38′43.56″E | 干扰湿地 | 芦苇-薹草群落, 孤立湿地, 旱化 |
| S15 | 永发3 | 47°35′50.31″N 134°36′31.62″E | 干扰湿地 | 芦苇-薹草群落, 沟渠化, 临近水稻田 |
| S16 | 四合2 | 47°30′43.24″N 134°30′15.43″E | 干扰湿地 | 苔草群落, 旱化 |
| S17 | 四合4 | 47°28′24.93″N 134°22′26.69″E | 干扰湿地 | 湿地旱化, 灌木侵入薹草群落 |
| S18 | 鸭绿河3 | 48°2′54.892″N 133°57′57.733″E | 干扰湿地 | 孤立湿地, 临近玉米田 |
| S19 | 鸭绿河4 | 48°3′07.438″N 133°55′50.966″E | 干扰湿地 | 薹草群落, 临近道路, 有人为活动干扰 |
| S20 | 鸭绿河7 | 48°5′31.243″N 134°3′01.408″E | 干扰湿地 | 菰-香蒲群落, 孤立湿地, 临近玉米田和大豆田 |
| S21 | 浓江蒙古点 | 47°58′06.344″N 133°56′33.389″E | 干扰湿地 | 芦苇群落, 灌木侵入, 临近道路和村庄 |
| S22 | 浓桥1 | 48°12′04.439″N 134°16′59.560″E | 干扰湿地 | 芦苇-香蒲群落, 放牧, 临近道路 |
| S23 | 浓桥2 | 48°0′37.897″N 134°6′10.710″E | 干扰湿地 | 小叶章-菰-香蒲群落, 孤立湿地, 临近道路 |
在研究区每处采样点设置1个30 m×30 m的样方。采用扫网法对样方内的节肢动物进行收集, 该方法是采集节肢动物较为有效的手段之一, 可以对栖居在植物群落中, 尤其是低空飞行的节肢动物类群进行较为全面的收集[18—19]。扫网由轻质的200目棉布制成, 直径为40 cm。选取晴好的、微风或无风的天气(风速 < 3.5 m/s)对节肢动物进行取样, 取样时间为9:00到15:00之间, 该时间段内节肢动物活动比较频繁。在每个样方内采用“Z”字形样带进行取样, 采集者沿样带以平均1 m/s的速度前行, 在样带上扫30网。进行两次采样重复, 混合两个重复样品为一个采样点的样品。将收集到的节肢动物迅速放入装有乙酰乙酯的毒瓶中进行毒杀, 然后放入75%酒精内保存, 贴上标签带回实验室。在实验室中将节肢动物样品进行分拣, 利用体式显微镜(Nikon SMZ745T)对样品进行形态鉴定。根据《昆虫分类》、《昆虫分类检索》、《中国昆虫生态大图鉴》及相关文献[20—22], 将全部标本鉴定到科。统计每个采样点的节肢动物数量, 计算多样性指数。
1.3 数据分析节肢动物群落生物多样性指数采用Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数进行分析, 计算公式如下[23]:
Shannon-Wiener多样性指数(H):
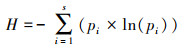
|
Simpson优势度指数(C):
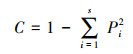
|
Pielou均匀度指数(E):

|
式中:S为物种丰富度, Pi为第i个节肢动物种类的多度比例, 即Pi=Ni/N, Ni表示第i个节肢动物种类的多度, N表示所有节肢动物种类的多度总和。
为识别不同湿地类型对节肢动物群落组成的影响, 使用基于Bray-Curtis相似性作为距离度量的聚类分析和非度量多纬尺度排序(NMDS), 然后使用相似性分析(ANOSIM)进行群落差异的显著性检验。采用指示值法评估不同节肢动物分类群对不同湿地类型的指示作用[16], 指示值取值范围为0(无指示作用)到1(绝对指示者), 并进行蒙特-卡罗检验以测试指示作用显著性。使用t检验分析不同湿地类型间节肢动物数量和多样性指数的差异。聚类分析和NMDS使用PRIMER7软件, 指示物种筛选和方差分析使用R语言(4.1.3)进行, 统计检验显著水平以P≤0.05为准。
2 结果与分析 2.1 节肢动物群落组成共采集到节肢动物1825只, 经鉴定, 共有49个分类单元, 隶属2纲10目47科(表 2)。主要以昆虫纲为主, 其中半翅目10科、鞘翅目9科、膜翅目9科、双翅目8科、直翅目2科。其中, 自然湿地共采集节肢动物1266头, 共35个分类单元, 干扰湿地共采集节肢动物559头, 共40个分类单元, 两种湿地类型有26个共同分类单元(表 2)。自然湿地中双翅目分类单元数占比最高, 干扰湿地中半翅目昆虫分类单元数占比最高, 两种湿地类型内均为双翅目昆虫个体数占比最高。不同湿地类型各节肢动物类群组成见图 1。
| 分类单元 Taxonomic groups |
自然湿地 Natural wetlands |
干扰湿地 Disturbed wetland |
分类单元 Taxonomic groups |
自然湿地 Natural wetlands |
干扰湿地 Disturbed wetland |
|
| 昆虫纲Insecta | 叶甲科Chrysomelidae | +++ | ++ | |||
| 半翅目Hemiptera | 叩甲科Elateridae | — | + | |||
| 蝽科Pentatomidae | + | + | 球棒甲科Monotomidae | + | — | |
| 花蝽科Anthocoridae | + | + | 象甲科Curculionidae | — | + | |
| 姬蝽科Nabidae | ++ | ++ | 蚁形甲科Anthicidae | + | — | |
| 猎蝽科Reduviidae | — | + | 瓢甲科Coccinellidae | + | ++ | |
| 盲蝽科Miridae | ++ | + | 蜻蜓目Odonata | |||
| 跷蝽科Berytidae | + | — | 蟌科Coenagrionidae | + | ++ | |
| 长蝽科Lygaeidae | — | ++ | 双翅目Diptera | |||
| 缘蝽科Coreidae | — | + | 大蚊科Tipulidae | + | + | |
| 叶蝉科Cicadellidae | +++ | +++ | 虻科Tabanidae | + | + | |
| 蚜科Aphididae | + | — | 食蚜蝇科Syrphidae | + | + | |
| 鳞翅目Lepidoptera | 水虻科Stratiomyidae | — | ++ | |||
| 螟蛾科Pyralidae | + | + | 水蝇科Ephydridae | + | — | |
| 毛翅目Trichoptera | 蚊科Culicidae | +++ | + | |||
| 径石蛾科Ecnomidae | — | + | 摇蚊科Chironomidae | +++ | +++ | |
| 膜翅目Hymenoptera | 长足虻科Dolichopodidae | + | — | |||
| 姬蜂总科Ichneumonoidea | +++ | ++ | 蝇类Flies | +++ | +++ | |
| 蜜蜂科Apidae | — | + | 缨翅目Thysanoptera | |||
| 泥蜂科Sphecidae | — | + | 蓟马科Thripidae | — | + | |
| 切叶蜂科Megachilidae | — | + | 直翅目Orthoptera | |||
| 隧蜂科Halictidae | — | + | 蝗科Acrididae | + | ++ | |
| 小蜂总科Chalcidoidea | ++ | + | 螽斯科Tettigoniidae | + | ++ | |
| 蚁科Formicidae | + | ++ | 蛛形纲Arachnida | |||
| 瘿蜂科CyniPidae | + | — | 蜘蛛目Araneae | |||
| 蛛蜂科Pompilidae | + | — | 狼蛛科Lycosidae | — | + | |
| 鞘翅目Coleoptera | 跳蛛科Salticidae | + | + | |||
| 步甲科Carabidae | + | — | 肖蛸科Tetragnathidae | + | ++ | |
| 隐翅甲科Staphylinidae | + | + | 蟹蛛科Thomisidae | + | + | |
| 葬甲科Silphidae | — | + | 园蛛科Araneidae | + | ++ | |
| +++为个体数占总数的5%以上; ++为个体数占总数的1%—5%;+为个体数占总数的1%以下; —表示为未见 | ||||||

|
| 图 1 不同湿地类型节肢动物群落组成 Fig. 1 Arthropod community composition in natural wetlands and disturbed wetlands |
三江保护区节肢动物个体数量在不同湿地类型具有显著差异(P < 0.001)。t检验结果表明, 自然湿地各采样点的节肢动物数量(158.25±22.44)明显高于干扰湿地(37.27±3.53)(图 2)。

|
| 图 2 不同湿地类型节肢动物数量 Fig. 2 Arthropod abundance in different types of wetlands |
生物多样性是衡量生态环境的重要评价参数。三江保护区内自然湿地的物种丰富度显著高于干扰湿地(P=0.006), 自然湿地各采样点节肢动物物种数约为12种, 干扰湿地约为8种(表 3), 而Pielou均匀度指数在干扰湿地显著高于自然湿地(P=0.016), Shannon-Wiener多样性指数和Simpson优势度指数在两种湿地类型无明显差异(表 3)。
| 节肢动物群落 Invertebrate community |
自然湿地 Natural wetlands |
干扰湿地 Disturbed wetlands |
t | P |
| 物种丰富度Species richness | 11.750±0.940 | 7.867±0.810 | -3.130 | 0.006 |
| Shannon-Wiener指数Shannon-Wiener diversity index | 1.473±0.075 | 1.501±0.138 | 0.173 | 0.865 |
| Simpson指数Simpson diversity index | 0.647±0.035 | 0.666±0.047 | 0.321 | 0.751 |
| 均匀度指数Pielou′s evenness index | 0.606±0.033 | 0.742±0.040 | 2.613 | 0.016 |
依据聚类结果得到NMDS图。NMDS(Stress=0.12)分析结果表明, 研究区内各样点节肢动物群落结构相似性较低, 自然湿地和干扰湿地节肢动物群落组成有较明显的差异。ANOSIM分析表明, 自然湿地和干扰湿地节肢动物群落差异显著(R=0.579, P=0.001)。自然湿地和干扰湿地在相似度为31时, 显示了23个采样点可以分为4个组(图 3)。除样点S1和S22以外, 自然湿地样点S2—S8以及干扰湿地S9—S12、S14、S16、S19、S23分为一组(组1), 干扰湿地S13、S15、S17、S18、S20、S21分为一组(组2)。在相似度为40时, 组1的自然湿地样点和干扰湿地样点刚好分裂成两个组(图 3)。总体来看保护区各采样点可以分为三个组:自然湿地S2—S8为一组, 干扰湿地S9—S12、S14、S16、S19、S23为一组, 干扰湿地S13、S15、S17、S18、S20、S21为一组。对三个组进行相似性分析, ANOSIM分析结果显示三个组的节肢动物群落间存在显著差异(R=0.677, P=0.001)。

|
| 图 3 不同湿地类型节肢动物群落非度量多纬尺度分析(NMDS) Fig. 3 NMDS of abundance of arthropod community dissimilarity (Bray-Curtis) recorded in different wetland use types S1—S8为自然湿地样点S9—S23为干扰湿地样点 |
指示值法分析表明: 8个分类单元的节肢动物为自然湿地的特征指示类群(P < 0.05), 干扰湿地未发现节肢动物指示类群(表 4)。
| 湿地类型 Wetland type |
分类单元 Taxonomic groups |
指示值 Indicator values |
P | 湿地类型 Wetland type |
分类单元 Taxonomic groups |
指示值 Indicator values |
P | |
| 自然湿地Natural wetlands | 叶甲科 | 0.851 | 0.007 | 盲蝽科 | 0.449 | 0.040 | ||
| 蝇类 | 0.803 | 0.001 | 蚜科 | 0.375 | 0.033 | |||
| 摇蚊科 | 0.753 | 0.012 | 蚁形甲科 | 0.375 | 0.021 | |||
| 叶蝉科 | 0.742 | 0.033 | 干扰湿地 | — | — | — | ||
| 姬蜂总科 | 0.740 | 0.026 | Disturbed wetlands |
人类活动对湿地产生的干扰会显著影响湿地生物群落, 进而影响湿地生态系统结构和功能[2]。通过对三江保护区不同湿地类型节肢动物群落调查发现, 干扰湿地的节肢动物数量和物种丰富度和自然湿地相比都显著降低。这可能由于干扰湿地频繁受到人类活动的影响, 其植物多样性减少, 群落趋于简化; 同时湿地垦殖和沟渠修建引起湿地生境破碎化, 切断了湿地间的连通性, 阻碍了节肢动物的自然迁移, 导致种群数量和多样性下降。本研究结果表明自然湿地完整的栖息环境和充足的食物资源对于维持节肢动物群落结构非常重要。
自然湿地和干扰湿地均以双翅目为优势类群, 可能是因为双翅目如摇蚊具有较强的飞行能力, 可以减轻湿地破碎化对其种群的影响。研究表明, 在河流湿地中, 摇蚊科相较其它底栖生物类群迁移能力最强, 可以达到距离河岸17公里[24]。除了双翅目, 半翅目的叶蝉科和鞘翅目的叶甲科也是湿地环境中的优势类群。研究表明, 湿地中很多植食性甲虫(如叶甲、象甲)多以特定的寄主植物为食, 可以对入侵植物进行有效生物防治[25], 因此这些植食性甲虫具有重要的生态功能。蜘蛛目和膜翅目的姬蜂总科是三江保护区主要的捕食性和寄生性节肢动物, 这与其它类型的湿地研究结果相类似[6]。湿地中大量的叶蝉、蚜虫和甲虫为其提供了丰富的食源, 同时这些更高营养级的节肢动物和湿地植物特征以及微环境条件关系十分密切。有研究表明植物物种丰富度和植株密度与蜘蛛的多样性呈正相关关系[26]。
NMDS结果显示节肢动物群落组成受人为活动干扰的强烈影响。在相似度为40时, 自然湿地和干扰湿地节肢动物群落组成显著不同。当相似度下降至31时, 自然湿地的7个采样点和干扰湿地的8个采样点分为一组, 而干扰湿地其余6个采样点分为一组。由节肢动物群落组成相似性可以看出三江保护区湿地可能遭受不同程度的人为干扰, 即与自然湿地相近的采样点为干扰程度较轻的湿地, 与自然湿地相差较远的采样点为干扰程度较重的湿地。不同干扰程度的湿地和自然湿地相比, 其水文情势条件、植物群落特征以及人类干扰强度具有差异, 因此会通过影响生物的栖息环境质量和食物资源等对节肢动物群落产生影响。自然湿地采样点以物种丰富的薹草群落为主[11], 薹草沼泽植物群落结构复杂, 湿地水文连通性较高, 复杂的生境和丰富的食源支持了数量更多的节肢动物; 而在干扰样点, 湿地周围存在沟渠化现象, 水位下降, 植物丰富度减少, 群落结构趋于简化, 节肢动物数量出现显著下降; 在干扰程度更大的样点, 湿地受到较为强烈的人类活动如农业生产或放牧的影响, 湿地水文连通性受阻, 水体质量恶化, 植物从薹草转变为菰、香蒲占优势的群落, 物种多样性较低[15]; 一些采样点湿地出现明显的旱化, 存在灌木入侵现象, 加剧了节肢动物数量的下降。在若尔盖高寒湿地的研究同样发现, 节肢动物群落组成和数量在不同退化生境间差异显著[27]。
对比两种不同湿地类型节肢动物的多样性指数发现, Shannon-Wiener多样性指数和Simpson优势度指数在自然湿地和干扰湿地没有显著差异, 而Pielou均匀度指数在干扰湿地显著大于自然湿地。这可能由于几种优势节肢动物类群在自然湿地聚集所致, 而干扰湿地总体节肢动物数量较少且种类分布相对分散。与我们的研究类似, Lougheed等(2008)比较了美国密歇根州16处自然湿地和退化湿地, 发现退化湿地中植物群落和昆虫群落的多样性更低, 但均质性更高[28], 这说明退化湿地比自然湿地的生物群落具有更低的异质性。为了保护和恢复湿地生物多样性, 需要进一步量化湿地退化和特定的压力源, 诸如旱化、垦殖化、沟渠化对湿地生物多样性维持机制的影响。
节肢动物监测可以作为评价生态系统健康状况的一种有效手段[9, 29]。目前关于节肢动物的指示作用研究大多偏向单一生物类群, 比如鞘翅目步甲科[14, 30]、蜻蜓目[10, 31]、EPT昆虫(蜉蝣目Ephemeroptera、襀翅目Plecoptera和毛翅目Trichoptera)[32], 以及双翅目摇蚊科[33]。针对节肢动物群落水平对湿地环境变化的指示作用的研究尚不多见。本研究基于对节肢动物49个分类单元的调查发现, 鞘翅目的叶甲科、蚁形甲科, 半翅目的叶蝉科、蚜科、盲蝽科, 双翅目摇蚊科、蝇类以及膜翅目的姬蜂总科为自然湿地的指示物种。在干扰湿地未发现有指示物种存在, 这可能是由于不同干扰湿地采样点受到的胁迫因素不同, 因此很难找出针对性的节肢动物指示类群。我们的研究表明利用自然湿地或受损较小的湿地中节肢动物群落来指示以恢复生物群落为目标的湿地修复是可行的。
本研究结果表明, 三江保护区自然湿地和干扰湿地节肢动物群落组成差异显著, 且节肢动物可以作为自然湿地的指示生物。干扰湿地节肢动物的数量和物种丰富度与自然湿地相比显著降低。这意味着节肢动物相对其它生物类群, 更容易受到环境的胁迫带来的不利影响。本研究结果强调减少人为活动干扰对自然湿地的影响对于维持节肢动物多样性十分重要。目前我国针对湿地节肢动物的研究工作十分滞后, 应进一步开展节肢动物群落的长期调查监测工作, 加强对节肢动物多样性和生态功能的研究。并且研究工作应该着重考虑影响节肢动物群落构建的主要环境要素和胁迫因子, 包括植物多样性[34]、水体连通性[35]、物种间相互作用[36]、周边土地利用情况[17]及人类活动强度[37]。同时, 我国目前基于节肢动物群落的生态评价体系并不完善, 如何将不同节肢动物类群和湿地干扰程度或健康等级间建立联系还需要未来更深入的研究。
| [1] |
Oliver T H, Morecroft M D. Interactions between climate change and land use change on biodiversity: attribution problems, risks, and opportunities. WIREs Climate Change, 2014, 5(3): 317-335. DOI:10.1002/wcc.271 |
| [2] |
Craig L S, Olden J D, Arthington A H, Entrekin S, Hawkins C P, Kelly J J, Kennedy T A, Maitland B M, Rosi E J, Roy A H, Strayer D L, Tank J L, West A O, Wooten M S. Meeting the challenge of interacting threats in freshwater ecosystems: a call to scientists and managers. Elementa: Science of the Anthropocene, 2017, 5: 72. DOI:10.1525/elementa.256 |
| [3] |
Courouble M, Davidson N, Dinesen L, Fennessy S, Galewski T, Guelmami A, Kumar R, McInnes R, Perennou C, Rebelo L M, Robertson H, Segura-Champagnon L, Simpson M, Stroud D. Convention on wetlands//Dudley N, ed. Global Wetland Outlook: Special Edition 2021. Gland, Switzerland: Secretariat of the Convention on Wetlands, 2021.
|
| [4] |
Collen B, Whitton F, Dyer E E, Baillie J E M, Cumberlidge N, Darwall W R T, Pollock C, Richman N I, Soulsby A M, Böhm M. Global patterns of freshwater species diversity, threat and endemism. Global Ecology and Biogeography, 2014, 23(1): 40-51. DOI:10.1111/geb.12096 |
| [5] |
Davidson N C. How much wetland has the world lost? Long-term and recent trends in global wetland area. Marine and Freshwater Research, 2014, 65(10): 934-941. DOI:10.1071/MF14173 |
| [6] |
Batzer D P, Wu H T. Ecology of terrestrial arthropods in freshwater wetlands. Annual Review of Entomology, 2020, 65(1): 101-119. DOI:10.1146/annurev-ento-011019-024902 |
| [7] |
Boix D, Batzer D. Invertebrate assemblages and their ecological controls across the world's freshwater wetlands//Batzer D, Boix D, eds. Invertebrates in Freshwater Wetlands: An International Perspective on their Ecology. Cham: Springer, 2016: 601-639.
|
| [8] |
Zhang X H, Xin Y, Zhang Z S, Zhang Z M, Wu H T. Effects of ant colonies on molecular characteristics of dissolved organic matter in peatland soils, Northeast China. Applied Soil Ecology, 2022, 171: 104298. DOI:10.1016/j.apsoil.2021.104298 |
| [9] |
苏兰, 黄俊浩, 吴明, 吴鸿. 湿地植被演替中昆虫多样性变化研究进展. 生态学杂志, 2012, 31(6): 1577-1584. DOI:10.13292/j.1000-4890.2012.0226 |
| [10] |
Kutcher T E, Bried J T. Adult Odonata conservatism as an indicator of freshwater wetland condition. Ecological Indicators, 2014, 38: 31-39. DOI:10.1016/j.ecolind.2013.10.028 |
| [11] |
潘媛, 姜明, 丛小丽, 薛振山, 刘波, 张文广, 焦健, 赵琬婧, 王瑜, 原卉. 黑龙江三江国家级自然保护区沼泽中的野生种子植物组成与区系分析. 湿地科学, 2021, 19(3): 342-352. DOI:10.13248/j.cnki.wetlandsci.2021.03.009 |
| [12] |
赵琬婧. 黑龙江三江自然保护区成功繁殖白尾海雕. 湿地科学与管理, 2021, 17(2): 81-81. |
| [13] |
苏文辉, 于晓菲, 王国平, 栾金花, 邹元春. 沟渠化对三江平原湿地铁元素沉积过程的影响. 环境科学, 2015, 36(4): 1431-1436. DOI:10.13227/j.hjkx.2015.04.041 |
| [14] |
暴晓, 吕宪国, 张帆, 武海涛. 三江平原湿地不同土地利用生境地表甲虫群落结构. 生态与农村环境学报, 2010, 26(1): 47-51. DOI:10.3969/j.issn.1673-4831.2010.01.009 |
| [15] |
王国栋, 姜明, 何兴元, 吕宪国. 香蒲入侵薹草沼泽的机制研究进展. 湿地科学, 2018, 16(3): 288-293. |
| [16] |
管强, 武海涛, 陈展彦, 李洪宇, 芦康乐, 刘吉平, 吕宪国. 三江平原典型沼泽湿地螺类组成生态指示. 生态学报, 2018, 38(9): 3320-3327. |
| [17] |
芦康乐, 杨萌尧, 武海涛, 管强, 张科. 黄河三角洲芦苇湿地底栖无脊椎动物与环境因子的关系研究——以石油开采区与淡水补给区为例. 生态学报, 2020, 40(5): 1637-1649. |
| [18] |
刘春明, 左悦, 任炳忠. 莫莫格国家自然保护区昆虫多样性研究. 东北师大学报: 自然科学版, 2011, 43(3): 112-116. |
| [19] |
Zhu Y, Zhong Z W, Pagès J F, Finke D, Wang D L, Ma Q H, Hassan N, Zhu H, Wang L. Negative effects of vertebrate on invertebrate herbivores mediated by enhanced plant nitrogen content. Journal of Ecology, 2019, 107(2): 901-912. |
| [20] |
郑乐怡, 归鸿. 昆虫分类. 南京: 南京师范大学出版社, 1999.
|
| [21] |
李鸿兴, 隋敬之, 周士秀, 周勤, 孙洪国. 昆虫分类检索. 北京: 农业出版社, 1987.
|
| [22] |
张巍巍, 李元胜. 中国昆虫生态大图鉴. 重庆: 重庆大学出版社, 2011.
|
| [23] |
杨赵, 杨效东. 哀牢山不同类型亚热带森林地表凋落物及土壤节肢动物群落特征. 应用生态学报, 2011, 22(11): 3011-3020. |
| [24] |
Muehlbauer J D, Collins S F, Doyle M W, Tockner K. How wide is a stream? Spatial extent of the potential "stream signature" in terrestrial food webs using meta-analysis. Ecology, 2014, 95(1): 44-55. |
| [25] |
Tipping P W, Martin M R, Rayamajhi M B, Pratt P D, Gettys L A. Combining biological and mechanical tactics to suppress Melaleuca quinquenervia. Biological Control, 2018, 121: 229-233. |
| [26] |
Cattin M F, Blandenier G, Banašek-Richter C, Bersier L F. The impact of mowing as a management strategy for wet meadows on spider (Araneae) communities. Biological Conservation, 2003, 113(2): 179-188. |
| [27] |
邵珍珍, 吴鹏飞. 小型表栖节肢动物群落对高寒湿地退化的响应. 生态学报, 2019, 39(19): 6990-7001. |
| [28] |
Lougheed V L, McIntosh M D, Parker C A, Stevenson R J. Wetland degradation leads to homogenization of the biota at local and landscape scales. Freshwater Biology, 2008, 53(12): 2402-2413. |
| [29] |
Liu J N, Zhang Y Y, Xu S P, Zhao X H, Yin L F, Hua J Z, Yao L Y, Guo L H. Advances in research on the structure and diversity variation of insect communities in wetland. Agricultural Science & Technology, 2017, 18(5): 914-917, 956-956. |
| [30] |
王玉, 高光彩, 付必谦, 吴专. 北京野鸭湖湿地地表甲虫群落组成与空间分布格局. 生物多样性, 2009, 17(1): 30-42. |
| [31] |
唐春英, 张翔, 李静, 李正波, 黄木娇, 段红莲, 卢志兴, 李巧, 陈又清. 保山市青华海国家湿地公园蜻蜓目昆虫多样性及其生物指示作用. 湿地科学, 2020, 18(6): 712-718. |
| [32] |
杜瑞卿, 王庆林, 张征田, 王明伟. EPT昆虫群落分布与环境因子的相关性. 昆虫学报, 2008, 51(3): 336-341. |
| [33] |
胡忠军, 刘其根, 陈立婧, 彭自然. 上海崇明明珠湖摇蚊幼虫群落结构特征及其对水质的指示作用. 应用生态学报, 2009, 20(4): 929-936. |
| [34] |
朱慧, 彭媛媛, 王德利. 植物对昆虫多样性的影响. 生态学杂志, 2008, 27(12): 2215-2221. |
| [35] |
Wang J N, Dong Z R, Liao W G, Li C, Feng S X, Luo H H, Peng Q D. An environmental flow assessment method based on the relationships between flow and ecological response: a case study of the Three Gorges Reservoir and its downstream reach. Science China Technological Sciences, 2013, 56(6): 1471-1484. |
| [36] |
朱玉, 王德利, 钟志伟. 生态系统基于性状调节的物种间接作用: 特征、成因及后果. 生态学报, 2017, 37(23): 7781-7790. |
| [37] |
Winton R S, Calamita E, Wehrli B. Reviews and syntheses: dams, water quality and tropical reservoir stratification. Biogeosciences, 2019, 16(8): 1657-1671. |