 2023, Vol. 43
2023, Vol. 43文章信息
- 陈礼敏, 韦小练, 范泽鑫, 刘亚男, 肖春芬, 黄国良, 付培立
- CHEN Limin, WEI Xiaolian, FAN Zexin, LIU Yanan, XIAO Chunfen, HUANG Guoliang, FU Peili
- 西双版纳楝径向生长和木质部发生季节动态及其对环境因子的响应
- Seasonal radial growth and xylogenesis of Melia azedarach and its response to environmental factors in Xishuangbanna, Southwest China
- 生态学报. 2023, 43(8): 3192-3202
- Acta Ecologica Sinica. 2023, 43(8): 3192-3202
- http://dx.doi.org/10.5846/stxb202203160643
-
文章历史
- 收稿日期: 2022-03-16
- 网络出版日期: 2022-12-29





2. 中国科学院大学, 北京 100049;
3. 云南哀牢山森林生态系统国家野外科学观测研究站, 景东 676209;
4. 中国科学院核心植物园, 勐腊 666303;
5. 中国科学院西双版纳热带植物园, 园林园艺中心, 勐腊 666303;
6. 云南省国家级自然保护区勐仑管护所, 勐腊 666303
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Ailaoshan Station of Subtropical Forest Ecosystem Studies, Jingdong 676209, China;
4. Core Botanical Garden of Chinese Academy of Sciences, Mengla 666303, China;
5. Center for Gardening and Horticulture, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla 666303, China;
6. Menglun Management Institute, Xishuangbanna National Nature Reserve, Meangla 666303, China
热带森林占地球陆地表面的7%, 但储存了全球陆地的25%碳, 并占净初级生产力的三分之一[1—2]。树木的生长过程是进行碳吸收和碳同化的重要过程[3]。树木生长能随着环境变化迅速做出反应[4]。因此, 热带森林在碳吸收方面水平具有重要意义[5], 在调节全球气候系统的碳水反馈方面发挥着关键作用[2]。随着全球气候变化加剧、大气CO2浓度升高和降水格局的改变, 全球植被生长已经受到影响[6], 树木死亡率不断增加[7—8], 在热带森林中树木的生长和繁殖也受到了较大的影响[9]。
关于热带树木的径向生长如何响应气候变化, 不同的研究所得出的结论尚不统一。在过去几十年内南美亚马逊热带雨林的树木生长速率、树种更新速率和森林生产力呈增长的趋势, 生长速率的增加可能主要受大气CO2浓度升高以及资源可利用性增加的影响[10—11]。而马来西亚地区以及哥斯达黎加地区的热带雨林树木生长速率呈下降趋势[12—13], 这可能主要是由于区域降水减少和日最低温度升高导致的[12, 14];温度升高导致了中非热带森林三个物种的生长呈现整体下降的趋势[15]。而van der Sleen等发现虽然CO2浓度升高引起了植物水分利用效率的增加, 但泰国、玻利维亚和喀麦隆三个地区热带森林的树木径向生长均未显著变化[16]。
研究热带树木径向生长和木质部发生过程的季节动态, 有助于精确预测和解析热带树木径向生长的长期变化趋势及其与气象因子的关系。研究树木径向生长季节动态的方法主要包括高精度生长仪法、生长环法和微树芯法等方法。高精度生长仪可在小时和天的尺度上获得树木径向变化的数据[17], 但由于生长仪的价格昂贵, 在研究上的应用有一定的局限性。生长环法由于价格低廉, 是研究树木年内生长动态较好的一种技术手段[18]。微树芯法是监测树木年内径向生长动态的有效手段, 可以直接监测到树木木质部生长量和形成层活动各阶段细胞动态等[19]。目前, 对树木年内径向生长动态和形成层活动方面的研究相对主要集中在温带和寒带地区[20—22], 而在热带地区的研究相对较少, 且关于热带和亚热带地区的大多数研究都位于海拔较高的地区或侧重于针叶树种[23], 只有少数针对阔叶树树种的研究[24]。对于热带地区树木径向生长和木质部发生的规律及其与气象因子的关系, 也还没有获得统一的结论。在季节性降水的热带雨林中树木的形成层在雨季处于活跃状态, 旱季的降水也可诱导形成层活动的提前发生[25]。在相对湿润的哥伦比亚热带森林中, 木质部的发育主要发生在雨季;而在较为干旱的玻利维亚热带森林中, 木质部主要在雨季中期开始生长[26]。马来西亚地区热带雨林的娑罗双属植物(Shorea)的形成层活动主要受到降水、相对湿度和饱和水汽压亏缺的影响[27]。在西双版纳的喀斯特森林中, 树木的径向生长开始于旱季末期, 径向生长日增量与相对湿度和降水量呈显著正相关关系[17]。热带地区树木的形成层活动受气候因子的影响因物种以及样点而异, 因此气候因子对热带树木径向生长和木材形成的影响尚需进一步研究[28]。
云南南部的西双版纳地区保留了中国现存的最大面积的热带森林[29]。由于西双版纳地处热带的北缘, 其水热条件已经接近热带森林分布的极限。西双版纳地区的相对湿度逐年降低、旱季的有雾日也在减少[30—31], 因此, 研究该地区树木的形成层活动季节动态及其与气候因子的关系, 有助于预测在气候变化的背景下该地区热带森林树木生长的变化趋势。目前在泰国西部地区, 已经开展了对楝(Melia azedarach)径向生长年际动态及其与环境因子关系的研究, 发现楝径向生长速率与温度呈负相关关系, 而与降水呈正相关关系[32]。对西双版纳喀斯特森林中楝的径向生长季节动态的研究发现, 楝的径向日增量与降水量和相对湿度正相关, 且树木径向生长在干旱年份开始较晚[33]。在之前的这部分研究中, 只关注了树木的径向生长, 并未对木质部发育过程的变化动态进行研究。本研究选取西双版纳热带植物园内的楝为研究对象, 利用微树芯法和生长环法监测其径向生长和木质部发育季节动态, 同时结合环境因子, 分析楝的径向生长季节动态和木质部发育与环境因子之间的关系, 本研究可为预测未来气候变化对热带地区树木径向生长动态的影响提供借鉴。
1 研究地区及研究方法 1.1 研究区概况及研究树种西双版纳热带植物园地处热带北缘(21°41′N, 101°25′E), 海拔580 m, 受西南季风影响, 属于热带季风气候。年平均气温为21.9 ℃, 年降水量为1467 mm(1959年—2018年)(图 1)。一年分为雨季(5月到10月)和旱季(11月到次年4月)两个季节, 其中雨季降雨量占全年降水量80%以上。旱季又包括雾凉季(11月至次年2月)和干热季(3月—4月)。从1959年开始, 该地区温度呈持续上升的趋势, 而降水呈下降趋势, 气候逐渐干热化[34]。

|
| 图 1 西双版纳勐仑地区1959—2018年月平均温度和降水量 Fig. 1 Monthly mean air temperature (℃) and precipitation (mm) during the period 1959—2018 in Menglun, Xishuangbanna 红色的线表示月均温, 蓝色的线是月降水量; 红色的点、蓝色的直线以及深蓝色的区域分别表示干旱、潮湿和湿润的时期; 在图的右上方的数字表示年均温和年均降水量; 左边的y坐标轴上的33.3和12.2分别表示最热月(4月)平均最高温度和最冷月(1月)的平均最低温度 |
楝(Melia azedarach), 为楝科楝属落叶乔木, 广布于亚洲热带和亚热带地区, 具有清晰的年轮边界, 属于半环孔材, 是热带地区研究树木形成层活动的良好材料。同时楝也是一种良好的造林和经济树种。本研究选取西双版纳植物园内5株楝树进行树木径向生长及木质部发生的季节动态研究, 研究树木平均胸径为(48.38±4.66)cm。
1.2 树木生长动态监测 1.2.1 生长环法选取生长良好且胸径大小相近的5株楝作为研究材料。对选择的5株楝安装金属生长环进行树木直径生长量的监测。生长环的制作参考Anemaet等的方法[35]。在树干胸径1.3 m处安装生长环, 测量起始胸围大小。在安装生长环时, 如果胸径处不平整或者有要脱落的树皮, 需要进行清理和平整, 以减少因树皮水分动态变化引起的膨胀或收缩导致的树木径向变化的干扰。生长环安装好后, 需要在生长环的环带上刻出划痕作为测量的起始点, 用游标卡尺每半个月测量一次树干的周长变化, 生长环法监测时间从2021年2月—2021年12月。
1.2.2 微树芯法对上述所选择安装生长环的5株楝树, 每半个月进行一次采样, 利用特制的微型生长锥(Trephor, 意大利)沿着树干的南面进行“Z”字型采样[19]。每个微树芯样品长约15 mm, 直径约2 mm。采集的样品放在50%的酒精溶液里进行保存和固定(固定3—5天), 带回实验室后进行软化、脱水和透明(经过不同浓度的酒精、柠檬烯和石蜡)、包埋和切片。最后制成永久装片, 并在安装了偏光模块的电子显微镜(DM2500, Leica, 德国)下进行偏光拍摄。将照片在ImageJ(http://rsb.info.nih.gov/ij/)中进行处理, 测量其形成层活动各个阶段的细胞宽度(形成层细胞、扩大细胞、细胞壁加厚细胞和成熟细胞), 对每个切片的不同位置进行3次测量, 最后取其平均值(图 2)。微树芯法监测时间从2021年1月—2022年1月。

|
| 图 2 楝木质部发生完整年轮和形成层局部切片 Fig. 2 Transvers section of the xylogenesis of Melia azedarach of one whole tree ring and details of the cambial zone Ph: 韧皮部Phloem; CZ: 形成层细胞Cambial cell; EZ: 扩大期细胞Enlarging cell; WZ: 细胞壁加厚期细胞Wall thickening cell; MZ: 成熟期细胞Maturing cell; 红色实线表示年轮边界Red line indicated ring boundary |
气候资料来自于中国科学院西双版纳热带雨林生态系统研究站。在西双版纳热带植物园的园区内平整开阔地建有标准气象观测场(所属中国生态系统研究网络, CERN), 本研究主要采用的气象数据包括:气温, 相对湿度, 降水, 光合有效辐射, 平均风速等。所用数据为日平均值, 时间为2021年1月—12月。饱和水汽压(VPD)亏缺根据气温和相对湿度计算得出:
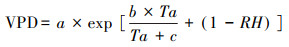
|
其中Ta为气温;RH为相对湿度;a、b、c为常数, 分别为0.614 kPa、17.502和240.97 ℃
1.4 数据分析 1.4.1 生长环数据分析采用McMahon等2015年提出的逻辑斯蒂生长模型拟合树木径向生长和物候参数[18]。其原理为:分别将每个个体12个月累积生长量(dbhdoy)与转化后的观测时间(年积日(DOY), 一年以365天计算)代入(2)
式中构建该个体的生长模型, 拟合生长曲线并计算出生长曲线的模型参数。基于模型参数再通过(3)式设置对应的生长参数d。由于树木在生长过程中会受到树木生理过程或水分动态引起的树干膨胀或收缩的干扰, 模型设定树木的生长达到生长曲线上下部渐近线截距之差(K-L)的2.5%时, 即表示树木生长开始, 此时的时间为生长开始的时间(图 3曲线左下方绿色小三角符号所在位置)。截距之差达到50%时为该树的年生长量的一半的时间, 对应的时间天数即为达到年生长量一半的时间。达到截距之差97.5%时表示生长停止, 此时的时间为树木停止生长的时间(图 3曲线右上方绿色小三角符号所在的位置)。生长曲线的拐点即为生长速率达到最大的位置, 对应的时间为该树最大生长速率达到的时间(图 3曲线拐点即绿色十字符号所在位置), 而拐点曲线的斜率则为最大生长速率。
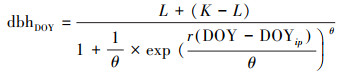
|
(2) |
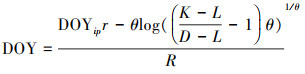
|
(3) |

|
| 图 3 西双版纳地区楝径向生长Logistic模拟曲线示意图 Fig. 3 Logistic growth curve of Melia azedarach in the Xishuangbanna 图中下方绿色三角符号所在位置表示生长开始的时间, 绿色十字符号所在位置表示生长速率达到最大的时间, 上方绿色三角符号所在位置表示结束生长的时间 |
DOY为年积日;dbhDOY为每一时间记录的胸径;DOYip为拟合生长曲线中拐点对应的天数;L为生长曲线下部渐近线;K为生长曲线上部渐近线;r为生长曲线拐点的斜率, θ为调整上部渐近线的拟合。为了和微树芯所测得的树木半径生长的数据相比较, 将直径生长的相关数据除以2转换为半径生长的数据。
月生长量是用后一个月的数据减去前一个月所得, 雨季生长量为每年的5—10月净生长量, 旱季生长量为11月到次年4月的净生长量, 最后统计树木的年、季节和月净生长量来分析楝的生长动态。在计算径向生长速率与气候数据的相关分析时, 用每半个月测定的5棵树的平均径向生长量(后一次测定减去前一次测定)与相应时间段的总降水量以及平均相对湿度、光合有效辐射、平均温度、最低温度、最高温度、饱和水汽压亏缺和风速进行Spearman相关分析。
1.4.2 微树芯数据分析利用广义加性混合模型对木质部累积生长量和生长速率进行拟合。在本研究中, 将扩大细胞开始出现的时间定义为微树芯法所获得的生长开始时间, 而将加厚期细胞停止出现的时间作为微树芯法所获得生长的结束时间。
所有的数据整理和图表绘制均在Excel 2016软件和R(3.5.1, R核心团队, 2018)软件中进行, 其中逻辑斯蒂生长模型利用“MASS”、“chron”、“RColorBrewer” 3个程序包进行, 广义加性混合模型(GAMM)利用“mgcv”程序包进行拟合。
2 结果与分析 2.1 环境因子的变化2021年的平均气温为22.7 ℃, 极端最高温为39.2 ℃(5月22日), 年降水量为1272 mm, 降水量低于该地区多年平均值(1959—2018年平均降水量1467 mm)。在2月8日出现一年中最大的降水量, 为78.4 mm。2021年1—3月的降水量为138.4 mm, 显著高于多年平均值(1959—2018年1—3月平均降水量为81.4 mm), 相对湿度在4月22日达到一年中的最小值, 为64%, 水汽压亏缺值在5月23日达到峰值, 为1.29 kPa(图 4)。

|
| 图 4 西双版纳勐仑地区2021年的气温、降水、相对湿度以及水气压亏缺的波动情况 Fig. 4 Variations in air temperature, precipitation, relative humidity and vapor pressure deficit (VPD) during 2021 in Menglun, Xishuangbanna 在气温图中红色的线表示日均气温, 灰色的阴影部分表示最高和最低气温。数据来源于中国科学院西双版纳热带雨林生态系统研究站 |
利用生长环法监测树木径向生长, 采样逻辑斯蒂模型拟合不同植株半径生长量, 结果表明:不同植株个体间存在一定的差异, 树M1在前期出现负生长的情况, 可能主要受到水分亏缺引起的茎干收缩的影响。树M3最早开始生长, 生长量最高, 为10.15 mm, 树M2是最晚结束生长, 而树M4和M5停止生长比较早, 树M5生长量最少, 为3.16 mm(图 5)。楝的径向生长主要集中在雨季, 雨季的平均生长量占全年生长量的90.9%, 旱季生长量仅占9.1%(图 5)。平均年径向生长量为(6.25±2.35)mm。在6月30日(DOY:181±6.2)生长速率最高, 为(0.056±0.02)mm/d, 此时生长量达到年生长量的50%, 持续生长时间为(209.8±9.6)d(表 1)。

|
| 图 5 楝年半径生长量及每月径向生长量在2021年的动态变化 Fig. 5 The stem radial variation and monthly radial growth of Melia azedarach during 2021 图半径生长量中不同的标志表示不同的个体 |
| 检测方法 Monitoring methods |
开始生长 Start day of growth (DOY) |
生长结束 End day of growth (DOY) |
持续时间 Duration/d |
年生长量 Annual growth/mm |
最大生长速率 Maximum growth rate/mm |
| 微树芯Microcoring | 60.2±1.6 | 345.4±3.1 | 285.2±2.4 | 7.8 | 0.039 |
| 生长环Band dendrometer | 76.6±8.6 | 286.4±6.8 | 209.8±9.6 | 6.25±2.4 | 0.056±0.020 |
楝形成层细胞全年变化趋势不显著(图 6), 这可能与在实验过程中, 形成层区域的细胞被切坏的情况较多有关。楝的扩大细胞在3月1日(DOY:60.2±1.6)左右开始出现, 此时第一个导管也已经开始形成, 在12月6日(DOY:339.8±2.5)扩大细胞停止出现。加厚期细胞开始出现时间是在3月15日(DOY:74±2.7), 在12月11日(DOY:345.4±3.1)左右细胞加厚结束, 成熟细胞在4月3日(DOY:93±3.6)左右开始出现。

|
| 图 6 西双版纳热带植物园内楝2021年木质部发生过程中形成层细胞各分化阶段的变化 Fig. 6 Changes of Melia azedarach cambium cells at different differentiation stages during the xylem formation process in 2021 in Xishuangbanna Tropical Botanical Garden |
生长环法监测的树干径向生长的开始时间(DOY:76.6±8.6)比微树芯法监测的树干径向生长时间(DOY:60.2±1.6)晚16.4 d, 同时生长环数据所获得的树干径向生长的结束时间(DOY:286.4±6.8)比微树芯监测法所观测到的树干径向生长的结束时间(DOY:345.4±3.1)提前了59 d(表 1)。生长环所测定的年生长量比微树芯法所测定木质部生长量少20%, 而生长环法测定的最大生长速率比微树芯法测定的最大生长速率大40%。
2.4 树干径向季节生长对气候的响应树木径向生长速率与降水(r=0.77, P < 0.001)、平均温度(r=0.61, P < 0.05)和最低温度(r=0.67, P < 0.001)显著正相关。与风速(r=-0.45, P < 0.05)显著负相关。而与光合有效辐射, 相对湿度, 水汽压亏缺值和最高温度无显著相关(图 7)。

|
| 图 7 楝树径向生长速率与气候因子(2021)的Spearman相关系数 Fig. 7 Correlation coefficients between radial growth rates of Melia azedarach and climatic factors in 2021 PAR: 光合有效辐射Photosynthetically active radiation; Pre: 降水量Precipitation; RH: 相对湿度Relative humidity; Ta-max: 最高温度Maximum temperature; Ta-mean: 平均温度Mean temperature; Ta-min: 最低温度Minimum temperature; VPD: 饱和水汽压亏缺Vapor pressure deficit; WS: 平均风速Mean wind speed. 其中虚线表示P < 0.05的临界值 |
关于热带树木是否具有休眠期, 目前还存在一定的争议。Abernethy等认为热带植物缺乏冬季休眠期, 植物可以持续生长和繁殖[36]。但通过模型模拟广泛分布于热带和亚热带地区楝的展叶和开花物候的研究发现, 在中国东南部热带地区的树木具有休眠期, 且低温通常不足以打破芽休眠, 因此在气候变暖的背景下展叶和开花物候延迟[37]。而在本研究中楝在3月1日出现扩大细胞, 在12月11日细胞壁加厚结束(图 6), 因此, 楝的休眠期约为2多个月。韦小练等对热带喀斯特森林多花白头树(Garuga floribunda)的研究发现, 扩大细胞在第70 d出现, 而细胞壁加厚结束在是在第263 d, 多花白头树的休眠期为172 d[38], 这些结果都显示了热带部分植物具有休眠期。而季节性降水模式是控制热带树木形成层活动的关键因素之一, 决定热带树木生长季节的持续时间[25]。每年2—3个月的旱季(月均降雨量少于60 mm), 可能是导致热带树木形成层休眠的原因[39]。西双版纳地区旱季的长度(11月至次年的4月为干季)达6个月, 旱季的月均降水量仅为41.2 mm。因此, 较长的旱季使得该地区的落叶树木具有明显的休眠期。楝的生长量主要集中在雨季, 雨季生长量占全年生长量的90.9%(图 5)。对非洲科特迪瓦Rubino地区的柚木(Tectona grandis)研究发现, 形成层和径向生长最活跃的时期是6月雨季[40]。对哀牢山中山湿性常绿阔叶林树木径向生长季节动态研究也发现, 树木的生长主要在雨季, 且雨季生长量占全年生长量的93%[41]。但Huang等研究发现, 在亚热带地区马尾松在旱季末期的生长速率较快[23]。
本研究中生长环法和微树芯法所测定的径向生长开始和结束时间存在一定的差异(表 1)。这主要是因为生长环法计算树木径向生长的开始时间和结束时间分别是当生长量达到年生长总量的2.5%和97.5%的实际时间[42], 而由于楝树在旱季受到一定的水分亏缺, 因此生长环法不能准确捕捉到树木径向生长开始和结束的时间。微树芯法能很好的区分各个时期细胞变化情况, 可以准确的分辨树木开始生长和结束生长的时间[43]。前期的研究也发现高精度生长仪法所估算的热带树木多花白头树的径向生长开始时间晚于微树芯法, 而生长仪法监测出来的生长结束时间早于微树芯法[38]。由于西双版纳地区11月到次年4月是旱季, 降水量较少, 树木的径向变化受到水分亏缺的影响发生收缩, 因此生长环或生长仪法均无法准确捕捉到木质部生长开始的时间。本研究中生长环法和微树芯法所获得的楝的年径向生长量接近(图 5, 图 6)。而韦小练等的结果发现高精度生长仪法所获得的年径向生长量是微树芯法的2倍[38], 这与多花白头树的树皮厚于楝有关。生长仪和生长环所测得的数据包含了由于树皮的收缩膨胀以及韧皮部生长的信息。生长环法由于成本较低, 可以大范围长时间的监测树木的径向生长变化, 但是不能监测形成层各个阶段的细胞变化, 特别是细胞壁加厚期的变化[44], 而微树芯法能很好的监测木质部细胞各个阶段的变化。由于楝是一个先锋树种, 径向生长速率较快, 因此生长环的方法也能较好地反映其径向生长的季节动态。但对于生长速率较慢或者水分胁迫严重的情况下, 通过生长环的方法来监测树木的径向生长季节动态存在一定的局限性。比如在水分条件较好的哥伦比亚热带森林中, 高精度生长仪所记录的生长趋势与微树芯法接近, 而在水分胁迫较为严重玻利维亚的干性森林中, 两种方法所获得的径向生长季节动态存在较大的差异[26]。
树木的生长受多种环境因子的共同作用, 物种间的差异也会影响树木生长对气候变化表现出的响应[45]。黄建国等发现北半球针叶植物形成层活动的启动与光周期和温度有关, 随着平均温度的降低和光周期的延长, 木材开始形成的时间延后[3]。厄瓜多尔地区的热带树木黄花风铃木(Tabebuia chrysantha)生长受到水分的限制, 旱季后期树木处于休眠状态, 雨季开始之后形成层开始恢复活动[46]。本研究中楝的径向生长速率与降水、最低温度以及平均温度均呈显著的正相关关系, 其中降水与径向生长速率的相关系数(0.77)最高(图 7)。胡丽凤等研究西双版纳卡斯特森林四个树种径向生长量与气候因子的关系, 结果发现菲律宾朴树(Celtis philippensis)的日径向生长量与最低温度、平均温度和降水呈显著正相关[17], 这与本结果一致。同样地, Kaewmano等发现热带喀斯特森林的红椿(Toona ciliata)的径向生长主要受降水、相对湿度和土壤水分含量的影响, 而热带季节雨林的红椿的径向生长与平均温度和降水均呈正相关关系[47]。西双版纳热带季节雨林多花白头树2020年的径向日生长量与生长季的降水和相对湿度正相关, 而与温度没有显著的相关关系[38]。类似的, 吴丽杰等发现喀斯特森林中楝在2018和2019年的径向生长速率主要与降水和相对湿度呈正相关关系[33], 表明喀斯特生境中楝的径向生长主要受到水分亏缺的限制。本研究中所测得的西双版纳热带植物园内的楝树的年径向生长量为(6.25—7.76)mm, 而在喀斯特森林中楝的年径向生长量小于1 mm[33]。每年的1—3月份, 是西双版纳地区比较干旱的一个时期。2021年的1—3月的降水量为138 mm, 远高于2020年的值(22.6 mm)以及多年平均值(81.2 mm)。2021年相对较高的1—3月降水量可能使得楝在生长季开始时受到水分亏缺的影响相对较小。本研究的结果表明树木生长与风速呈负相关关系, 风速可能是通过间接影响空气湿度, 进而影响树木的生长。
4 结论本研究首次报道了楝的形成层活动季节动态, 发现楝的扩大期细胞开始出现时间为3月1日, 加厚期细胞结束时间为12月11日, 楝的休眠期为2个月左右。生长环和微树芯所监测的树木径向生长的季节动态趋势较为一致, 年径向生长量也接近, 微树芯法能够准确测得楝的木质部生长开始和结束时间。本研究中楝的径向生长与降水、平均温度以及最低温度均呈显著的正相关关系, 其中与降水的相关性最高。热带树木的径向生长和形成层活动季节动态在不同树种以及不同年份可能存在较大的差异, 因此, 未来的研究中多物种以及长时间尺度的监测研究有助于更好的认识热带地区树木的生长规律和调控因子。
| [1] |
Bonan G B. Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science, 2008, 320(5882): 1444-1449. DOI:10.1126/science.1155121 |
| [2] |
Corlett R T. Impacts of warming on tropical lowland rainforests. Trends in Ecology & Evolution, 2011, 26(11): 606-613. |
| [3] |
Huang J G, Ma Q Q, Rossi S, Biondi F, Deslauriers A, Fonti P, Liang E Y, Mäkinen H, Oberhuber W, Rathgeber C B K, Tognetti R, Treml V, Yang B, Zhang J L, Antonucci S, Bergeron Y, Camarero J J, Campelo F, Cˇufar K, Cuny H E, De Luis M, Giovannelli A, Gričar J, Gruber A, Gryc V, Güney A, Guo X L, Huang W, Jyske T, Kašpar J, King G, Krause C, Lemay A, Liu F, Lombardi F, Martinez Del Castillo E, Morin H, Nabais C, Nöjd P, Peters R L, Prislan P, Saracino A, Swidrak I, Vavrčík H, Vieira J, Yu B Y, Zhang S K, Zeng Q, Zhang Y L, Ziaco E. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(34): 20645-20652. DOI:10.1073/pnas.2007058117 |
| [4] |
Deslauriers A, Fonti P, Rossi S, Rathgeber C B K, Gricˇar J. Ecophysiology and plasticity of wood and phloem formation//Amoroso M M, Daniels L D, Baker P J, Camarero J J, eds. Dendroecology: Tree-Ring Analyses Applied to Ecological Studies. Cham: Springer, 2017: 13-33.
|
| [5] |
Lewis S L, Lloyd J, Sitch S, Mitchard E T A, Laurance W F. Changing ecology of tropical forests: evidence and drivers. Annual Review of Ecology, Evolution, and Systematics, 2009, 40: 529-549. DOI:10.1146/annurev.ecolsys.39.110707.173345 |
| [6] |
Fauset S, Baker T R, Lewis S L, Feldpausch T R, Affum-Baffoe K, Foli E G, Hamer K C, Swaine M D. Drought-induced shifts in the floristic and functional composition of tropical forests in Ghana. Ecology Letters, 2012, 15(10): 1120-1129. DOI:10.1111/j.1461-0248.2012.01834.x |
| [7] |
Allen C D, Macalady A K, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears D D, Hogg E H, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J H, Allard G, Running S W, Semerci A, Cobb N. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 2010, 259(4): 660-684. DOI:10.1016/j.foreco.2009.09.001 |
| [8] |
McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams D G, Yepez E A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. New Phytologist, 2008, 178(4): 719-739. DOI:10.1111/j.1469-8137.2008.02436.x |
| [9] |
Soudzilovskaia N A, Elumeeva T G, Onipchenko V G, Shidakov I I, Salpagarova F S, Khubiev A B, Tekeev D K, Cornelissen J H C. Functional traits predict relationship between plant abundance dynamic and long-term climate warming. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(45): 18180-18184. DOI:10.1073/pnas.1310700110 |
| [10] |
Phillips O L, Lewis S L, Baker T R, Chao K J, Higuchi N. The changing Amazon forest. Philosophical Transactions of the Royal Society B: Biological Sciences, 2008, 363(1498): 1819-1827. DOI:10.1098/rstb.2007.0033 |
| [11] |
Laurance W F, Oliveira A A, Laurance S G, Condit R, Nascimento H E M, Sanchez-Thorin A C, Lovejoy T E, Andrade A, D′Angelo S, Ribeiro J E, Dick C W. Pervasive alteration of tree communities in undisturbed Amazonian forests. Nature, 2004, 428(6979): 171-175. DOI:10.1038/nature02383 |
| [12] |
Feeley K J, Joseph Wright S, Nur Supardi M N, Kassim A R, Davies S J. Decelerating growth in tropical forest trees. Ecology Letters, 2007, 10(6): 461-469. DOI:10.1111/j.1461-0248.2007.01033.x |
| [13] |
Clark D A, Piper S C, Keeling C D, Clark D B. Tropical rain forest tree growth and atmospheric carbon dynamics linked to interannual temperature variation during 1984-2000. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(10): 5852-5857. DOI:10.1073/pnas.0935903100 |
| [14] |
Lewis S L, Malhi Y, Phillips O L. Fingerprinting the impacts of global change on tropical forests. Philosophical Transactions of the Royal Society B: Biological Sciences, 2004, 359(1443): 437-462. DOI:10.1098/rstb.2003.1432 |
| [15] |
Bowman D M J S, Williamson G J, Keenan R J, Prior L D. A warmer world will reduce tree growth in evergreen broadleaf forests: evidence from Australian temperate and subtropical eucalypt forests. Global Ecology and Biogeography, 2014, 23(8): 925-934. DOI:10.1111/geb.12171 |
| [16] |
van der Sleen P, Groenendijk P, Vlam M, Anten N P R, Boom A, Bongers F, Pons T L, Terburg G, Zuidema P A. No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience, 2015, 8(1): 24-28. DOI:10.1038/ngeo2313 |
| [17] |
Hu L F, Fan Z X. Stem radial growth in response to microclimate in an Asian tropical dry karst forest. Acta Ecologica Sinica, 2016, 36(5): 401-409. DOI:10.1016/j.chnaes.2016.09.005 |
| [18] |
McMahon S M, Parker G G. A general model of intra-annual tree growth using dendrometer bands. Ecology and Evolution, 2015, 5(2): 243-254. DOI:10.1002/ece3.1117 |
| [19] |
Rossi S, Anfodillo T, Menardi R. Trephor: a new tool for sampling microcores from tree stems. IAWA Journal, 2006, 27(1): 89-97. DOI:10.1163/22941932-90000139 |
| [20] |
Cuny H E, Rathgeber C B K. Xylogenesis: coniferous trees of temperate forests are listening to the climate tale during the growing season but only remember the last words!. Plant Physiology, 2016, 171(1): 306-317. DOI:10.1104/pp.16.00037 |
| [21] |
De Micco V, Carrer M, Rathgeber C B K, Julio Camarero J, Voltas J, Cherubini P, Battipaglia G. From xylogenesis to tree rings: wood traits to investigate tree response to environmental changes. IAWA Journal, 2019, 40(2): 155-182. DOI:10.1163/22941932-40190246 |
| [22] |
Cuny H E, Rathgeber C B K, Frank D, Fonti P, Mäkinen H, Prislan P, Rossi S, Del Castillo E M, Campelo F, Vavrčík H, Camarero J J, Bryukhanova M V, Jyske T, Gričar J, Gryc V, De Luis M, Vieira J, Čufar K, Kirdyanov A V, Oberhuber W, Treml V, Huang J G, Li X X, Swidrak I, Deslauriers A, Liang E, Nöjd P, Gruber A, Nabais C, Morin H, Krause C, King G, Fournier M. Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nature Plants, 2015, 1(11): 15160. DOI:10.1038/nplants.2015.160 |
| [23] |
Huang J G, Guo X L, Rossi S, Zhai L H, Yu B Y, Zhang S K, Zhang M F. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season. Tree Physiology, 2018, 38(8): 1225-1236. DOI:10.1093/treephys/tpy046 |
| [24] |
Pumijumnong N, Danpradit S, Tadang N, Buajan S, Muangsong C. Cambial activity and radial growth dynamics of three tropical tree species at Chang island, Thailand. Journal of Tropical Forest Science, 2019, 31(4): 404-414. DOI:10.26525/jtfs2019.31.4.404 |
| [25] |
Rahman H, Nugroho W D, Nakaba S, Kitin P, Kudo K, Yamagishi Y, Begum S, Marsoem S N, Funada R. Changes in cambial activity are related to precipitation patterns in four tropical hardwood species grown in Indonesia. American Journal of Botany, 2019, 106(6): 760-771. DOI:10.1002/ajb2.1297 |
| [26] |
Mendivelso H A, Camarero J J, Gutiérrez E, Castaño-Naranjo A. Climatic influences on leaf phenology, xylogenesis and radial stem changes at hourly to monthly scales in two tropical dry forests. Agricultural and Forest Meteorology, 2016, 216: 20-36. DOI:10.1016/j.agrformet.2015.09.014 |
| [27] |
Wang K H, Hamzah M Z. Different cambial activities in response to climatic factors of three Malaysian rainforest Shorea species with different stem diameters. Trees, 2018, 32(6): 1519-1530. DOI:10.1007/s00468-018-1730-z |
| [28] |
Locosselli G M. The cambium activity in a changing world. Trees, 2018, 32(1): 1-2. DOI:10.1007/s00468-017-1616-5 |
| [29] |
Hua Z. The tropical flora of southern yunnan, China, and its biogeographic affinities. Annals of the Missouri Botanical Garden, 2008, 95(4): 661-680. DOI:10.3417/2006081 |
| [30] |
Xu C X, Sano M, Yoshimura K, Nakatsuka T. Oxygen isotopes as a valuable tool for measuring annual growth in tropical trees that lack distinct annual rings. Geochemical Journal, 2014, 48(4): 371-378. DOI:10.2343/geochemj.2.0312 |
| [31] |
刘文杰, 李红梅. 我国西双版纳雾资源及其评价. 自然资源学报, 1996, 11(3): 263-267. DOI:10.3321/j.issn:1000-3037.1996.03.011 |
| [32] |
Vlam M, Baker P J, Bunyavejchewin S, Zuidema P A. Temperature and rainfall strongly drive temporal growth variation in Asian tropical forest trees. Oecologia, 2014, 174(4): 1449-1461. DOI:10.1007/s00442-013-2846-x |
| [33] |
吴丽杰, Kaewmano A, 付培立, 王文礼, 范泽鑫. 热带季节性湿润林苦楝(Melia azedarach)径向生长季节动态及其对环境因子的响应. 生态学报, 2020, 40(19): 6831-6840. |
| [34] |
刘佳旭, 李丽娟, 李九一, 王志勇, 陈素景, 张凯. 1954—2014年云南省降水变化特征与潜在的旱涝区域响应. 地球信息科学学报, 2016, 18(8): 1077-1086. |
| [35] |
Anemaet E R, Middleton B A. Dendrometer Bands Made Easy: Using Modified Cable Ties to Measure Incremental Growth of Trees. Applications in Plant Sciences, 2013, 1(9): 1300044. DOI:10.3732/apps.1300044 |
| [36] |
Abernethy K, Bush E R, Forget P M, Mendoza I, Morellato L P C. Current issues in tropical phenology: a synthesis. Biotropica, 2018, 50(3): 477-482. DOI:10.1111/btp.12558 |
| [37] |
Chen X Q, Wang L X, Inouye D. Delayed response of spring phenology to global warming in subtropics and tropics. Agricultural and Forest Meteorology, 2017, 234-235: 222-235. DOI:10.1016/j.agrformet.2017.01.002 |
| [38] |
韦小练, 范泽鑫, Kaewmano A, 林友兴, 陈礼敏, 付培立. 热带季节雨林多花白头树年内径向生长动态及其对环境因子的响应. 应用生态学报, 2021, 32(10): 3567-3575. |
| [39] |
Worbes M. Annual growth rings, rainfall-dependent growth and long-term growth patterns of tropical trees from the caparo forest reserve in venezuela. Journal of Ecology, 1999, 87(3): 391-403. DOI:10.1046/j.1365-2745.1999.00361.x |
| [40] |
Dié A, Kitin P, Kouamé F N, van den Bulcke J, van Acker J, Beeckman H. Fluctuations of cambial activity in relation to precipitation result in annual rings and intra-annual growth zones of xylem and phloem in teak (Tectona grandis) in Ivory Coast. Annals of Botany, 2012, 110(4): 861-873. DOI:10.1093/aob/mcs145 |
| [41] |
周博, 范泽鑫, 杞金华. 哀牢山中山湿性常绿阔叶林木径向生长季节动态及其对气候因子的响应. 生态学报, 2020, 40(5): 1699-1708. |
| [42] |
Anemaet E R, Middleton B A. Dendrometer bands made easy: using modified cable ties to measure incremental growth of trees. Applications in Plant Sciences, 2013, 1(9): 1300044. DOI:10.3732/apps.1300044 |
| [43] |
张军周, 勾晓华, 赵志千, 刘文火, 张芬, 曹宗英, 周非飞. 树轮生态学研究中微树芯石蜡切片制作的方法探讨. 植物生态学报, 2013, 37(10): 972-977. |
| [44] |
Deslauriers A, Morin H, Begin Y. Cellular phenology of annual ring formation of Abies balsamea in the Quebec boreal forest (Canada). Canadian Journal of Forest Research, 2003, 33(2): 190-200. DOI:10.1139/x02-178 |
| [45] |
Anderegg W R L, Schwalm C, Biondi F, Camarero J J, Koch G, Litvak M, Ogle K, Shaw J D, Shevliakova E, Williams A P, Wolf A, Ziaco E, Pacala S. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science, 2015, 349(6247): 528-532. DOI:10.1126/science.aab1833 |
| [46] |
Volland-Voigt F, Bräuning A, Ganzhi O, Peters T, Maza H. Radial stem variations of Tabebuia chrysantha (Bignoniaceae) in different tropical forest ecosystems of southern Ecuador. Trees, 2010, 25(1): 39-48. |
| [47] |
Kaewmano A, Fu P L, Fan Z X, Pumijumnong N, Zuidema P A, Bräuning A. Climatic influences on intra-annual stem radial variations and xylem formation of Toona ciliata at two Asian tropical forest sites with contrasting soil water availability. Agricultural and Forest Meteorology, 2022, 318: 108906. |