 2023, Vol. 43
2023, Vol. 43文章信息
- 孟晓倩, 邵占海, 许丽, 李国雷, 王佳茜
- MENG Xiaoqian, SHAO Zhanhai, XU Li, LI Guolei, WANG Jiaxi
- 苗圃氮加载及水分胁迫对栓皮栎苗木叶片光合氮分配及生物量积累的影响
- The combined effects of nitrogen loading and spring drought on photosynthetic machinery nitrogen distribution and accumulation of biomass of Quercus variabilis seedlings
- 生态学报. 2023, 43(8): 3203-3212
- Acta Ecologica Sinica. 2023, 43(8): 3203-3212
- http://dx.doi.org/10.5846/stxb202203080563
-
文章历史
- 收稿日期: 2022-03-08
- 网络出版日期: 2022-09-20





2. 北京林业大学森林培育与保护教育部重点实验室, 北京 100083;
3. 北京林业大学城乡生态环境北京实验室, 北京 100083;
4. 珍贵落叶树种产业国家创新联盟, 北京 100083;
5. 北京市西山试验林场, 北京 100041
2. Key Laboratory for Silviculture and Conservation, Ministry of Education, Beijing Forestry University, Beijing 100083, China;
3. Beijing Laboratory of Urban and Rural Ecological Environment, Beijing Forestry University, Beijing 100083, China;
4. National Innovation Alliance of Precious Deciduous Tree Species Industry, Beijing 100083, China;
5. Beijing Xishan Forest Farm, Beijing 100041, China
在全球气候变化下, 干旱愈加严重, 世界范围内的树木面临生产力下降甚至死亡的风险[1—2]。在此背景下, 预计我国困难立地造林将面临更严重的极端干旱。幼苗由于根系较浅、无法吸收土壤深层水分、且自身储水能力有限, 面对干旱时其死亡率明显高于其他龄级树木[3], 制约林分更新建成。
氮(N)是决定苗木质量的重要矿质营养之一, 苗圃期科学施N是提高苗木质量与造林表现的核心手段。因此, 研究N加载对苗木干旱生理及生长的影响机制, 能够为干旱立地苗木定向培育提供理论支撑。苗圃木质化期N加载能够使得苗木体内形成N贮存库[4—5]。在早春, 贮存N通过N内循环能够提供植物当前生长所需养分的一半以上[6—7], 如栓皮栎(Quercus variabilis Blume)移栽后第一个月, 新生叶片中70%—98%的N由贮存N转移而来, 且在干旱下贮存N对新组织生长的贡献更大[8]。
光合作用是叶片最为重要的生理生化过程。其中, N在光合作用中发挥着核心作用, 约有一半以上的叶片N分配于光合系统[9], 并且植物生长与光合系统N含量显著相关[10]。光合N利用效率(PNUE)能够表述N含量与光合作用之间的相关性[11], 已经明确, 土壤N含量会影响N向叶片分配, 但目前土壤N含量是否与PNUE呈正相关尚存争议[9, 12—14], 此外, 对于干旱是否以及如何改变苗木PNUE、进而影响生物量积累的问题尚不清楚。
除PNUE外, 叶片光合系统内不同组分的N分配也会影响光合作用的强弱和叶片生理功能[15]。数值模拟结果表明, 不增加N素投入, 只是优化N素在光合蛋白之间的分配, 就可以提高作物光合能力的60%[16]。光合系统内不同组分的N分配受光强、CO2浓度、土壤N含量以及土壤水分状况等因素的影响[17—19]。例如, 黄瓜(Cucumis sativus)幼苗生长环境中的光强由强调弱, 其叶片Rubisco酶比例下降, 更多的N分配至捕光系统以弥补外界环境的局限[20]。此外, 升高CO2浓度可降低Rubisco酶比例, 增加其他组分分配比例[21—22]。还有研究表明, 增加土壤N含量, 库页蓼(Polygonum sachalinense)叶片中的Rubisco酶比例(PR)增加[23], 欧洲赤松(Pinus sylvestris)[24]、火炬松(Pinus taeda)[25]的Rubisco酶含量(NR)增加。干旱胁迫下的研究表明, 水分胁迫会损伤光合系统, 影响同化能力[26], 但对于干旱如何影响叶片光合系统N分配、及其如何受到植物本身N贮存水平的影响尚不清楚。除此之外, 目前还未明确叶片光合系统N分配如何影响生物量。阐明这些问题, 有助于理解N加载对苗木干旱生理及生长的影响机制。
栓皮栎(Q. variabilis)是我国重要的乡土树种, 生态价值极高[27]。但其分布区内存在严重的春季干旱问题, 极大的制约造林初期苗木的生长与存活[28—29]。本研究拟采用施N处理模拟木质化期N加载, 并在翌年苗木移栽后设置水分处理模拟干旱胁迫, 以明确以下两个科学问题:(1)干旱如何影响不同N贮存下苗木叶片N在不同功能组分的分配, 进而影响生物量积累;(2)不同N贮存水平的苗木如何调整PNUE以应对干旱。
1 研究方法 1.1 试验材料和设计试验用种采集于北京市平谷区四座楼林场(117°148′E, 40°282′N;海拔316—467 m)。2013年3月28日, 于北京市中国林业科学研究院温室内(116°14′E, 40°40′N), 挑选催芽后的种子进行播种育苗, 所用基质为消毒后的丹麦品氏草炭土(Pindstrup Plus, 丹麦Pindstrup Mosebrug A/S公司)与珍珠岩的混合物, 比例为3∶1。
2013年4月12日(胚芽萌出)开始施肥, 施肥模型为指数施肥[30—31], 施肥方法为随水施肥。每周施肥1次, 共计16周, 累计施N量50 mg/株(N∶P∶K为50∶26∶33), 同时补施微量元素。每2—3天对育苗托盘称重, 基质含水量低于80%时进行浇水。2013年8月25日至9月4日, 对苗木进行短日照处理以促进其形成顶芽、停止高生长。9月8日至10月6日, 进行5次木质化期N加载处理, 以获得N加载处理:24 mg N/株(24N)、无N加载处理:0 mg N/株(0N)两个不同水平养分库的苗木[32—33]。此外, 所有处理补充等量磷、钾、乙二胺四乙酸二钠(EDTA)、二乙烯三胺五乙酸(DTPA)。木质化期间(2013年9月8日至2013年11月1日), 采用称重法进行浇水, 灌溉参数为70%的基质含水量。
2013年11月1日, 将苗木移至室外炼苗并移入假植坑越冬。2014年3月30日, 在室外遮雨棚下进行移栽, 移栽容器为硬质塑料花盆(佛山市南海区金沙奥东五金塑料有限公司;上径24 cm、下径14 cm、高48 cm), 移栽基质为丹麦品氏草炭土(Pindstrup Plus, 丹麦Pindstrup Mosebrug A/S公司)与水洗沙的混合物(3∶1)。
移栽结束后, 分别对不同N贮存的苗木进行水分处理, 设置水分充足(85%田间持水量)(W)、水分胁迫(D)(40%田间持水量[34])2个水平。试验共4个处理, 每处理90株苗。每3—4天对单株苗木称重并补充水分至各处理水平。水分胁迫期间, 用JL-19空气温湿光记录仪测得温度为14—30℃, 相对湿度为20%—40%, 平均光强为1500 μmol m-2 s-1。
1.2 叶片N、叶绿素和脯氨酸含量测定分别在苗木四个不同生长阶段进行破坏取样:T0(移栽前, 2014年3月28日), 此阶段苗木无生长迹象;T1(移栽后4周, 2014年4月27日), 此时大部分苗木已完成85%的新茎生长, 但叶片生长并未完全;T2(移栽后8周, 2014年5月28日), 苗木新茎与叶片已经完成生长;T3(移栽后12周, 2014年6月25日), 苗木此时没有进一步的高生长。
每阶段每处理随机选取20株苗木破坏取样, 将干净的叶片、新茎、老茎、根分别装入信封, 于65℃烘箱中烘干至恒重, 每5株混合为一个重复, 称其重量, 计算生物量增长量。在T1、T2阶段破坏取样前, 使用CI-3000便携式叶面积仪(LI-COR, Lincoln Nebraska, 美国)测定苗木叶面积(cm2), 计算单株叶面积。另取5株苗木的部分叶片立即放入液氮中并置于-80 ℃超低温冰箱冻存。将烘干叶片混合粉碎, 通过0.25 mm孔径的细筛后用H2SO4-H2O2法消煮, 用smartcham450全自动化学分析仪(意大利AMS Alliance公司)测定叶片N含量。取冻存叶片, 测定叶绿素含量[35]与脯氨酸含量[36]。
1.3 气体交换参数测定在T1、T2生长阶段, 每处理选取3株苗木, 每株苗木选择从顶梢向下的第3—4片健康的功能叶, 以测定光响应曲线和CO2响应曲线。在晴朗天气上午的9:00—11:30, 使用配备红蓝光源的便携式光合仪(LI-6400XT, LI-COR Inc, Lincoln, NE, 美国)进行测定。设置温度为30℃, 测光响应曲线时, 设置12个光合有效辐射梯度:0、20、50、80、100、150、200、400、600、900、1200和1500 μmol m-2 s-1, 每个梯度平衡120—200 s, 控制参比室CO2浓度为400 μmol m-2 s-1;测定CO2响应曲线所设置光强为每株苗木的饱和光强, 利用CO2注入系统控制CO2浓度梯度:400、300、200、100、50、400、600、900、1200和1500 μmol/mol, 每个梯度平衡120—200 s。
1.4 光合系统N分配及PNUE的计算通过拟合光响应曲线, 计算苗木光饱和净光合速率(Amax′, μmol m-2 s-1);通过拟合CO2响应曲线, 计算最大羧化速率Vcmax[37]与最大电子传递速率Jmax[38]。计算Rubisco酶组分(PR)、电子传递组分(PB)以及捕光组分(PL)含量占叶片总N含量比例[39], 方法如下:
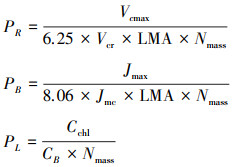
|
其中, LMA(g/m2)为比叶重;Nmass(g/g)为单位干重叶片N含量;CChl(mmol/g)为叶片总叶绿素浓度;Vcr和Jmc为常数, 分别取值为20.78 μmol CO2 g-1 Rubisco s-1、155.65 μmol e- (μmol-1Cyf) s-1[40], CB为叶绿素与N的比值, 通过CB与比叶重拟合的线性公式估算(r2=0.97)[17]:
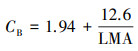
|
计算光合N占叶片总N比例(PP):
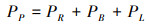
|
将这四个参数分别与叶片N含量(mg)相乘, 获得Rubisco羧化组分N含量(NR), 电子传递组分N含量(NB), 捕光组分N含量(NL)和光合系统N含量(NP)。
计算光合N利用效率(PNUE, μmol g-1 s-1)[30]:
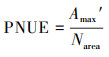
|
式中:Narea为单位面积叶N含量(g/m2)。
1.5 数据分析使用SPSS16.0(美国Statistical Package for Social Science, SPSS公司)对响应曲线进行拟合并对所有数据进行分析, 并用Sigmaplot14.0(美国Systat Software公司)作图。
2 结果分析 2.1 木质化期N加载和翌年春季水分处理对叶面积的影响T1时期, 水分胁迫处理(D)叶面积比水分充足处理(W)显著低27.5%左右(F=7.3, P=0.019);虽然木质化期N加载对叶面积无显著影响, 但N加载处理(24N)比无N加载处理(0N)高9.5%(图 1)。T2时期, 各处理之间的叶面积无显著差异。此外, T1到T2时期的叶面积增量无显著差异, 但就均值而言, N加载处理(24N)比无N加载处理(0N)高131.4%。

|
| 图 1 氮加载对不同水分处理栓皮栎苗木叶面积的影响 Fig. 1 Effect of hardening phase N loading on leaf area of Q. variabilis seedlings under different water conditions T1: 第1时期The frist time of sampling; T2: 第2时期The second time of sampling; 0N-D: 无N加载-干旱胁迫处理The treatment of no nitrogen loading-drought; 0N-W: 无N加载-水分充足处理The treatment of no nitrogen loading-water sufficiency; 24N-D: N加载-干旱胁迫处理The treatment of nitrogen loading-drought; 24N-W: N加载-水分充足处理The treatment of nitrogen loading-water sufficiency; 不同大写字母表示T2时期不同处理间差异显著, 不同小写字母表示T1时期和T2-T1时期不同处理间差异显著(P < 0.05)(数值=均值±标准误) |
T1时期, N加载处理(24N)的叶片N含量比无N加载处理(0N)显著高44.9%(F=16.7, P=0.002), 水分胁迫对叶片N含量影响不显著。T2时期, N加载对叶片N含量无显著影响, 而水分胁迫显著降低叶片N含量(F=9.6, P=0.009)(图 2)。

|
| 图 2 N加载对不同水分处理栓皮栎苗木叶片N含量、叶绿素含量和脯氨酸含量的影响 Fig. 2 Effect of hardening phase N loading on content of N、content of chlorophyll、content of proline in leaf of transplanted Q. variabilis seedlings under different water conditions 不同小写字母表示不同处理间差异显著(P < 0.05) |
T1时期, 水分胁迫处理(D)的叶绿素含量比水分充足处理(W)显著低15.9%(F=9.7, P=0.013)(图 2);木质化期N加载对叶绿素含量无显著影响。在T2时期, N加载和水分胁迫对叶绿素含量无显著影响, 但就均值而言, 水分胁迫处理(D)比水分充足处理(W)低12.4%。
不同于叶绿素, N加载和水分胁迫对T1时期的脯氨酸含量无显著影响, 但水分胁迫处理(D)的均值比水分充足处理(W)高58.3%左右(图 2)。T2时期, 脯氨酸含量受到N加载和水分胁迫交互效应的影响(F=8.7, P=0.016)。0N-D比其他3个处理高43.6%—55.7%(图 2)。
2.3 木质化期N加载和翌年春季水分处理对叶片光合N分配比例、含量和PNUE的影响在T1时期, PP、PR、PB均受N加载和水分胁迫交互作用影响(Fpp=90.1, Ppp < 0.001; FPR=53.2, PPR < 0.001; FPB=124.1, PPB < 0.001), 24N-W的PP、PR分别较24N-D高141.2%、184.3%, 而0N-W的PP、PR分别比0N-D高6.8%、5.2%。24N-W的PB比24N-D高128.3%, 0N-W的PB则比0N-D高低28.3%。N加载(FPL=72.1, PPL < 0.001)和翌年春水分胁迫(FPL=56.5, PPL < 0.001)显著降低PL, N加载处理(24N)的PL较无N加载处理(0N)低16.3%;水分胁迫处理(D)较水分充足处理(W)低14.2%。到了T2时期, PP、PR、PB仍受N加载和水分胁迫交互作用影响(FPP=147.8, PPP < 0.001; FPR=600.9, PPP < 0.001; FPB=57.2, PPP < 0.001), PL不受N加载和水分胁迫的影响。24N-W的PP、PR、PB分别比24N-D高130.0%、175.1%、128.3%;0N-W的PP、PR、PB则分别比0N-D低1.4%、22.3%、28.3%(图 3)。

|
| 图 3 N加载对不同水分处理栓皮栎苗木叶片N分配比例的影响 Fig. 3 Effect of hardening phase N loading on N allocation proportion of different functional components in photosynthetic machinery of Q. variabilis seedlings under different water conditions |
T1时期, NP、NR、NB受N加载和水分处理交互作用的影响(FNP=128.3, PNP < 0.001; FNR=102.9, PNR < 0.001; FNB=98.6, PNB < 0.001)(图 4)。24N-W的NP较24N-D高173.9%左右, 0N-W的NP则仅高出0N-D约33.0%;24N-W的NR比24N-D高222.5%, 0N-W的NR仅比0N-D高30.9%;24N-W的NB较24N-D高约170.0%, 0N-W的NB则仅比0N-D高31.3%。NL则分别受木质化N加载(FNL=53.3, PNL < 0.001)和水分处理(FNL=196.0, PNL < 0.001)均显著影响NL:高N处理(24N)较低N处理(0N)高23.60%;水分胁迫处理(D)较水分充足处理(W)低30.29%。到了T2时期, NP、NR、NB仍受N加载和水分处理交互作用的影响(FNP=102.9, PNP < 0.001; FNR=142.2, PNR < 0.001; FNB=152.1, PNB < 0.001)。24N-W的NP较24N-D约高198.6%, 0N-W的NP则仅高出0N-D约31.9%;24N-W的NR比24N-D高257.1%, 0N-W的NR仅比0N-D高24.3%;24N-W的NB较24N-D高约196.3%, 0N-W的NB则仅高出0N-D约13.8%。NL则受N加载(FNL=6.8, PNL=0.035)和水分处理(FNL=9.6, PNL=0.017)的显著影响:高N处理(24N)较低N处理(0N)高54.8%;干旱处理(D)较水分充足处理(W)低41.7%。

|
| 图 4 N加载对不同水分处理栓皮栎苗木叶片N分配含量的影响 Fig. 4 Effect of hardening phase N loading on N allocation content of different functional components in photosynthetic machinery of Q. variabilis seedlings under different water conditions |
T1时期, 水分胁迫处理(D)的PNUE较水分充足处理(W)显著降低了45.6%(F=14.9, P=0.005);N加载对叶片PNUE无显著影响。T2时期, PNUE不受木质化期N加载和翌年春季水分处理的影响(图 5)。

|
| 图 5 N加载对不同水分处理下栓皮栎苗木叶片光合N利用效率的影响 Fig. 5 Effect of hardening phase N loading on PNUE of Q. variabilis seedlings under different water conditions |
T1的叶片N在不同功能库之间的含量和分配比例与苗木生物量积累不相关;到了T2, 光合N含量(NP)与T2—T3的根、老茎、新茎、叶片的生物量增长量均正相关, 非光合N含量(NNP)与该阶段新茎增长呈负相关(表 1)。此外, T1时期光合系统内的N分配也与苗木生物量积累不相关。随着叶片的发育, T2的NR和NB均与T2—T3苗木各器官的生物量增长正相关, NL与根的生物量生长不相关, 而与老茎、新茎、叶片生物量生长呈正相关(表 1)。
| T2至T3时期 T2—T3 |
Rubisco羧化组分N含量 NR/mg |
电子传递组分N含量 NB/mg |
捕光组分N含量 NL/mg |
光合系统N含量 NP/mg |
非光合系统N含量 NNP/mg |
脯氨酸含量 Proline content/ (μg/g) |
| 根生物量积累量 Biomass accumulation of root |
0.665** | 0.728* | — | 0.691* | — | — |
| 老茎生物量积累量 Biomass accumulation of old stem |
0.775* | 0.833* | 0.758** | 0.809** | — | — |
| 新茎生物量积累量 Biomass accumulation of new stem |
0.851** | 0.839** | 0.823** | 0.864** | -0.650* | — |
| 叶片生物量积累量 Biomass accumulation of leaf |
0.652* | 0.658* | 0.843** | 0.690* | — | -0.586* |
| NR: Rubisco羧化组分N含量allocation content of N in Rubisco components; NB: 电子传递组分N含量allocation content of N in electron transport components; NL: 捕光组分N含量allocation content of N in light-harvesting components; NP: 光合系统N含量allocation content of N in the photosynthetic system; NNP: 非光合系统N含量allocation content of N in the non-photosynthetic system; —: 无显著差异;*: 5%水平显著差异;**: 1%水平显著差异 | ||||||
T1时期的脯氨酸含量与苗木生物量积累无显著相关性, T2时期, 叶片脯氨酸含量与叶片生长呈负相关(R2=-0.586, P < 0.05)。叶片完成发育后(T2), 叶片脯氨酸含量与捕光组分N含量显著相关(R2=-0.668, P < 0.05), 与光合系统内部各组分N含量不相关。
2.5 PNUE与叶片N分配含量、比例的相关性分析T1时期, PNUE与NB、PL显著正相关。T2时期, PNUE与NR、NB、NL、PR和PL均呈显著正相关关系(表 2)。
| 时期 Time |
Rubisco羧化 组分N含量 NR/mg |
电子传递 组分N含量 NB/mg |
捕光组分 N含量 NL/mg |
Rubisco羧化组 分N分配比例 PR/(g/g) |
电子传递组分 N分配比例 PB/(g/g) |
捕光组分 N分配比例 PL/(g/g) |
|
| 光合N利用效率 | T1 | — | 0.672* | — | — | — | 0.632* |
| PNUE | T2 | 0.668* | 0.629* | 0.747* | 0.608* | — | 0.653* |
| PR: Rubisco羧化组分N分配比例N allocation proportion of Rubisco components; PB: 电子传递组分N分配比例N allocation proportion of electron transport components; PL: 捕光组分N分配比例N allocation proportion of light-harvesting components; PNUE: 光合N利用效率Photosynthetic N-use efficiency. | |||||||
研究表明, 叶片发育完成后, 干旱抑制N向光合系统分配, 苗圃N加载能够缓解这种抑制作用。这主要是由于N加载苗木在干旱下向Rubisco分配了更多的N(图 4), 同时, N加载能够促进叶片扩张(图 1)、提高叶片N含量(图 2), 且在T2时期, 光合N含量越高, 后续(T2—T3)的生物量积累量越大(表 1), 因此N加载促进N向光合组分分配, 从而在一定程度上抵消水分胁迫对生物量积累的抑制。以往研究表明, 由于干旱不限制N的再利用, 所以无论是否干旱, 高N贮存苗木均比低N贮存苗木具有更高的生长量[8]。刘涛[13]的研究表明, 施N能够提高油菜的叶片干重和叶面积。另有研究表明, 拟南芥RAF1与LS在烟草中的共表达提高了Rubisco含量和光合作用, 从而促进了植物生长[41]。Cao等[42]针对水稻的研究也发现了类似的结论。本研究揭示了在干旱胁迫下, N加载苗木能够向叶片的光合功能组分中分配更多的N, 从而促进苗木进行生物量积累。
本研究还发现, 叶片生长与脯氨酸含量之间存在权衡关系。叶片完成发育后, 脯氨酸含量越高, 后续叶片生物量积累越少(表 1)。干旱促进叶片形成脯氨酸, 而苗圃N加载能够抑制叶片脯氨酸含量的增加(图 2), 同时, N加载苗木的叶片生长较大(图 1), 说明苗圃N加载促进干旱胁迫下的苗木对生长的投资, 而无N加载的苗木更倾向于投资于防御。此外, 干旱胁迫抑制N向捕光组分分配(图 4)、促进向脯氨酸分配(图 2), 而N加载能够抑制脯氨酸增加, 维持捕光组分较高的N含量。有研究发现, 在轻度水分胁迫下, 水稻通过增加游离氨基酸和脯氨酸进行渗透调节以保护光合系统, 从而维持高光合效率以及高生产力[43], 这与本研究的对象所受到的水分胁迫程度不同, 因而结论也有差异。本研究的发现与Zhong等[44]的研究一致, 该研究发现, 干旱导致低N处理水稻光捕获组分的N分配减少15%。本研究进一步说明了苗圃N加载促进苗木在干旱下将更多的N分配于光合系统, 以促进生物量积累;无N加载苗木则偏好采取保守的策略, 提高脯氨酸水平, 但向光合系统投资更少。
3.2 光合N分配受干旱影响而改变以维持较高的PNUE本研究还发现, 栓皮栎苗木通过调整不同功能组分光合N分配以维持干旱下较高的PNUE。叶片发育完成后, 干旱胁迫促进无N加载苗木将N向羧化组分分配, 而N加载抑制叶片N向羧化系统以及电子传递系统分配。但该时期干旱胁迫下的PNUE较T1时期有所上升(图 5), 而PNUE的提高与叶片光合系统不同功能组分的N分配比例及含量密切相关(表 2)。也有研究表明, N在光合系统不同组分分配的差异是影响冬油菜苗期叶片PNUE的重要原因[45]。核酮糖-1, 5-二磷酸羧化酶/加氧酶(RuBPCase)决定了羧化速率大小, 是光合能力的限制因子[37, 46], 此外, RuBPCase还具有N贮存功能[47], 研究显示, N充裕的条件下, 叶片向RuBPCase投入的比例降低[45], 且该酶活性也较低, 即在满足羧化功能外, 额外的RuBPCase以贮存N形式存在。本研究认为, 干旱下, 由于N吸收受抑制[8]导致N水平匮乏, 使得RuBPCase无法满足贮存功能需求。干旱下低N贮存苗木体内N较为匮乏, 推测此时向羧化系统投入更多N以满足羧化作用基本需求, 而高N贮存苗木N水平较高, 因此向羧化系统分配的N也较高(图 3, 图 4), 推测已能够满足羧化需求, 受到抑制可能由于RuBPCase无法履行贮存功能。此外, 干旱所造成的气孔限制导致CO2进入气孔的扩散阻力增大, 胞间CO2浓度减少, 减少了光合作用所需的底物浓度[48], 这也可能是干旱下高N贮存苗木向羧化系统和电子传递组分分配N比例下降的原因之一。因此, 本研究在前人研究基础上进一步表明, 干旱胁迫影响叶片N向羧化组分、电子传递组分分配, 从而提高PNUE。
4 结论本研究表明, 叶片发育完成后, 干旱抑制N向光合系统的分配, 但苗圃N加载能够缓解干旱对光合N的抑制作用, 从而在一定程度上抵消水分胁迫对生物量积累的抑制;无N加载苗木则偏好采取保守的策略, 向光合系统投资更少, 而提高脯氨酸水平。低N贮存的苗木在遭受干旱后将N向羧化组分分配, 而高N贮存的苗木遭遇干旱后则显著抑制叶片将N向羧化系统、以及电子传递系统分配, 捕光组分N的分配则不受植物体内N贮存或外部水分状况的影响, 栓皮栎苗木通过调整不同功能组分光合N含量和分配以维持干旱下较高的PNUE。本研究结果对于干旱立地下森林植被恢复以及造林苗木选择、培育具有生态学理论和应用意义。
| [1] |
Brodribb T J, Powers J, Cochard H, Choat B. Hanging by a thread? Forests and drought. Science, 2020, 368(6488): 261-266. DOI:10.1126/science.aat7631 |
| [2] |
Choat B, Brodribb T J, Brodersen C R, Duursma R A, López R, Medlyn B E. Triggers of tree mortality under drought. Nature, 2018, 558(7711): 531-539. DOI:10.1038/s41586-018-0240-x |
| [3] |
谭凤森, 宋慧清, 李忠国, 张启伟, 朱师丹. 桂西南喀斯特季雨林木本植物的水力安全. 植物生态学报, 2019, 43(3): 227-237. |
| [4] |
李国雷, 刘勇, 祝燕. 秋季施肥调控苗木质量研究评述. 林业科学, 2011, 47(11): 166-171. DOI:10.11707/j.1001-7488.20111127 |
| [5] |
Jacobs D F, Salifu K F, Seifert J R. Growth and nutritional response of hardwood seedlings to controlled-release fertilization at outplanting. Forest Ecology and Management, 2005, 214(1/3): 28-39. |
| [6] |
Millard P, Grelet G A. Nitrogen storage and remobilization by trees: ecophysiological relevance in a changing world. Tree Physiology, 2010, 30(9): 1083-1095. DOI:10.1093/treephys/tpq042 |
| [7] |
Villar-Salvador P, Uscola M, Jacobs D F. The role of stored carbohydrates and nitrogen in the growth and stress tolerance of planted forest trees. New Forests, 2015, 46(5/6): 813-839. |
| [8] |
Wang J X, Villar-Salvador P, Li G L, Liu Y. Moderate water stress does not inhibit nitrogen remobilization, allowing fast growth in high nitrogen content Quercus variabilis seedlings under dry conditions. Tree Physiology, 2019, 39(4): 650-660. DOI:10.1093/treephys/tpy130 |
| [9] |
Mu X H, Chen Q W, Chen F J, Yuan L X, Mi G H. Within-leaf nitrogen allocation in adaptation to low nitrogen supply in Maize during grain-filling stage. Frontiers in Plant Science, 2016, 7: 699. |
| [10] |
Feng Y L, Lei Y B, Wang R F, Callaway R M, Valiente-Banuet A, Inderjit, Li Y P, Zheng Y L. Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(6): 1853-1856. DOI:10.1073/pnas.0808434106 |
| [11] |
Hikosaka K. Optimal nitrogen distribution within a leaf canopy under direct and diffuse light. Plant, Cell & Environment, 2014, 37(9): 2077-2085. |
| [12] |
Ibrahim M H, Jaafar H Z E, Rahmat A, Rahman Z A. The relationship between phenolics and flavonoids production with total non structural carbohydrate and photosynthetic rate in Labisia pumila Benth. under high CO2 and nitrogen fertilization. Molecules, 2011, 16(1): 162-174. |
| [13] |
刘涛. 氮供应对油菜叶片氮形态分配与光合氮利用效率的影响机制[D]. 武汉: 华中农业大学, 2018.
|
| [14] |
Zhang L L, Chen X Y, Wen D Z. Interactive effects of rising CO2 and elevated nitrogen and phosphorus on nitrogen allocation in invasive weeds Mikania micrantha and Chromolaena odorata. Biological Invasions, 2016, 18(5): 1391-1407. DOI:10.1007/s10530-016-1089-2 |
| [15] |
Hikosaka K. Interspecific difference in the photosynthesis-nitrogen relationship: patterns, physiological causes, and ecological importance. Journal of Plant Research, 2004, 117(6): 481-494. DOI:10.1007/s10265-004-0174-2 |
| [16] |
Zhu X G, de Sturler E, Long S P. Optimizing the distribution of resources between enzymes of carbon metabolism can dramatically increase photosynthetic rate: a numerical simulation using an evolutionary algorithm. Plant Physiology, 2007, 145(2): 513-526. DOI:10.1104/pp.107.103713 |
| [17] |
史作民, 唐敬超, 程瑞梅, 罗达, 刘世荣. 植物叶片氮分配及其影响因子研究进展. 生态学报, 2015, 35(18): 5909-5919. |
| [18] |
唐敬超, 史作民, 罗达, 刘世荣. 遮荫处理对灰木莲幼苗叶片光合氮利用效率的影响. 生态学报, 2017, 37(22): 7493-7502. |
| [19] |
乌佳美, 唐敬超, 史作民, 冯秋红, 赵广东, 刘顺, 曹向文. 巴郎山糙皮桦叶片光合氮利用效率的海拔响应. 应用生态学报, 2019, 30(3): 751-758. |
| [20] |
Trouwborst G, Hogewoning S W, Harbinson J, vanIeperen W. Photosynthetic acclimation in relation to nitrogen allocation in cucumber leaves in response to changes in irradiance. Physiologia Plantarum, 2011, 142(2): 157-169. DOI:10.1111/j.1399-3054.2011.01456.x |
| [21] |
孙谷畴, 赵平, 曾小平, 彭少麟. 大气CO2浓度升高对香蕉光合作用及光合碳循环过程中叶氮分配的影响. 应用生态学报, 2001, 12(3): 429-434. DOI:10.3321/j.issn:1001-9332.2001.03.027 |
| [22] |
Crous K Y, Walters M B, Ellsworth D S. Elevated CO2 concentration affects leaf photosynthesis-nitrogen relationships in Pinus taeda over nine years in FACE. Tree Physiology, 2008, 28(4): 607-614. DOI:10.1093/treephys/28.4.607 |
| [23] |
Akita R, Kamiyama C, Hikosaka K. Polygonum sachalinense alters the balance between capacities of regeneration and carboxylation of ribulose-1, 5-bisphosphate in response to growth CO2 increment but not the nitrogen allocation within the photosynthetic apparatus. Physiologia Plantarum, 2012, 146(4): 404-412. DOI:10.1111/j.1399-3054.2012.01631.x |
| [24] |
Warren C R, Dreyer E, Adams M A. Photosynthesis-Rubisco relationships in foliage of Pinus sylvestris in response to nitrogen supply and the proposed role of Rubiscoand amino acids as nitrogen stores. Trees, 2003, 17(4): 359-366. DOI:10.1007/s00468-003-0246-2 |
| [25] |
Tissue D T, Thomas R B, Strain B R. Long-term effects of elevated CO2 and nutrients on photosynthesis and Rubisco in loblolly pine seedlings. Plant, Cell & Environment, 1993, 16(7): 859-865. |
| [26] |
葛晋纲, 蔡庆生, 周兴元, 宋刚. 土壤干旱胁迫对2种不同光合类型草坪草的光合特性和水分利用率的影响. 草业科学, 2005, 22(4): 103-107. DOI:10.3969/j.issn.1001-0629.2005.04.029 |
| [27] |
李文英, 王冰, 黎祜琛. 栎类树种的生态效益和经济价值及其资源保护对策. 林业科技通讯, 2001(8): 13-15. DOI:10.3969/j.issn.1671-4938.2001.08.004 |
| [28] |
谢清霞, 李刚, 袁晨, 谷晓平. 基于CI指数的西南地区1961-2012年春季干旱分布特征. 沙漠与绿洲气象, 2016, 10(4): 53-58. |
| [29] |
刘秀红, 李智才, 刘秀春, 宋燕, 张国勇, 李婧婧. 山西春季干旱的特征及成因分析. 干旱区资源与环境, 2011, 25(9): 156-160. |
| [30] |
Timmer V R, Armstrong G. Growth and nutrition of containerized Pinusresinosa at exponentially increasing nutrient additions. Canadian Journal of Forest Research, 1987, 17(7): 644-647. |
| [31] |
Ingestad T, Lund A B. Theory and techniques for steady state mineral nutrition and growth of plants. Scandinavian Journal of Forest Research, 1986, 1(1/4): 439-453. |
| [32] |
Li G L, Zhu Y, Liu Y, Wang J X, Liu J J, Dumroese R K. Combined effects of pre-hardening and fall fertilization on nitrogen translocation and storage in Quercus variabilis seedlings. European Journal of Forest Research, 2014, 133(6): 983-992. |
| [33] |
Wang J X, Yu H Q, Li G L, Zhang F. Growth and nutrient dynamics of transplanted Quercus variabilis seedlings as influenced by pre-hardening and fall fertilization. Silva Fennica, 2016, 50(2): 1475. |
| [34] |
杨自立, 马履一, 贾忠奎, 王梓, 公宁宁. 栓皮栎播种苗年生长动态. 东北林业大学学报, 2012, 40(5): 9-12. |
| [35] |
徐敏, 刘君, 阿衣古力·阿布都瓦依提. 植物生理实验教学中叶绿素提取方法比较. 实验科学与技术, 2018, 16(4): 129-133. |
| [36] |
赵天然. 水分和盐分胁迫对流苏树的生长及光合生理特性的影响[D]. 山东: 山东农业大学, 2021.
|
| [37] |
Farquhar G D, von Caemmerer S, Berry J A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta, 1980, 149(1): 78-90. |
| [38] |
Loustau D, Brahim M B, Gaudillère J P, Dreyer E. Photosynthetic responses to phosphorus nutrition in two-year-old maritime pine seedlings. Tree Physiology, 1999, 19(11): 707-715. |
| [39] |
Niinemets V, Tenhunen J D. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum. Plant, Cell & Environment, 1997, 20(7): 845-866. |
| [40] |
刘涛, 鲁剑巍, 任涛, 汪威, 王振, 王少华. 适宜氮水平下冬油菜苗期不同叶位叶片光合氮分配特征. 中国农业科学, 2016, 49(18): 3532-3541. |
| [41] |
Whitney S M, Birch R, Kelso C, Beck J L, Kapralov M V. Improving recombinant Rubisco biogenesis, plant photosynthesis and growth by coexpressing its ancillary RAF1 chaperone. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(11): 3564-3569. |
| [42] |
Cao X C, Zhong C, Zhu C Q, Zhang J H, Zhu L F, Wu L H, Jin Q Y. Variability of leaf photosynthetic characteristics in rice and its relationship with resistance to water stress under different nitrogen nutrition regimes. Physiologia Plantarum, 2019, 167(4): 613-627. |
| [43] |
Zhong C, Cao X C, Bai Z G, Zhang J H, Zhu L F, Huang J L, Jin Q Y. Nitrogen metabolism correlates with the acclimation of photosynthesis to short-term water stress in rice (Oryza sativa L.). Plant Physiology and Biochemistry, 2018, 125: 52-62. |
| [44] |
Zhong C, Jian S F, Huang J, Jin Q Y, Cao X C. Trade-off of within-leaf nitrogen allocation between photosynthetic nitrogen-use efficiency and water deficit stress acclimation in rice (Oryza sativa L.). Plant Physiology and Biochemistry, 2019, 135: 41-50. |
| [45] |
刘涛, 鲁剑巍, 任涛, 李小坤, 丛日环. 不同氮水平下冬油菜光合氮利用效率与光合器官氮分配的关系. 植物营养与肥料学报, 2016, 22(2): 518-524. |
| [46] |
Hikosaka K, Terashima I. Nitrogen partitioning among photosynthetic components and its consequence in sun and shade plants. Functional Ecology, 1996, 10(3): 335-343. |
| [47] |
Warren C R, Adams M A. Evergreen trees do not maximize instantaneous photosynthesis. Trends in Plant Science, 2004, 9(6): 270-274. |
| [48] |
Gholipoor M, Choudhary S, Sinclair T R, Messina C D, Cooper M. Transpiration response of maize hybrids to atmospheric vapour pressure deficit. Journal of Agronomy and Crop Science, 2013, 199(3): 155-160. |