 2023, Vol. 43
2023, Vol. 43文章信息
- 余海霞, 汤行昊, 刘南, 王浩, 张子雷, 邵长亮, 董刚, 曲鲁平
- YU Haixia, TANG Xinghao, LIU Nan, WANG Hao, ZHANG Zilei, SHAO Changliang, DONG Gang, QU Luping
- 控水与补水条件下连续热浪对闽楠光合特性和生长速率的影响
- Influences of multiple successive heat waves combined with water control and supplement on photosynthetic characteristics and growth rate of Phoebe bournei seedlings
- 生态学报. 2023, 43(8): 3224-3235
- Acta Ecologica Sinica. 2023, 43(8): 3224-3235
- http://dx.doi.org/10.5846/stxb202201190183
-
文章历史
- 收稿日期: 2022-01-19
- 网络出版日期: 2022-12-22





2. 福建省林业科学研究院, 福州 350002;
3. 中国农业科学院农业资源与农业区划研究所, 北京 100081;
4. 山西大学生命科学学院, 太原 030006
2. Fujian Academy of Forestry, Fuzhou 350002, China;
3. Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing 100081, China;
4. College of Life Science, Shanxi University, Taiyuan 030006, China
热浪通常是指持续一段时间高温的极端气候事件[1]。在全球气候变暖的背景下, 热浪的发生频率和强度明显增加, 危害程度也在增强[2]。研究表明, 高温胁迫并不是热浪影响植物生长发育的唯一因素, 热浪末期的水分胁迫也是影响植物生长发育的关键因素[3]。一方面, 热浪发生时的骤然高温比全球变暖的缓慢增温对生物的影响更大;另一方面, 这种持续的骤然高温过程通常伴随着水分胁迫的发生, 两者的交互作用会对人类和自然生态系统造成严重影响[4—6]。因此, 热浪对植物生长发育影响的研究并不应仅仅关注高温胁迫的影响, 更应偏重低水分环境下高温胁迫的影响, 即高温与干旱共同主导的复杂环境胁迫。目前, 国内外对植物抗逆生理学在热浪方面的研究多关注高温胁迫的影响, 这将导致热浪对植物生长发育影响机制理解的片面性。
光合作用是植物生长发育和产量形成的重要生理过程, 该过程对植物生长环境的变化十分敏感, 是研究植物抗逆性重要指标之一。如前所述, 热浪阶段除温度显著升高外, 还伴随着强光照和低降水等环境变化, 均会影响植物的气孔导度、蒸腾速率、光电子传输等相关的光合过程[7—9]。此外, 高温环境会显著增加植物的蒸发散和生态系统耗水量[10], 形成高水分亏缺(VPD)环境, 抑制植物的光合特性和生长速率, 导致水分利用效率(WUE)下降, 对植物造成短中长期影响[11]。因此, 研究高VPD环境下植物的光合特性对热浪的响应机制, 即高温干旱复合胁迫的影响, 将有助于植物抗逆生理研究在极端气候方向的发展。
闽楠(Phoebe bournei (Hemsl.) Yang)隶属于樟科(Lauraceae)楠属(PhoebeNees), 是我国东部亚热带常绿阔叶林优势树种之一, 属于国家二级珍稀渐危种, 具有极高的经济价值与生态意义。目前对闽楠的研究主要集中在种群分布[12]、光环境变化[13—14]、施肥[15—16]和CO2升高[17]等方面。但对于近年来亚热带地区热浪和季节性干旱频发的背景下, 热浪造成的复合胁迫对闽楠光合特性和生长速率的研究较少。此外, 亚热带地区近年来连续热浪时有发生, 即一年内可能存在多次热浪事件[18]。目前, 连续热浪对植物生长的影响尚无定论, 且闽楠光合特性和生长速率对连续热浪响应的研究未见报道。
基于此, 本研究通过人工模拟热浪的方法, 研究不同水分条件下连续热浪对闽楠幼苗光合特性和生长速率的影响, 探讨不同环境下热浪对闽楠光合特性和生长速率影响的差异, 评估和量化连续热浪对闽楠光合特性在短、中期的影响。由此本研究假设如下:(1)相较于补水组, 控水条件下热浪胁迫对闽楠的影响强度更高。(2)连续发生的热浪事件可能存在累加效应, 表现为经历过热浪的闽楠再次经历热浪时抗性下降。
1 材料和方法 1.1 试验地概况试验地位于福建省林业科学研究院苗圃, 地理坐标为26°9′2″N、东经119°17′2″E, 海拔153.9 m。试验地所在区域属典型的亚热带季风气候, 夏长冬短, 温暖湿润, 年平均降水量为1342 mm;最冷月1月平均为10.5 ℃, 最热月7月平均为28.2 ℃, 年平均气温为19.6 ℃, 年相对湿度约77%, 年平均日照数为1700—1980 h。地势平坦, 适宜开展各种苗木栽培试验。
1.2 试验设计 1.2.1 试验处理实验设计分为两个因素:人工模拟热浪和水分控制。热浪分为无热浪、单次热浪(持续5 d, H1)和两次热浪(每次持续5 d, 发生间隔时间30 d, H2)。水分组分为控水组(D)和补水组(W)。共6个处理, 分别是:无热浪控水组(D)、单次热浪控水组(DH1)、两次热浪控水组(DH2)、无热浪补水组(对照C)、单次热浪补水组(WH1)和两次热浪补水组(WH2)。每个处理即为一个开顶箱(OTC), OTC为长4.8 m、宽2.2 m、高2.2 m的矩形, 由28 mm的空心钢管组建, 每个OTC内包含6株闽楠苗木, 共36株。
1.2.2 试验材料研究选择长势均一、根系完整、无病害的两年生实生闽楠(Phoebe bournei (Hemsl.) Yang)苗木进行试验处理, 平均株高60.9 cm, 地径5.9 mm。试验前, 苗木种植于福建省林业科学院苗圃中, 在试验前半年将试验用苗木移植至花盆中。花盆为圆形, 尺寸为直径44 cm, 高度30 cm, 该尺寸确保花盆内部空间为苗木根系生长提供了充足的空间。土壤选择为收集于福州周边海拔500 m的红壤土, 总有机碳47.3 g/kg, 总氮3.2 g/kg, pH 4.9, 每盆干土重20 kg。同时将花盆水平埋在地面的土壤中, 避免花盆侧面空气升温对植物根系的影响。
1.2.3 人工热浪模拟方法及水分条件设置本研究通过OTC和电加热器共同作用的方式进行热浪模拟[19]。在热浪模拟阶段, OTC用高透光聚乙烯网布覆盖, 四周开小窗, 保证内外空气流通。在每个OTC内安装两个3.5 kW工业电加热器(BGE, 德国), 两台加热器放置在中线位置, 放置高度为2.0 m, 角度为20°, 以防止热风对幼苗直吹影响热浪模拟效果。提供四个轴向流动的风扇悬挂在加热器的两侧, 使OTC内空气流通。热控装置用于保证OTC内温度保持在30—40 ℃, 符合极端高温天气的标准[20]。保持日间增温幅度为5—10 ℃, 夜间为3—5 ℃, 实现了模拟热浪效果即昼夜不对称增温(白天增温幅度高于夜间, 日温差增大)(图 1)。为了避免OTC覆盖产生微气候变化影响研究结果, 无热浪处理组同样覆盖OTC, 但不放置加热装置。模拟热浪在9月进行, 避开自然热浪对热浪模拟试验的影响, 选择连续干燥的晴天进行热浪试验。除热浪阶段外, 拆除OTC, 安装遮光布以避免闽楠苗木受到自然热浪影响。其中, 热浪阶段隔绝自然降雨, 控水组(D)无每日水分补充, 补水组(W)每日补充3 L降水, 早晚通过喷灌装置浇灌。非热浪阶段, 控水组和补水组均按照苗圃日常管理, 不隔绝自然降雨, 且定时浇灌补水。

|
| 图 1 模拟热浪阶段微气候变化 Fig. 1 Microclimate variation during heat wave simulation 图中灰色区域为模拟热浪阶段 |
在热浪模拟试验期间及前后, 利用Li-6800便携式光合测定仪(Li-6800, LI-COR Bioscience, Lincoln, NE, 美国)在上午8点到10点测定闽楠幼苗的叶片净光合速率(Pn, mmol m-2 s-1), 气孔导度(Gs, mol m-2 s-1), 蒸腾速率(Tr, mmol m-2 s-1), 叶片饱和水汽压亏缺(Leafvpd, kPa)和实际光化学转化效率(ΦPSⅡ)等指标。测定时光强设定为1000 μmol·m-2s-1, CO2浓度设定为400 μmol/mol。叶片的水分利用效率(WUE, g/kg)的计算公式为:WUE= Pn/Tr。本研究通过计算热浪结束后不同处理条件下光合特性与对照组的差值比来观测闽楠在热浪后的恢复过程。
1.3.2 生态环境指标测定在试验地设置了微气象和水文观测系统, 监测5cm高度的空气温度、5 cm深度的土壤温度、地表温度和相对空气湿度。其中, 土壤温度的测量通过12个捷克(TOMST)进行持续测量, 每个OTC内放置2个;空气温湿度通过Lascar仪器的独立USB数据记录器记录, 每个OTC内放置1个。实验结果显示, 在热浪期间空气温度、土壤温度和地表温度分别平均增加了2.92 ℃, 2.22 ℃和2.72 ℃(图 1)。热浪处理下的温度显著高于无热浪处理组, 而无热浪处理条件下的相对空气湿度高于热浪处理组, 但在热浪期结束后, 热浪处理组与无热浪处理组不存在显著差异(图 1)。
1.3.3 苗高、地径测定将所有处理每株闽楠进行编号, 并于第一次热浪前和第二次热浪模拟后30 d对应编号进行苗高和地径等生长指标的测定, 然后通过相应公式计算生长率。
苗高相对增长率SHRGR(%)的计算公式为:
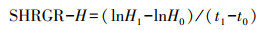
|
地径相对增长率GDRGR(%)计算公式:
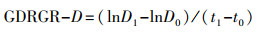
|
其中, H0和B0分别是热浪处理前闽楠苗高、地径, H1和B1分别是热浪模拟30 d后闽楠苗高、地径, t1-t0为时间间隔。
1.4 数据处理本研究通过双因素方差分析不同处理及其交互作用对热浪阶段光合特性(净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、水分利用效率(WUE)、叶片水汽压亏缺(Leafvpd)和实际光转化效率(ΦPSⅡ))的影响。通过单因素方法分析检验热浪阶段不同处理下光合特性和闽楠生长速率的差异(0.05水平, 邓肯t检验)。采用重复测量方差分析热浪后水分条件差异, 不同热浪频次和恢复时间交互作用下闽楠光合特性的恢复情况。采用Excel 2019软件进行数据整理, SPSS 26.0软件进行统计分析和检验, Origin 2018软件进行绘图。
2 结果与分析 2.1 热浪对闽楠苗木光合特性的影响试验阶段闽楠光合特性日动态变化表明热浪事件的发生会显著影响闽楠的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和水分利用效率(WUE)(图 2), 且该影响不会随着热浪的结束而马上消失, 存在明显的恢复过程。因此, 本研究中将热浪胁迫的影响分为热浪中和热浪后两个阶段, 对应评估闽楠对热浪的抵抗力和热浪后的恢复力。

|
| 图 2 不同处理下闽楠净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和水分利用效率(WUE)的日值变化 Fig. 2 Daily value variation of Phoebe bournei net photosynthesis rate (Pn), transpiration rate (Tr), stomatal conductance (Gs) and water use efficiency (WUE) under different treatments D: 无热浪控水组; DH1: 单次热浪控水组; DH2: 两次热浪控水组; C: 无热浪补水组; WH1: 单次热浪补水组; WH2: 两次热浪补水组; 图中灰色区域为两次热浪模拟阶段 |
本研究通过第二次热浪最后一日(10月19日)光合特性的变化评估闽楠对热浪胁迫的抵抗力。方差分析结果表明(表 1), 控水处理显著影响了闽楠Pn、Tr、Gs、Leafvpd和ФPSⅡ, 热浪显著影响了闽楠Tr、WUE和Leafvpd, 两种处理的交互作用显著影响了闽楠Leafvpd。
| 因素 Factors |
df | P | |||||
| 净光合速率 Pn/ (mmol m-2 s-1) |
蒸腾速率 Tr / (mmol m-2 s-1) |
气孔导度 Gs/ (mol m-2 s-1) |
水分利用效率 WUE/ (g/kg) |
叶片水汽压亏缺 Leafvpd/ kPa |
实际光转化效率 ΦPSⅡ |
||
| 控水Water control (a) | 1 | 0.004* | < 0.001* | 0.001* | 0.077 | < 0.001* | 0.002* |
| 热浪Heat wave (b) | 2 | 0.111 | < 0.001* | 0.432 | < 0.001* | < 0.001* | 0.467 |
| a × b | 2 | 0.356 | 0.207 | 0.801 | 0.504 | 0.001* | 0.312 |
| 统计分析基于第二次热浪处理最后一日(10/19)日值, *号表示显著性(P < 0.05) | |||||||
与对照组相比, 热浪的发生会显著升高闽楠Tr(P < 0.05), 且随着热浪发生次数增加。但干湿环境下升高幅度不同, 单次热浪补水组(WH1)和两次热浪补水组(WH2)比无热浪补水组(C)分别增加118%、207%, 单次热浪控水组(DH1)和两次热浪控水组(DH2)比无热浪控水组(D)分别增加91%、243%(图 3)。无论控水处理还是热浪均会显著降低闽楠的Pn, 其中补水热浪组WH1和WH2比C降低34%、25%, 控水热浪组DH1和DH2的比D降低6%、24%(图 3)。无论在控水或补水组, 热浪的发生均导致WUE的显著下降(P < 0.05), 其中DH1和DH2比D显著降低58%和83%, WH1和WH2比C显著降低75%和77%(图 3)。闽楠Gs和ФPSⅡ在热浪处理下升高, 但差异不显著, 但在控水组和补水组存在差异(图 3)。此外, 热浪的发生会显著增加闽楠Leafvpd(P < 0.05), 且该趋势在控水组和补水组一致(图 3)。

|
| 图 3 热浪阶段不同处理对闽楠蒸腾速率(Tr)、净光合速率(Pn)、水分利用效率(WUE)、气孔导度(Gs)、实际光化学效率(ФPSⅡ)和叶片水汽压亏缺(Leafvpd)的影响 Fig. 3 The effect of heat waves on Phoebe bournei transpiration rate (Tr), net photosynthesis rate (Pn), water use efficiency (WUE), stomatal conductance (Gs), PSⅡ operating efficiency in the light (ФPSⅡ) and leaf vapor pressure deficit (Leafvpd) during heat wave stage under different treatment 统计分析基于第二次热浪处理最后一日(10月19日)日值; 不同字母表示各处理之间存在显著差异(P < 0.05, 单因素方差分析, Duncan检验);图中数据为平均值±标准误差 |
热浪阶段Pn、Tr、Gs和WUE与Leafvpd线性拟合结果表明(图 4)。补水组闽楠Pn、Tr和WUE与Leafvpd显著相关(P < 0.05), 其中Pn和WUE与Leafvpd呈负相关, Tr与Leafvpd呈现正相关。控水组闽楠Leafvpd与Tr、Gs和WUE显著相关(P < 0.05), 其中WUE与Leafvpd呈负相关, Tr和Gs与Leafvpd呈现正相关。

|
| 图 4 热浪阶段净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和水分利用效率(WUE)与水汽压亏缺(Leafvpd)的线性关系图 Fig. 4 Linear relationships between net photosynthesis rate (Pn), transpiration rate (Tr), stomatal conductance (Gs) and water use efficiency (WUE) to leaf vapor pressure deficit (Leafvpd) during the heat wave period 统计分析基于第二次热浪处理最后一日(10月19日)日值, 仅保留显著关系(P < 0.05) |
本研究通过热浪后闽楠光合特性在30 d内的动态变化过程评估闽楠在经历不同频次热浪胁迫后的恢复能力。热浪发生后闽楠Pn、Tr、Gs和WUE均会随着恢复时间发生显著变化(表 2), 说明热浪事件后闽楠的光合特性存在恢复过程, 其中, 热浪对闽楠光合特性的影响强度会随着恢复时间延长而变化。该恢复过程可以通过热浪处理组闽楠光合特性指标与非热浪组的差值比表现(图 5)。其中, 热浪后结束第14 d(A14), 补水组WH1 Pn和Tr显著高于WH2、DH1和DH2(图 5)。补水组WH1 Gs显著高于DH1和DH2(P < 0.05), 而补水组WH2与WH1、DH1和DH2差异不明显(图 5)。WH1和WH2 WUE显著高于DH1和DH2。热浪结束第31 d(A31), 控水组DH1和DH2处理Pn、Tr和Gs显著高于补水组WH1和WH2(P < 0.05)(图 5)。DH2处理WUE显著高于DH1、WH1和WH2(P < 0.05), DH2处理WUE显著高于WH1和WH2(P < 0.05), 而WH1和WH2无显著差异(图 5)。
| 因素 Factors |
df | P | |||
| 净光合速率 Pn (mmol m-2 s-1) |
蒸腾速率 Tr (mmol m-2 s-1) |
气孔导度 Gs (mol m-2 s-1) |
水分利用效率 WUE (g/kg) |
||
| 控水water control (a) | 1 | < 0.001* | < 0.001* | 0.001* | 0.230 |
| 热浪Heat wave (b) | 2 | < 0.001* | 0.016* | 0.081 | 0.003* |
| a × b | 2 | 0.151 | 0.130 | 0.600 | 0.843 |
| 恢复时间Time (c) | 2 | < 0.001* | < 0.001* | < 0.001* | < 0.001* |
| a × c | 2 | 0.875 | < 0.001* | 0.028 | < 0.001* |
| b × c | 4 | < 0.001* | 0.001* | < 0.001* | 0.025* |
| a × b × c | 4 | 0.489 | 0.148 | 0.121 | 0.170 |
| 统计分析数据见图 5, *号表示显著性(P < 0.05) | |||||

|
| 图 5 热浪发生后闽楠净光合速率、蒸腾速率、气孔导度和水分利用效率差值比的变化 Fig. 5 Changes of Phoebe bournei net photosynthesis rate (ΔPn), transpiration rate (ΔTr), stomatal conductance (ΔGs) and water use efficiency (ΔWUE) variation since heat wave occurred H5:热浪的第5天(10月19日);A14:热浪后第14天(11月2日);A31:热浪后第31天(11月9日); 差值比计算公式为, 控水组:ΔPn (ΔTr/ΔGs/ΔWUE)=100%×(DH-D)/D; 补水组:ΔPn (ΔTr/ΔGs/ΔWUE)=100%×(WH-C)/C |
方差分析结果表明, 不同频次热浪处理显著影响闽楠幼苗的生长速率, 且该影响在不同水分环境下存在差异(表 3)。热浪结束后, WH2苗高增长率显著高于其他处理组, D显著高于DH1、DH2、WH1和, DH2和C显著高于DH1和WH1, 而DH2和C无显著差异, DH1和DH2差异不显著。
| 处理 Treatment |
苗高 H0/cm |
苗高 H1/cm |
苗高相对增长率 SHRGR/% |
地径 D0/mm |
地径 D1/mm |
地径相对增长率 GDRGR/% |
| D | 45.95±2.75 c | 63.60±3.32 bc | 0.21±0.04 ab | 4.77±0.32 a | 6.19±0.29 b | 0.17±0.05 |
| DH1 | 53.42±1.04 abc | 61.97±3.23 bc | 0.09±0.04 c | 3.94±0.20 b | 5.77±0.13 b | 0.25±0.04 |
| DH2 | 54.33±2.34 ab | 69.38±1.51 ab | 0.16±0.03 bc | 4.90±0.22 a | 6.65±0.37 ab | 0.20±0.04 |
| C | 49.65±2.96 bc | 58.37±1.28 c | 0.11±0.03 bc | 5.14±0.27 a | 7.25±0.42 a | 0.23±0.03 |
| WH1 | 59.47±2.31 a | 68.35±4.36 ab | 0.09±0.04 c | 3.85±0.28 b | 5.92±0.29 b | 0.29±0.03 |
| WH2 | 48.85±2.89 bc | 74.32±2.24 a | 0.28±0.04 a | 5.01±0.16 a | 6.42±0.27 ab | 0.16±0.04 |
| 表中数据为平均值±标准差, 同列数据后不同字母表示各处理之间有显著性差异(P < 0.05, 邓肯检验); 苗高相对增长率(Seeding height relative growth rate)=(lnH1-lnH0)/(t1-t0), 地径相对增长率(Ground diameter relative growth rate)=(lnD1-lnD0)/(t1-t0), t0为6月30日测量, t1为11月30日测量; D: 无热浪控水组No heat wave without water; DH1: 单次热浪控水组One heat wave without water; DH2: 两次热浪控水组Two heat wave without water; C: 无热浪补水组No heat wave with water; WH1: 单次热浪补水组One heat wave with water; WH2: 两次热浪补水组Two heat wave with water | ||||||
植物的光合特性随环境变化是植物在进化过程中逐渐形成的重要生存策略, 植物可以通过调节光合特性应对气候变化的影响[21]。光合反应对热浪伴随的骤然增温的反应十分敏感, 这是因为当极端高温超出植物的耐受阈值时植物Rubisco活化酶(1.5-二磷酸核酮糖羧化酶)活性降低和光系统PSⅡ受损, 表现为光合效率的降低[22]。本研究中, 热浪胁迫显著抑制闽楠的光合作用, 与对照组相比, 单次热浪导致闽楠净光合速率(Pn)下降了34%(补水组)和39%(控水组)。这与亚热带地区阔叶树种木荷(Schima superba)和针叶树种杉木(Cunninghamia lanceolata)净光合速率对热浪的响应表现为下降55%和51%研究结果相一致[19]。因此, 由于光合系统的高温耐受性差异, 不同树种对于热浪的抗性不同, 净光合速率受到抑制的幅度也存在差异[9, 23]。
本研究发现控水和补水处理下, 热浪对闽楠净光合特性的影响不同, 特别是对蒸腾速率和气孔导度的影响存在显著差异(图 3)。已有研究表明植物对于热浪胁迫的应激反应多表现为增大其气孔导度, 加速蒸腾作用通过叶片水分的蒸发来降低叶片的温度[7, 24], 这种现象对于经常面对高温胁迫且区域水量丰沛地区的植物更加明显[25]。本研究中热浪阶段闽楠的蒸腾速率和气孔导度显著增加, 并且差异程度随着热浪发生次数的增加而升高, 但补水组的蒸腾速率和气孔导度高于控水组(图 3), 这是因为同一植物面对不同的环境条件表现出不同的水分调节行为[26]。高温条件下植物的高蒸发和强耗水与低降水极易造成生态系统的水分亏缺, 植物应对水分胁迫的敏感性与调节方式存在差异, 表现为等水和非等水两种调节方式:等水行为的植物在面临水分亏缺时通过降低气孔大小来减少水分的散失, 较小的气孔对干旱的反应迅速, 可以使植物避免木质部栓塞[27], 但较小的气孔导度也能降低植物的蒸腾作用从而减少水分的散失, 其光合速率也随之降低[28];非等水行为的植物将气孔维持在较高的开放水平, 其中茎的水势随水分胁迫的增强而减小同时保持较高的碳同化能力[29]。因此, 热浪胁迫下闽楠的水分调节方式趋近于非等水行为, 且水分供给有利于缓解热浪胁迫下的高水分亏缺。
不同控水环境下, 闽楠的蒸腾速率变化程度不一致, 但水分利用效率均显著下降, 且不存在显著差异(图 3)。该结论与前期热浪对植物影响的研究一致, 植物遭受热浪胁迫时表现为水分利用效率的显著下降, 从而保证植物生长与水分消耗的平衡[11, 19]。补水条件下热浪阶段蒸腾速率的增加是水分效率降低的主要原因, 闽楠可能通过加速蒸腾维持叶片温度, 保证光合速率的稳定。研究发现高温条件下, 植物可以通过高蒸腾降低热胁迫的强度[7], 但是植物水分的散失不受控制, 叶片水分甚至可能通过叶片角质层丧失, 与气孔导度控制无关, 使水分的消耗量与光合强度关系强度降低[30]。而本研究中补水组闽楠热浪阶段气孔导度与水分胁迫不存在显著线性关系, 控水条件下闽楠在热浪阶段蒸腾强度和光合强度降低幅度的不一致性可能是导致其水分利用效率下降的主要原因。因此, 不同水分环境条件下主导热浪阶段闽楠水分利用效率下降的原因是不同的, 且热浪期间的水分供给缓解了光合和蒸腾速率的降低。
高温干旱复合胁迫下的植物通常面临两难选择:如果气孔调节保证水分供给, 降温调节功能必然降低, 可能遭受热损伤;如果加速耗水增加降温调节功能效率, 将加剧水分胁迫, 引发水力障碍, 导致植物干旱致死[19, 31]。本研究中, 控水组闽楠在热浪处理下净光合速率与水分胁迫无显著线性关系(图 4), 可能是闽楠在不同复合胁迫下选择不同所造成的。因此, 闽楠光合特性在热浪的复合胁迫下的变化与单一胁迫下的响应不同, 其最终呈现的响应方式应该是两难选择下平衡的结果, 且可能随着热浪持续时间变长下两种胁迫方式的强度变化而出现响应方式的变化。
3.2 闽楠光合特性在热浪后的恢复及连续热浪的累加效应热浪事件的持续时间虽短(一般定义为持续5 d以上), 但可能对植物造成深远的影响, 且热浪后植物的光合特性存在明显的恢复过程[32]。热浪对植物/生态系统的影响分为亚致死影响、致死影响、间接影响和复合影响[33]。其中, 复合影响指的是热浪高温事件与常伴随发生干旱或火灾等事件的共同影响, 该复合影响可能增强热浪的亚致死影响或致死影响, 其本质就是影响植物的恢复过程[34]。本研究中, 热浪结束14 d后, 多处理组闽楠净光合速率、蒸腾速率恢复至未干扰水平(图 5), 这说明闽楠光合特性在热浪事件后也存在着恢复过程。植物对极端气候的耐受性可以分为极端气候事件发生阶段植物的抗性, 即植物抵御环境干扰的能力;以及植物在极端气候影响后的恢复力, 即植物在热浪干扰后恢复至未干扰状态的能力[35]。如果低抵抗性的植物具备高恢复力, 可以在热浪后快速恢复抵消影响, 反之亦然。本研究中闽楠在不同控水条件下光合特性的恢复过程也存在显著差异, 特别在31 d后表现出的补偿性的增长(图 5)。因此, 闽楠在热浪后具有较强的恢复能力, 特别是在复合胁迫的强干扰后, 可以通过光合特性的补偿性增长得加速恢复, 甚至可能超过没有受到干扰的状态, 但该补偿性的高速增长能持续多长时间仍未所知。
热浪发生频率快于植物恢复所需时间, 连续热浪可能对植物造成负面的影响, 有研究表明多年连续的热浪对草地生态系统的影响存在累加效应。该累加效应可能表现为负面效果的累加, 例如, 连续热浪影响下草地生态系统净初级生产力降低幅度逐年增加[11];也可能表现为正面的效果, 例如, 连续热浪影响下桉树(Corymbia calophylla)叶片存在生理可塑性, 连续热浪表现为增强了桉树叶片的高温耐受性及更广的温度安全范围[35]。本研究中连续发生的热浪对闽楠蒸腾速率, 气孔导度、水分利用效率的影响强度都与单次热浪的影响存在显著差异(图 3), 说明闽楠对热浪事件存在记忆。同样在热浪后14 d时, 连续热浪下的闽楠光合特性与单次热浪影响下存在显著差异, 表现为将光合速率、蒸腾速率和气孔导度的变化幅度显著下降和水分利用效率变化幅度的显著升高, 但这种差异在热浪31 d后消失(图 5)。如此, 连续热浪的对闽楠的累加效应主要集中在热浪阶段及热浪后的一段时间内, 但随着闽楠的恢复过程而逐渐消失。
热浪对闽楠光合特性的影响将最终表现为对闽楠生长速率的影响。在本研究中, 热浪胁迫后补水组闽楠苗高增长率显著高于控水组(表 3)。说明闽楠在高温胁迫下, 如果保证充足的水分供给, 在热浪后将获得补偿性生长, 促进苗高增长;若水分胁迫发生, 闽楠光合能力和水分利用效率将下降, 进而抑制植物的生长[36]。水分亏缺的条件下, 水势降低导致细胞扩张和细胞分裂功能减弱, 以及光合作用的降低, 最终导致碳同化减少, 抑制植物生长[37—38]。这与本文研究闽楠在控水条件下, DH1和DH2的苗高增长率显著低于D结果相一致。实际上, 有研究表明, 土壤水分充沛条件下的热浪可能会通过改变植物形态, 例如, 增加根冠比而促进植物后期的生长[19]。本研究中闽楠在补水处理组WH2的苗高增长率显著高于WH1和C, 同样表明热浪胁迫反而促进了闽楠的苗高生长。然而, 本研究中各处理组间地径增长率无显著差异, 该结论可能是由于热浪胁迫下闽楠地上植物形态趋向于向高处生长。另一方面, 可能是由于本研究中对闽楠生长指标观测的时间跨度不够长, 造成闽楠的地径增长率对热浪的响应无显著差异, 可能仍需更长期观测试验数据的证明。但本研究仅关注了热浪对闽楠地上部分生长的影响, 缺乏对植物地下生长和营养物质分配的研究, 无法准确评估热浪对植物生长发育的影响。
4 结论本研究结果表明, 热浪阶段闽楠净光合速率和蒸腾速率均显著下降, 但不同控水条件下蒸腾速率, 水分利用效率的变化存在差异。其中, 补水条件下光合速率的变化是闽楠生理上主动调控的结果, 表现为增加耗水散热减少光合系统损伤, 缓解了热浪胁迫对植物光合过程的影响;且在热浪事件后, 闽楠的净光合速率在较短时间内(15 d)即恢复至未受干扰水平。表明若闽楠在热浪阶段能获得水分补给, 可以增强植物在热浪阶段的抗性及加速热浪后植物的恢复速度。反之, 控水条件下高水分亏缺环境可能加剧热浪对闽楠光合特性的影响, 降低闽楠对热浪的抗性和恢复力。此外, 连续发生的热浪存在累加效应, 主要表现为降低闽楠对热浪的抗性, 但对恢复力影响不大。综上, 亚热带阔叶树种闽楠的热浪耐受性与热浪阶段生态系统的水分环境存在关联, 能否保证水分供给是闽楠能否表现出优良热浪耐受性的关键。由此可见, 植物在单逆境环境下和复合逆境环境下的生理表现存在差异, 因此未来对于亚热带地区复杂气候变化环境下植物抗逆生理的研究更应关注高温干旱等多胁迫因子复合作用对植物生长的影响。
| [1] |
IPCC. Climate Change 2021:The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the IPCC. Cambridge: Cambridge University Press, 2021.
|
| [2] |
Easterling D R, Meehl G A, Parmesan C, Changnon S A, Karl T R, Mearns L O. Climate extremes: observations, modeling, and impacts. Science, 2000, 289(5487): 2068-2074. DOI:10.1126/science.289.5487.2068 |
| [3] |
De Boeck H J, Bassin S, Verlinden M, Zeiter M, Hiltbrunner E. Simulated heat waves affected alpine grassland only in combination with drought. New Phytologist, 2016, 209(2): 531-541. DOI:10.1111/nph.13601 |
| [4] |
Piao S L, Zhang X P, Chen A P, Liu Q, Lian X, Wang X H, Peng S S, Wu X C. The impacts of climate extremes on the terrestrial carbon cycle: a review. Science China Earth Sciences, 2019, 62(10): 1551-1563. DOI:10.1007/s11430-018-9363-5 |
| [5] |
De Boeck H J, Dreesen F E, Janssens I A, Nijs I. Whole-system responses of experimental plant communities to climate extremes imposed in different seasons. New Phytologist, 2011, 189(3): 806-817. DOI:10.1111/j.1469-8137.2010.03515.x |
| [6] |
Xu H, Xiao J F, Zhang Z Q. Heatwave effects on gross primary production of northern mid-latitude ecosystems. Environmental Research Letters, 2020, 15(7): 074027. DOI:10.1088/1748-9326/ab8760 |
| [7] |
Drake J E, Tjoelker M G, Vårhammar A, Medlyn B E, Reich P B, Leigh A, Pfautsch S, Blackman C J, López R, Aspinwall M J, Crous K Y, Duursma R A, Kumarathunge D, De Kauwe M G, Jiang M K, Nicotra A B, Tissue D T, Choat B, Atkin O K, Barton C V M. Trees tolerate an extreme heatwave via sustained transpirational cooling and increased leaf thermal tolerance. Global Change Biology, 2018, 24(6): 2390-2402. DOI:10.1111/gcb.14037 |
| [8] |
De Boeck H J, Van De Velde H, De Groote T, Nijs I. Ideas and perspectives: heat stress: more than hot air. Biogeosciences, 2016, 13(20): 5821-5825. DOI:10.5194/bg-13-5821-2016 |
| [9] |
Guha A, Han J M, Cummings C, McLennan D A, Warren J M. Differential ecophysiological responses and resilience to heat wave events in four co-occurring temperate tree species. Environmental Research Letters, 2018, 13(6): 065008. DOI:10.1088/1748-9326/aabcd8 |
| [10] |
王国华, 赵文智, 刘冰, 常学向, 张智慧. 河西走廊荒漠-绿洲蒸散对夏季高温天气响应的初步研究. 干旱区研究, 2013, 30(1): 173-181. |
| [11] |
Qu L P, Chen J Q, Dong G, Shao C L. Heavy mowing enhances the effects of heat waves on grassland carbon and water fluxes. Science of the Total Environment, 2018, 627: 561-570. DOI:10.1016/j.scitotenv.2018.01.287 |
| [12] |
吴大荣, 王伯荪. 濒危树种闽楠种子和幼苗生态学研究(英文). 生态学报, 2001, 21(11): 1751-1760. |
| [13] |
唐星林, 刘光正, 姜姜, 刘斌, 张运兴, 狄岚. 遮阴对闽楠一年生和三年生幼树叶绿素荧光特性及能量分配的影响. 生态学杂志, 2020, 39(10): 3247-3254. |
| [14] |
王振兴, 朱锦懋, 王健, 汪滢, 卢钰茜, 郑群瑞. 闽楠幼树光合特性及生物量分配对光环境的响应. 生态学报, 2012, 32(12): 3841-3848. |
| [15] |
杨志坚, 冯金玲, 吴小慧, 陈来贺, 余新晓, 陈丽华. 氮磷钾施肥对闽楠幼苗营养元素吸收与利用的影响. 生态学杂志, 2021, 40(4): 998-1011. |
| [16] |
许文斌, 余坦蔚, 洪小敏, 李梦娇, 胡亚林. 亚热带4种典型人工林幼树光合特征和生物量对土壤水肥因子的响应. 福建农林大学学报: 自然科学版, 2021, 50(1): 61-68. |
| [17] |
王晓, 韦小丽, 吴高殷, 陈胜群. CO2浓度升高条件下不同氮素供应对闽楠幼苗光合特性及生长的影响. 林业科学, 2021, 57(4): 173-181. |
| [18] |
安宁, 左志燕. 1961-2017年中国地区热浪的结构变化. 中国科学: 地球科学, 2021, 51(8): 1214-1226. |
| [19] |
Qu L P, De Boeck H J, Fan H H, Dong G, Chen J Q, Xu W B, Ge Z Q, Huang Z J, Shao C L, Hu Y L. Diverging responses of two subtropical tree species (Schima superba and Cunninghamia lanceolata) to heat waves. Forests, 2020, 11(5): 513. DOI:10.3390/f11050513 |
| [20] |
谭红建, 蔡榕硕. 2000年以来福州地区夏季极端高温的新特征及成因探讨. 大气科学, 2015, 39(6): 1179-1190. |
| [21] |
Teskey R, Wertin T, Bauweraerts I, Ameye M, McGuire M A, Steppe K. Responses of tree species to heat waves and extreme heat events. Plant, Cell & Environment, 2015, 38(9): 1699-1712. |
| [22] |
Dusenge M E, Wittemann M, Mujawamariya M, Ntawuhiganayo E B, Zibera E, Ntirugulirwa B, Way D A, Nsabimana D, Uddling J, Wallin G. Limited thermal acclimation of photosynthesis in tropical montane tree species. Global Change Biology, 2021, 27(19): 4860-4878. DOI:10.1111/gcb.15790 |
| [23] |
Duarte A G, Katata G, Hoshika Y, Hossain M, Kreuzwieser J, Arneth A, Ruehr N K. Immediate and potential long-term effects of consecutive heat waves on the photosynthetic performance and water balance in Douglas-fir. Journal of Plant Physiology, 2016, 205: 57-66. |
| [24] |
Wang D, Heckathorn S A, Mainali K, Tripathee R. Timing effects of heat-stress on plant ecophysiological characteristics and growth. Frontiers in Plant Science, 2016, 7: 1629. |
| [25] |
Wu G L, Liu H, Lei H, Qi L, Lin Y X, He P C, Feng S W, Liu J X, Ye Q. Differential responses of stomata and photosynthesis to elevated temperature in two Co-occurring subtropical forest tree species. Frontiers in Plant Science, 2018, 9: 467. |
| [26] |
Conesa M R, De La Rosa J M, Domingo R, Bañon S, Pérez-Pastor A. Changes induced by water stress on water relations, stomatal behaviour and morphology of table grapes (cv. Crimson Seedless) grown in pots. Scientia Horticulturae, 2016, 202: 9-16. |
| [27] |
Mitchell P J, O'grady A P, Tissue D T, White D A, Ottenschlaeger M L, Pinkard E A. Drought response strategies define the relative contributions of hydraulic dysfunction and carbohydrate depletion during tree mortality. New Phytologist, 2013, 197(3): 862-872. |
| [28] |
Upadhyay R K, Soni D K, Singh R, Dwivedi U N, Pathre U V, Nath P, Sane A P. SlERF36, an EAR-motif-containing ERF gene from tomato, alters stomatal density and modulates photosynthesis and growth. Journal of Experimental Botany, 2013, 64(11): 3237-3247. |
| [29] |
McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Wliiams D G, Yepez E A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. New Phytologist, 2008, 178(4): 719-739. |
| [30] |
Duursma R A, Blackman C J, Lopéz R, Martin-StPaul N K, Cochard H, Medlyn B E. On the minimum leaf conductance: its role in models of plant water use, and ecological and environmental controls. New Phytologist, 2019, 221(2): 693-705. |
| [31] |
McDowell N G, Grossiord C, Adams H D, Pinzón-Navarro S, Mackay D S, Breshears D D, Allen C D, Borrego I, Dickman L T, Collins A, Gaylord M, McBranch N, Pockman W T, Vilagrosa A, Aukema B, Goodsman D, Xu C G. Mechanisms of a coniferous woodland persistence under drought and heat. Environmental Research Letters, 2019, 14(4): 045014. |
| [32] |
Ruehr N K, Grote R, Mayr S, Arneth A. Beyond the extreme: recovery of carbon and water relations in woody plants following heat and drought stress. Tree Physiology, 2019, 39(8): 1285-1299. |
| [33] |
Breshears D D, Fontaine J B, Ruthrof K X, Field J P, Feng X, Burger J R, Law D J, Kala J, Hardy G E S J. Underappreciated plant vulnerabilities to heat waves. New Phytologist, 2021, 231(1): 32-39. |
| [34] |
Ahrens C W, Challis A, Byrne M, Leigh A, Nicotra A B, Tissue D, Rymer P. Repeated extreme heatwaves result in higher leaf thermal tolerances and greater safety margins. New Phytologist, 2021, 232(3): 1212-1225. |
| [35] |
Ingrisch J, Bahn M. Towards a comparable quantification of resilience. Trends in Ecology & Evolution, 2018, 33(4): 251-259. |
| [36] |
Muller B, Pantin F, Génard M, Turc O, Freixes S, Piques M, Gibon Y. Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs. Journal of Experimental Botany, 2011, 62(6): 1715-1729. |
| [37] |
Ameye M, Wertin T M, Bauweraerts I, McGuire M A, Teskey R O, Steppe K. The effect of induced heat waves on Pinus taeda and Quercus rubra seedlings in ambient and elevated CO2 atmospheres. New Phytologist, 2012, 196(2): 448-461. |
| [38] |
Smith A M, Stitt M. Coordination of carbon supply and plant growth. Plant, Cell & Environment, 2007, 30(9): 1126-1149. |