 2023, Vol. 43
2023, Vol. 43文章信息
- 杨熙来, 朱榴骏, 冯兆忠
- YANG Xilai, ZHU Liujun, FENG Zhaozhong
- 臭氧胁迫冬小麦叶片高光谱特征和叶绿素含量估算
- Hyperspectral characteristics and chlorophyll content estimation of winter wheat under ozone stress
- 生态学报. 2023, 43(8): 3213-3223
- Acta Ecologica Sinica. 2023, 43(8): 3213-3223
- http://dx.doi.org/10.5846/stxb202201200196
-
文章历史
- 收稿日期: 2022-01-20
- 网络出版日期: 2022-12-29

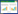




2. 河海大学长江保护与绿色发展研究院, 南京 210024
2. Yangtze Institute for Conservation and Development, Hohai University, Nanjing 210024, China
近地面臭氧在中国乃至全球的污染形势都十分严峻。臭氧不仅危害人体, 致使人体感染呼吸道疾病, 还会影响地面作物正常的生理生长活动, 进而影响作物的产量[1]。Feng等[2]发现臭氧污染导致东亚地区每年作物减产损失合计约630亿美元, 其中以中国地区的小麦、杂交稻、常规稻和玉米减产最为严重, 减产率分别为32.6%、29.3%、12.9%和8.6%。Kaliakatsou等[3]发现AOT40(白天小时O3浓度值超过40 nmol/mol的累积值)增加10%, 小麦产量下降0.54%。Ren等[4]评估了长三角地区臭氧污染对稻麦的影响研究, 结果显示2014—2017年, 冬小麦平均产量损失约2025吨, 占实际产量的20.4%, 平均经济损失约7.36亿美元。为保障国家粮食安全, 作物臭氧胁迫监测研究刻不容缓。现有定性方法是根据城市空气质量监测站获取的地表臭氧浓度数据是否超过敏感植物受害阈值[5](AOT40)判定臭氧胁迫。随着遥感技术的迅速发展, 大面积臭氧胁迫定量监测将成为可能, 其具有易获得、无损、数据更新速度快等优点。
目前, 光谱遥感监测植物生理参量已经被广泛应用于生物、非生物胁迫领域[6—14], 但是应用于臭氧胁迫的研究较少。程小云等[15]利用O3胁迫下27种木本植物的光谱特征构建叶绿素含量与红边位置反射率混合模型;Gab等[16]研究发现近红外波段能够有效的监测森林树木叶片的臭氧胁迫;Marchica等[17]利用遥感光谱技术对臭氧胁迫下鼠尾草(Salvia japonica Thunb)的光谱特征进行研究, 发现即使没有可见症状也能通过光谱判别鼠尾草是否受到臭氧胁迫。Calzone等[18]对不同臭氧浓度梯度胁迫下的石榴(Punica granatum)进行光谱分析, 发现光谱可以快速、无损的评估早期植物臭氧胁迫。而冬小麦臭氧胁迫的光谱研究更是匮乏。Chi等[19]分析了不同生育期四个冬小麦品种在O3-FACE平台下的胁迫影响, 筛选出与臭氧胁迫相关性较好的遥感植被指数, 但是没有研究光谱特征参数与冬小麦生理参数的相关性, 并且其建模方法较为单一, 无法筛选出最佳估测模型。
为此, 本研究利用自由式臭氧浓度增加系统(O3-FACE)平台, 分析冬小麦三个生育期(拔节期、开花期和灌浆期)叶片光谱特征和叶绿素含量的变化规律。通过分析光谱参数与叶绿素含量的相关性, 筛选出臭氧胁迫下与叶绿素含量相关性较高的光谱指标, 并以叶绿素数据为因变量, 构建三种叶绿素含量估算模型, 评价其可行性。筛选出能够准确、快速反演冬小麦叶绿素含量的模型, 为实际生产过程中冬小麦受到臭氧胁迫的动态监测提供理论依据。
1 材料与方法 1.1 研究区概况实验区位于江苏省扬州市江都区南京信息工程大学扬州绿色农业研究与示范基地(32°25′12″N, 119°43′48″E), 属亚热带季风气候, 2009—2018年平均气温16.2℃, 年平均降水量1131.3 mm, 年日照时间1936.1 h, 年无霜期>290 d, 当地实行长期的稻麦轮作体系, 6月—11月种植水稻, 11月—第二年6月种植冬小麦, 属于典型的农田生态系统[20]。
1.2 实验设计实验于2021年3月—6月开展, 供试冬小麦品种为农麦88。2020年11月播种, 2021年6月收获。O3-FACE是最接近自然的一种模拟大气气体浓度增加对生态系统影响的研究平台, 已广泛应用于各种生态系统[21]。该平台分为自然对照处理(A-O3)和臭氧胁迫处理(E-O3), 每种处理各有四个重复, 共8个处理区, 每个处理区有10个亚区, 单个亚区的面积为3 m×3 m, 本研究利用其中一个亚区进行实验。实验期间, 该系统每天从8点到18点持续提供臭氧熏蒸。为验证小麦不同生长阶段的臭氧胁迫, 分别在拔节期(4月17日)、开花期(5月9日)、灌浆期(5月21日)测定, 每次随机从八个处理区的实验亚区中采10片冬小麦叶片进行试验。共有对照光谱样本134个, 臭氧胁迫光谱样本131个。由于拔节期臭氧对冬小麦几乎无影响, 本实验使用开花期和灌浆期共91个光谱样本建模。叶绿素数据数量与光谱数据数量一致。
1.3 光谱采集与预处理实验采用SOC710便携式成像光谱仪采集冬小麦叶片光谱反射率数据, 其测量光谱范围为400—1000 nm, 光谱分辨率为4.6875 nm, 共有波段128个, 包含常用的可见光波段以及一部分近红外波段。光谱观测均选择晴朗无云或少云的天气, 观测时间为叶绿素含量观测日的上午10点到下午15点。观测时利用标准灰板进行校正, 以消除太阳偏射以及太阳光强不均匀带来的误差。利用光谱数据分析软件SRAnal710对原始光谱数据(DN值)进行辐射定标, 得到各叶片光谱反射率图像。利用ENVI软件从叶片图像上随机提取10—20个点的光谱反射率, 通过取均值得到每片叶片的光谱反射率。利用MATLAB软件统计分析, 通过Origin软件作图。图表中数据为平均值±标准差。
1.4 叶绿素含量的测定与预处理在测量叶片光谱时, 同步使用便携式SPAD-502叶绿素仪测定冬小麦叶绿素含量(SPAD)。类似于光谱观测, 随机选取叶片不同方向的10个点进行测定SPAD, 最终取平均值代表该叶片的叶绿素含量。利用JMP Pro16软件对数据进行显著性分析, 利用Origin软件作图。
1.5 光谱指标的计算与筛选本文选取原始光谱反射率、常用遥感植被指数以及常用的基于光谱位置、光谱面积等光谱特征参数, 作为估算冬小麦叶片叶绿素的光谱参数。采用式(1)对原始反射率进行一阶微分计算:
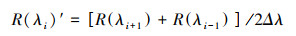
|
(1) |
式中:λi为波段, R(λi)为波段λi对应的光谱反射率, Δλ为相邻波段间隔, R(λi)′为反射率在波段λi处的一阶微分值。
根据植被的光谱特性, 将可见光和近红外波段进行组合, 形成了各种遥感植被指数。本文在前人做的研究工作中找出并计算25种常见的植被指数[22—27](表 1)和17种常用的光谱特征参数[9—10, 28](表 2)。利用SPSS 21软件将这两类光谱参数分别与SPAD值进行相关性分析, 并利用Origin软件作图。
| 遥感植被指数 Remote sensing vegetation indices |
名称 Name |
计算公式 Calculation formula |
| GI | 绿度指数 | R554/R667 |
| ARVI | 大气阻抗植被指数 | (R810-(2R680-R480))/(R810+(2R680-R480)) |
| VARI | 可视化气压阻抗指数 | (R555-R680)/(R555+R680-R480) |
| NDVI705 | 归一化植被指数 | (R750-R705)/(R750+R705) |
| MSR705 | 改进红边比值指数 | (R705-R445)/(R705+R445) |
| NDVI670 | 归一化植被指数 | (R800/R670)/(R800+R670) |
| CI | 叶绿素指数 | (R750/R705)-1 |
| PSRI | 植被衰老反射指数 | (R680-R500)/R750 |
| RGI | 相对绿色指数 | R690/R550 |
| EVI | 增强植被指数 | (2.5×(R782-R675))/(R782+6×R675-7.5×R445+1) |
| RVI | 比值植被指数 | R800/R760 |
| MCARI | 改进叶绿素吸收比指数 | R700-R670-0.2×(R700-R550)×(R700/R670) |
| TCARI | 增强叶绿素吸收比指数 | 3×(R700-R670)-0.2×(R700-R550)×(R700/R670) |
| CIrededge | 土壤调节植被指数 | (R700-R800)/(R690+R720)-1 |
| PRI | 光化学反射指数 | (R570-R531)/(R570+R531) |
| CRI | 类胡萝卜素反射指数 | (1/R510)-(1/R550) |
| ANT | 花青素反射指数 | (1/R550)-(1/R700) |
| NDVI | 归一化植被指数 | (R750-R550)/(R750+R550) |
| mND705 | 改进红边归一化植被指数 | (R750-R705)/(R750+R705-2×R445) |
| SR705 | 比值植被指数 | R750/R705 |
| DD | 双差分指数 | (R750-R720)/(R800+R680) |
| GM | 相对深绿指数 | R750/R700 |
| R800 | 色素含量指数 | R800-R550 |
| NDRE | 归一化红边指数 | (R790-R720)/(R790+R720) |
| NPCI | 叶绿素色素比率指数 | (R650-R420)/(R650+R420) |
| Ri为i波段的光谱反射率 | ||
| 高光谱特征参数 Hyperspectral characteristics parameters |
名称 Name |
说明 Illustration |
| AR | 红光反射率平均值 | 光谱段600—700 nm的平均值 |
| Rg | 绿峰反射率 | 光谱段510—560 nm内最大光谱反射率值 |
| λg | 绿峰位置 | 光谱段510—560 nm内最大光谱反射率对应波长位置 |
| Rr | 红谷反射率 | 光谱段650—690 nm内最小的光谱反射率 |
| Rg/Rr | 绿峰和红谷反射率的比值 | |
| Db | 蓝边幅值 | 光谱段490—530 nm内一阶微分的最大值 |
| λb | 蓝边位置 | 光谱段490—530 nm内一阶微分最大值对应的波长位置 |
| Dy | 黄边幅值 | 光谱段560—640 nm内一阶微分的最大值 |
| λy | 黄边位置 | 光谱段560—640 nm内一阶微分最大值对应的波长位置 |
| Dr | 红边幅值 | 光谱段680—760 nm内一阶微分的最大值 |
| λr | 红边位置 | 光谱段680—760 nm内一阶微分最大值对应的波长位置 |
| SDb | 蓝边面积 | 蓝边范围内一阶微分波段值的总和 |
| SDy | 黄边面积 | 黄边范围内一阶微分波段值的总和 |
| SDr | 红边面积 | 红边范围内一阶微分波段值的总和 |
| SDg | 绿峰面积 | 波长510—560 nm内原始光谱曲线所包围的面积 |
| SDr/SDb | 红边与蓝边面积的比值 | |
| SDr/SDy | 红边与黄边面积的比值 |
将上述筛选出的两类光谱指标分别与叶绿素含量进行线性回归分析, 得到一元线性回归方程。人工神经网络(ANN)模型[29]是基于生物学中神经网络的基本原理, 在理解和抽象了人脑结构和外界刺激响应机制后, 以网络拓扑知识为理论基础, 模拟人脑的神经系统对复杂信息的处理机制的一种数学模型。同样的, 将相关性较好的两类光谱指标分别代入输入层神经元, 将样本量随机分成50个建模训练集和41个模型验证集, ANN模型设置5个隐藏层节点, 最后输出SPAD含量。偏最小二乘回归(PLSR)模型[30]在建模过程中集中了主成分分析, 典型相关分析和线性回归分析方法的特点, 提供一种对多线性回归建模的方法, 特别当两组变量的个数很多、都存在多重相关性, 且观测数据的数量(样本量)又较少时, 其具有传统的经典回归分析等方法所没有的优点。利用原始反射率和筛选出的光谱特征参数作为建模参量, 将光谱样本随机分为训练样本集和验证样本集, 50个训练样本代入模型中得到估算模型, 41个验证样本做验证。
本文使用决定系数(R2)、均方根误差(RMSE)对估算模型进行评价, 且决定系数越大、均方根误差越小, 代表模型精度越高, 公式如下:
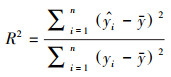
|
(2) |
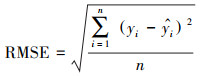
|
(3) |
式中: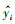
由图 1可知, 拔节期时不同处理间差异不显著。开花期时不同处理间叶片SPAD值无显著差异, 但此时臭氧胁迫处理下的叶片SPAD开始降低。随着生育期的推进, 在灌浆期时, 冬小麦叶片SPAD比前两个生育期低, 且不同处理间出现显著差异(P < 0.001)。从三个生育期的冬小麦叶片SPAD含量中可得出, 臭氧在拔节期对冬小麦叶片无影响, 在开花期开始出现影响, 在灌浆期影响最大。

|
| 图 1 不同生育期叶片叶绿素含量(SPAD)变化 Fig. 1 The average of SPAD value with and without ozone stress in three growth stages 不同小写字母表示处理间差异显著, P < 0.05 |
由图 2可知, 不同生育期的冬小麦叶片光谱反射率具有相似的光谱特征:在可见光波段(390—780 nm)存在明显的“绿峰”、“红谷”, 即在绿波段550 nm附近存在反射峰, 在红波段680 nm附近存在反射谷。在近红外波段(780—1000 nm), 叶片光谱反射率陡增, 增幅最大处为“红边”, 即一阶微分光谱的最大值。

|
| 图 2 不同生育期冬小麦叶片的平均高光谱响应特征 Fig. 2 Hyperspectral response of winter wheat leaves in different growth stages (包络线范围为正负一个标准差) (The envelope range is plus or minus one standard deviation) |
不同生育期, 臭氧胁迫对冬小麦叶片光谱特征的影响不同。拔节期时, 臭氧胁迫处理下的光谱曲线与自然对照处理下的光谱曲线无差异。开花期时, 臭氧对冬小麦叶片有轻微的影响, 主要体现在近红外波段内。该波段内臭氧胁迫处理下的叶片光谱反射率相比于自然对照处理下的叶片光谱反射率有升高的趋势, 此时臭氧对叶片光谱特征开始有影响。灌浆期时, 位于可见光波段内两种处理的叶片光谱反射率相较之前的生育期都有一定程度的升高。臭氧胁迫下的叶片光谱曲线在绿峰处升高, 且伴随有“红移”现象, 即绿峰中心位置向红光波段偏移。为了进一步验证臭氧在灌浆期对冬小麦光谱特征的影响, 对灌浆期的原始光谱反射率进行一阶微分处理(图 3)。相较于自然对照处理, 臭氧胁迫处理下叶片一阶微分光谱内700 nm波段附近的峰往左移动, 即红边位置向短波方向移动, 出现“蓝移”现象。蓝移、红移现象明显, 说明在灌浆期, 臭氧对叶片光谱特征有较大的影响。不同生育期冬小麦叶片的高光谱响应特征与上述不同生育期冬小麦叶片叶绿素含量变化规律一致, 说明臭氧胁迫下的冬小麦叶片光谱与叶绿素含量之间存在一定关系。

|
| 图 3 灌浆期两种处理下冬小麦叶片一阶微分光谱 Fig. 3 First-order differential spectra of winter wheat leaves under two ozone treatments at grain filling stage |
为构建臭氧胁迫下叶绿素含量与冬小麦叶片光谱的关系, 对所有候选的遥感植被指数以及光谱特征参数(表 1和表 2)与叶绿素含量进行相关性分析, 制作热力图矩阵(图 4), 矩阵左下部分为相关系数, 右上部分为热力图, 椭圆越扁相关性越好。将与叶绿素含量相关系数较高的8个遥感植被指数(|r|>0.9)和8个光谱特征参数(|r|≥0.85), 作为光谱指标, 用于后续构建模型, 包括:NDVI705、CI、CIredege、NDVI、mND705、SR705、DD、GM、AR、Rg、λg、Db、λr、SDb、SDg、SDr/SDb。

将上述相关性较好的光谱指标分别与SPAD值构建一元线性回归模型, 结果表明:遥感植被指数中NDVI705和mND705构建的一元线性回归模型最佳, R2达到0.852, RMSE为7.3 SPAD(表 3)。遥感植被指数构建的一元线性回归模型精度高、误差小, 较光谱特征参数而言更适合作为构建一元线性回归模型的参数。
| 光谱指标 Spectral index |
回归关系式 Regression |
叶绿素含量SPAD | ||
| r | R2 | 均方根RMSE | ||
| NDVI705 | y=98.245x+6.522 | 0.923** | 0.852 | 7.3 SPAD |
| mND705 | y=79.458x+6.383 | 0.923** | 0.852 | 7.3 SPAD |
| **表示通过0.001水平显著性检验, x为光谱特征参数, y为SPAD值, n=91 | ||||
将相关性较好的两类光谱指标分别与SPAD值构建人工神经网络叶绿素估算模型, 随机选取50个样本训练模型, 再用41个样本验证。结果表明:基于遥感植被指数的模型R2为0.86, RMSE为6.7 SPAD, 基于光谱特征参数的模型R2为0.89, RMSE为5.9 SPAD。人工神经网络模型的精度比一元线性回归模型的精度高, 均方根误差较低。以光谱特征参数建立人工神经网络模型精度更高, 误差更小(图 5)。

|
| 图 5 人工神经网络模型验证集拟合结果 Fig. 5 Fitting results of artificial neural network model validation set RMSE: 均方根误差 |
对原始光谱数据和光谱特征参数进行主成分分析。以原始数据为自变量时三个主成分能解释98%的因变量SPAD, 而以光谱特征参数为自变量时两个主成分就能解释98%的因变量。
从样本中随机抽取50份作为训练集, 剩下的41份作为验证集, 得出PLSR模型SPAD拟合结果(图 6)。结果表明:以原始数据为自变量的PLSR模型训练集的R2为0.87, RMSE为6.4 SPAD, 验证集的R2为0.78, RMSE为9.3 SPAD。以光谱特征参数为自变量进行PLSR建模, 训练集R2为0.97, RMSE为2.8 SPAD, 验证集R2为0.98, RMSE为2.3 SPAD。PLSR模型精度整体高于前两种模型, 且以光谱特征参数为自变量构建的PLSR模型精度最高, 误差最小。

|
| 图 6 偏最小二乘回归(PLSR)模型训练集和验证集叶绿素含量实测与预测的拟合关系 Fig. 6 Fitting of measured and estimated chlorophyll content in training set and validation set of PLSR model A、B基于原始光谱数据, C、D基于光谱特征参数 |
叶绿素能直接影响植物的光合速率, 反映植物的生理进程, 但臭氧胁迫会降低作物叶片的叶绿素含量。通过对臭氧胁迫下作物叶绿素含量的定量测定, 可得知作物受到臭氧伤害的程度。前期冬小麦在营养生长阶段, 由于此时的臭氧浓度较低, 积累臭氧量也较低, 臭氧对叶片的损害不大。随着生育期的推进, 在灌浆期时, 臭氧的累计浓度值已超过了植物的受害阈值, 因此导致叶片褪绿、褐斑等症状, 加速叶片衰老。已有报道:O3暴露下的细胞出现塌陷、液泡破裂、叶绿体结构受到破坏, 同时出现叶绿体体积和个数减少、叶绿体基质发生浓缩等现象, 严重影响叶绿素的正常合成[31], 导致臭氧胁迫处理下冬小麦叶片的叶绿素含量显著降低。Feng等[32—33]研究表明, 不同品种的小麦在灌浆期对臭氧浓度较为敏感, 其中叶绿素含量受到臭氧影响最为显著。Chi等[19]利用FACE平台研究也发现四个小麦品种在灌浆期受到臭氧胁迫最大。灌浆期的叶片叶绿素含量受到臭氧影响最大, 对冬小麦产量的影响也最大, 是今后臭氧胁迫监测研究需要重点关注的时间段。
3.2 臭氧胁迫对冬小麦叶片光谱特征的影响在可见光波段, 细胞内叶绿素、叶黄素、类胡萝卜素等色素的反射光谱均有响应, 主要的响应对象是叶绿素, 其主要吸收蓝光和红光为植物体内活动提供能量, 对绿光的吸收没有其他可见光吸收的多, 所以作物反射光谱曲线在蓝波段和红波段形成“波谷”, 在绿波段形成“波峰”[34]。袁小康等[35]研究表明, 当植物受到病虫害胁迫时, 绿色植物会出现“失绿”现象, 即叶片变黄, 绿峰升高且伴有红移趋势。本研究中, 拔节期和开花期叶片光谱在可见光波段无变化, 而灌浆期臭氧胁迫下的冬小麦叶片颜色变黄、叶绿素减少, 导致绿峰向红波段偏移、反射率增高, 产生“红移”现象。徐道青等[7]研究结果表明, 随着渍水胁迫程度的增加, 植物光谱中“绿峰”光谱反射率升高, 出现红移现象, 与本研究臭氧胁迫下小麦叶片的光谱响应现象一致。
在近红外波段, 作物叶片的反射光谱由叶片细胞的多层结构主导, 多层细胞间的多次反射对光谱的影响远大于色素的影响, 导致作物的反射光谱在近红外波段780—1000 nm之间陡增, 形成“红边”[34]。Gao等[31]研究表明, 随着O3胁迫时间的延长, 细胞会发生塌陷、细胞间距增大。本研究中, 开花期臭氧胁迫下冬小麦叶片光谱反射率在近红外波段升高, 在灌浆期这种变化却不明显。这可能是因为臭氧进入细胞内先伤害叶片内部细胞结构, 再破坏叶绿体。因此, 臭氧胁迫下开花期时的冬小麦叶片由于细胞结构的变化导致叶片光谱反射率在近红外波段升高, 而灌浆期影响更多的是叶绿素含量变化。Goswami等[14]通过对水稻进行氮、二氧化碳、温度升高等光谱研究, 认为反射光谱的红边位置是作物生长的胁迫检测指标。另有研究表明[34], 植物光谱近红外波段的红边位置朝着长波方向移动, 说明植物叶片叶绿素含量增加;朝着短波方向移动, 说明该植物因受到胁迫导致叶片在成熟期间叶绿素含量迅速减少。本研究中, 臭氧胁迫下灌浆期叶片叶绿素含量迅速降低, 导致叶片光谱在红边位置朝短波方向移动, 发生明显的“蓝移”现象, 但在拔节期和开花期却没有这种明显现象, 说明灌浆期小麦受到严重的臭氧影响。程小云等[15]研究结果也表明, 植物受到臭氧胁迫后, 作物光谱近红外波段的红边位置向短波移动。李庆禄[36]和张曼义[37]研究结果表明, 植物受到外界环境的胁迫, 红边位置会出现“蓝移”现象。此外, Pleijel等[38]在春小麦不同生育期进行相同O3剂量的处理试验, 发现在开花期至成熟期(生殖生长阶段)进行O3熏蒸引起的产量损失(-11%)远大于开花期前(营养生长阶段, -2%)。可见, 利用光谱技术观测不同时期臭氧胁迫冬小麦的研究结果与田间实验研究结论一致。
3.3 基于光谱的叶绿素含量估算模型本研究中, 将两类光谱指标代入不同建模方法进行比较, 找出适合臭氧胁迫下冬小麦叶片的叶绿素估算模型。通过变换不同的模型参数以及模型类型, 结果发现都有较好的估测精度, 且不同类型的光谱指标适合不同种模型。构建线性回归模型时, 遥感植被指数适合作为其建模参数。构建人工神经网络模型时, 光谱特征参数适合作为其建模参数。构建偏最小二乘回归模型时, 光谱特征参数适合作为其建模参数。其中以光谱特征参数为自变量的偏最小二乘法回归模型精度最高, 原因可能是其用少量的主成分估算因变量, 减少其他成分变量带来的误差, 且PLSR模型利用主成分分析提取了自变量最重要的信息, 在利用全部光谱信息的同时, 降低了数据的误差对模型的影响。李燕丽等[9]对渍水胁迫下小麦叶绿素含量进行估算研究, 发现使用偏最小二乘回归模型的精度最高, R2高达0.98。王伟东等[39]研究表明, 偏最小二乘回归模型能进一步提高模型的预测精度。虽然本研究的臭氧胁迫下冬小麦叶绿素估算模型取得较好的效果, 但由于实验样本量有限, 模型能否用于其他区域未知。此外, 由于光谱指标与胁迫没有一一对应的效果, 因此利用光谱监测臭氧胁迫时, 需要通过其他观测或经验知识排除其他胁迫的影响。总的来说, 利用光谱特征参数构建偏最小二乘回归模型估算臭氧胁迫下的叶绿素具有一定的可行性。
4 结论从臭氧胁迫下不同生育期冬小麦叶片光谱特征和叶绿素含量的变化规律中可以得出, 冬小麦从开花期开始受到臭氧影响, 在灌浆期明显增大。说明臭氧对冬小麦的损害是逐步累加的, 这对冬小麦臭氧胁迫监测时期的选择具有重要意义。筛选出与臭氧胁迫下叶片叶绿素含量相关性较好的光谱指标:NDVI705、CI、CIredege、NDVI、mND705、SR705、DD、GM、AR、Rg、λg、Db、λr、SDb、SDg、SDr/SDb, 有助于臭氧诱导冬小麦损伤的诊断和实时监测。利用光谱特征参数为自变量构建的偏最小二乘回归模型可为高臭氧浓度区域下冬小麦叶片受损的动态监测提供理论基础和技术方法。
| [1] |
冯兆忠, 李品, 袁相洋, 高峰, 姜立军, 代碌碌. 我国地表臭氧生态环境效应研究进展. 生态学报, 2018, 38(5): 1530-1541. |
| [2] |
Feng Z Z, Xu Y S, Kobayashi K, Dai L L, Zhang T Y, Agathokleous E, Calatayud V, Paoletti E, Mukherjee A, Agrawal M, Park R J, Oak Y J, Yue X. Ozone pollution threatens the production of major staple crops in East Asia. Nature Food, 2022, 3(1): 47-56. DOI:10.1038/s43016-021-00422-6 |
| [3] |
Kaliakatsou E, Bell J N B, Thirtle C, Rose D, Power S A. The impact of tropospheric ozone pollution on trial plot winter wheat yields in Great Britain-An econometric approach. Environmental Pollution, 2010, 158(5): 1948-1954. DOI:10.1016/j.envpol.2009.10.033 |
| [4] |
Ren X Y, Shang B, Feng Z Z, Calatayud V. Yield and economic losses of winter wheat and rice due to ozone in the Yangtze River Delta during 2014-2019. Science of the Total Environment, 2020, 745: 140847. DOI:10.1016/j.scitotenv.2020.140847 |
| [5] |
Mills G, Buse A, Gimeno B, Bermejo V, Holland M, Emberson L, Pleijel H. A synthesis of AOT40-based response functions and critical levels of ozone for agricultural and horticultural crops. Atmospheric Environment, 2007, 41(12): 2630-2643. DOI:10.1016/j.atmosenv.2006.11.016 |
| [6] |
车永飞, 肖策, 王正庆, 唐振平, 宋静, 李新春, 王升, 马艳. 基于遥感技术的植物非生物胁迫性研究进展. 南华大学学报: 自然科学版, 2020, 34(2): 34-41. |
| [7] |
徐道青, 刘小玲, 王维, 陈敏, 阚画春, 李常凤, 郑曙峰. 淹水胁迫下棉花叶片高光谱特征及叶绿素含量估算模型. 应用生态学报, 2017, 28(10): 3289-3296. |
| [8] |
夏天, 杨可明, 冯飞胜, 郭辉, 张超. 新型铜胁迫植被指数NCSVI探索铜污染下玉米叶片光谱敏感区间. 光谱学与光谱分析, 2021, 41(8): 2604-2610. |
| [9] |
李燕丽, 熊勤学, 卢碧林, 李继福, 李磊, 田广丽. 渍害胁迫下小麦SPAD的高光谱监测方法研究. 灌溉排水学报, 2020, 39(12): 41-48. |
| [10] |
谢凯, 蒋蘋, 罗亚辉. 稻瘟病胁迫下水稻叶片叶绿素含量与光谱特征参数的相关性研究. 中国农学通报, 2017, 33(17): 117-122. |
| [11] |
Wen W, Timmermans J, Chen Q, Van Bodegom P M. A review of remote sensing challenges for food security with respect to salinity and drought threats. Remote Sensing, 2021, 13(1): 6. |
| [12] |
Krishna G, Sahoo R N, Singh P, Bajpai V, Patra H, Kumar S, Dandapani R, Gupta V K, Viswanathan C, Ahmad T, Sahoo P M. Comparison of various modelling approaches for water deficit stress monitoring in rice crop through hyperspectral remote sensing. Agricultural Water Management, 2019, 213: 231-244. DOI:10.1016/j.agwat.2018.08.029 |
| [13] |
Ren Y, Huang W J, Ye H C, Zhou X F, Ma H Q, Dong Y Y, Shi Y, Geng Y, Huang Y R, Jiao Q J, Xie Q Y. Quantitative identification of yellow rust in winter wheat with a new spectral index: Development and validation using simulated and experimental data. International Journal of Applied Earth Observation and Geoinformation, 2021, 102: 102384. DOI:10.1016/j.jag.2021.102384 |
| [14] |
Goswami J, Das R, Sarma K K, Raju P L N. Red edge position (REP), an indicator for crop stress detection: implication on rice (Oryza sativa L). International Journal of Environment and Climate Change, 2021, 11(4): 88-96. |
| [15] |
程小云, 罗春旺, 刘琪璟, 周亚爽, 孟盛旺. O3胁迫对植物叶片光谱特征和叶绿素含量的影响. 环境科学与技术, 2017, 40(5): 1-8. |
| [16] |
Gab M, Hoffmann K, Lobe M, Metzger R, Van Ooyen S, Elbers G, Köllner B. NIR-spectroscopic investigation of foliage of ozone-stressed Fagus sylvatica trees. Journal of Forest Research, 2017, 11(2): 69-75. |
| [17] |
Marchica A, Loré S, Cotrozzi L, Lorenzini G, Nali C, Pellegrini E, Remorini D. Early detection of sage (Salvia officinalis L.) responses to ozone using reflectance spectroscopy. Plants, 2019, 8(9): 346. DOI:10.3390/plants8090346 |
| [18] |
Calzone A, Cotrozzi L, Remorini D, Lorenzini G, Nali C, Pellegrini E. Oxidative stress assessment by a spectroscopic approach in pomegranate plants under a gradient of ozone concentrations. Environmental and Experimental Botany, 2021, 182: 104309. DOI:10.1016/j.envexpbot.2020.104309 |
| [19] |
Chi G Y, Huang B, Shi Y, Chen X, Li Q, Zhu J G. Detecting ozone effects in four wheat cultivars using hyperspectral measurements under fully open-air field conditions. Remote Sensing of Environment, 2016, 184: 329-336. DOI:10.1016/j.rse.2016.07.020 |
| [20] |
付娆, 尚博, 张国友, 冯兆忠. 不同生育期臭氧熏蒸对水稻光合作用及生长的影响差异. 农业环境科学学报, 2021, 40(10): 2066-2075. |
| [21] |
冯兆忠, 徐彦森, 尚博. FACE实验技术和方法回顾及其在全球变化研究中的应用. 植物生态学报, 2020, 44(4): 340-349. |
| [22] |
Peñuelas J, Gamon J A, Fredeen A L, Merino J, Field C B. Reflectance indices associated with physiological changes in nitrogen-and water-limited sunflower leaves. Remote Sensing of Environment, 1994, 48(2): 135-146. DOI:10.1016/0034-4257(94)90136-8 |
| [23] |
Schlerf M, Atzberger C, Hill J. Remote sensing of forest biophysical variables using HyMap imaging spectrometer data. Remote Sensing of Environment, 2005, 95(2): 177-194. DOI:10.1016/j.rse.2004.12.016 |
| [24] |
Jordan C F. Derivation of leaf-area index from quality of light on the forest floor. Ecology, 1969, 50(4): 663-666. |
| [25] |
Gamon J A, Peñuelas J, Field C B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sensing of Environment, 1992, 41(1): 35-44. |
| [26] |
Daughtry C S T, Walthall C L, Kim M S, De Colstoun E B, McMurtrey Ⅲ J E. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sensing of Environment, 2000, 74(2): 229-239. |
| [27] |
Le Maire G, François C, Dufrêne E. Towards universal broad leaf chlorophyll indices using PROSPECT simulated database and hyperspectral reflectance measurements. Remote Sensing of Environment, 2004, 89(1): 1-28. |
| [28] |
伍南, 刘君昂, 闫瑞坤, 周国英. 利用光谱特征参数估算病害胁迫下杉木叶绿素含量. 植物保护, 2012, 38(4): 72-76, 88-88. |
| [29] |
朱大奇. 人工神经网络研究现状及其展望. 江南大学学报: 自然科学版, 2004, 3(1): 103-110. |
| [30] |
黄舜, 徐永海. 基于偏最小二乘回归的系统谐波阻抗与谐波发射水平的评估方法. 中国电机工程学报, 2007, 27(1): 93-97. |
| [31] |
Gao F, Calatayud V, García-Breijo F, Reig-Armiñana J, Feng Z Z. Effects of elevated ozone on physiological, anatomical and ultrastructural characteristics of four common urban tree species in China. Ecological Indicators, 2016, 67: 367-379. |
| [32] |
Feng Z Z, Pang J, Kobayashi K, Zhu J G, Ort D R. Differential responses in two varieties of winter wheat to elevated ozone concentration under fully open-air field conditions. Global Change Biology, 2011, 17(1): 580-591. |
| [33] |
Feng Z Z, Calatayud V, Zhu J G, Kobayashi K. Ozone exposure-and flux-based response relationships with photosynthesis of winter wheat under fully open air condition. Science of the Total Environment, 2018, 619-620: 1538-1544. |
| [34] |
Gates D M. Heat transfer in plants. Scientific American, 1965, 213(6): 76-84. |
| [35] |
袁小康, 周广胜, 王秋玲, 何奇瑾. 不同灌溉量夏玉米叶绿素含量的高光谱特征及其反演. 生态学报, 2021, 41(2): 543-552. |
| [36] |
李庆禄. 渍水胁迫下小麦高光谱特征分析与主要生理参数估测研究[D]. 扬州: 扬州大学, 2021.
|
| [37] |
张曼义. 干旱胁迫对黄瓜植株生长的影响机理及光谱估算模型的研究[D]. 南京: 南京信息工程大学, 2018.
|
| [38] |
Pleijel H, Danielsson H, Gelang J, Sild E, Selldén G. Growth stage dependence of the grain yield response to ozone in spring wheat (Triticum aestivum L.). Agriculture Ecosystems & Environment, 1998, 70(1): 61-68. |
| [39] |
王伟东, 常庆瑞, 王玉娜. 冬小麦叶片花青素相对含量高光谱监测. 麦类作物学报, 2020, 40(6): 754-761. |