 2023, Vol. 43
2023, Vol. 43文章信息
- 许文文, 赵燕翘, 王楠, 赵洋
- XU Wenwen, ZHAO Yanqiao, WANG Nan, ZHAO Yang
- 人工蓝藻结皮对沙区表层土壤酶活性及其恢复速率的影响
- Effects of artificial cyanobacterial crusts on enzyme activities and recovery rate of surface soil in sandy areas
- 生态学报. 2023, 43(7): 2856-2864
- Acta Ecologica Sinica. 2023, 43(7): 2856-2864
- http://dx.doi.org/10.5846/stxb202204201075
-
文章历史
- 收稿日期: 2022-04-20
- 网络出版日期: 2022-11-19






2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
土壤酶是指由土壤微生物、植物根系、土壤动物及动植物残体等分泌的生物催化剂[1]。土壤酶参与土壤中的各种化学反应, 其活性不但可以判断出该地区微生物的活性大小, 也是反映土壤功能的重要指标之一, 通过对干旱区土壤酶活性的研究有助于了解该地土壤肥力以及土壤功能的恢复情况[2]。
生物土壤结皮(Biological Soil Crusts, BSCs)是由藻类、地衣、藓类等隐花植物和细菌、真菌等土壤中的微生物通过菌丝体、假根、分泌物等与土壤表层颗粒胶结形成的地表稳定复合体, 是荒漠生态系统中一种常见的群落结构, 具有防风固沙、水文调节、增加土壤养分等多种重要生态功能[3—4]。目前, 国内外在干旱区关于BSCs对土壤酶活性的影响已开展了一些工作, 樊瑾等[5]在毛乌素沙地发现BSCs可以显著提高土壤酶活性, 且演替后期的藓类结皮土壤酶活性显著高于演替初期藻结皮的土壤酶活性, 同时随着土层加深, 土壤酶活性逐渐降低, 这与Liu等[6]在腾格里沙漠东南缘人工植被区和Zhang[7]等在古尔班通古特沙漠所得结果一致, 在国外的荒漠地区也有相同的结论[8]。同时这些研究结果还表明土壤酶的活性不但随季节而变化, 并且与土壤中的理化性质高度相关, 而BSCs的存在会对土壤水分以及土壤养分等产生影响, 不仅如此, BSCs的基本特征也会影响土壤酶活性[9], 因此, BSCs可以通过多方面作用对土壤酶活性产生影响。
人工BSCs指将BSCs中的主要生物体(藻类、地衣和藓类)进行人工培育并接种到沙地表面, 通过养护成活, 加快地表BSCs的形成, 是一种新兴的绿色环保型固沙方法。人工BSCs技术能够缩短BSCs的形成时间, 使沙面趋于稳定, 增强其抗风蚀能力, 并且在BSCs形成后可以改善土壤肥力, 促进荒漠植被的生长, 是一种有效治理沙化土地的新技术[10—11]。近年来, 一些学者相继开展了关于人工BSCs技术方面的研究, 证实了其固沙的可行性[12—13], 但涉及其对土壤酶活性影响的研究却相当薄弱[14—15], 如发育初期的人工BSCs对土壤酶有何影响?与自然发育的BSCs相比恢复速率如何?因此, 开展人工BSCs对沙区土壤酶活性及恢复速率的影响等方面的研究, 可以直接或间接地反映人工BSCs技术对土壤质量恢复的作用, 对深入评估人工BSCs在荒漠生态系统恢复中的作用和机理具有重要的理论支撑。
基于此, 本研究以腾格里沙漠东南缘的人工蓝藻结皮、自然发育的蓝藻结皮和流沙为研究对象, 分析了人工蓝藻结皮的形成与发育是否改变了表层(0—2 cm)土壤蔗糖酶、纤维素酶、淀粉酶、过氧化氢酶的活性, 探讨了人工蓝藻结皮的基本特征对土壤酶的影响, 并计算其与人工植被区自然发育的蓝藻结皮表层土壤酶活性相比的恢复速率, 这些研究补充了目前关于人工蓝藻结皮对荒漠生态系统土壤功能恢复方面研究的不足, 并且有助于深入了解人工蓝藻结皮对土壤环境恢复的影响, 对人工蓝藻结皮治理荒漠化具有重要理论意义, 也将为荒漠生态系统修复提供重要实践指导。
1 研究区概况研究区位于腾格里沙漠东南缘沙坡头沙漠试验站(37°27′35″N, 105°00′37″E), 属于草原化荒漠地带, 温带大陆性干旱气候显著, 该区海拔1250 m, 年降雨量186 mm, 主要集中在6—8月份, 年蒸发3000 mm, 年平均温度9.6 ℃, 年均风速2.8 m/s, 气候干燥、高温、多风。土壤基质为松散贫瘠的流沙, 主要土壤类型为灰钙土和风沙土。该区人工植被主要物种有花棒(Corethrodendron scoparium)、柠条(Caragana korshinskii)、油蒿(Artemisia ordosica)、砂蓝刺头(Echinops gmelinii)、小画眉草(Eragrostis poaeoides)等, 天然植被区中常见草本植物有沙米(Agriophyllum squarrosum)、虫实(Artemisia desertorum)、刺蓬(Salsola tragus)、雾冰藜(Bassia dasyphylla)、小画眉草等[16]。
2 研究方法 2.1 试验设计荒漠蓝藻的大规模培养。人工蓝藻结皮所用蓝藻藻种为为鱼腥藻(Anabaena sp.)、念珠藻(Nostoc sp.)、席藻(Phormidium sp.)、伪枝藻(Scytonema sp.)和单歧藻(Tolypothrix sp.), 培养基为BG·11。5种混合蓝藻先用三角瓶在无菌条件下置于摇床上进行一级培养, 然后将培养10—15 d的5种混合蓝藻转入室内白色塑料箱(0.5 m×0.8 m×0.6 m)中进行二级扩繁, 培养条件为温度(25±2)℃、光照强度4000 lux, 光暗比12 h: 12 h, 最后将白色塑料箱中培养10—15 d的荒漠蓝藻转入蓝藻培养池内(1.0 m×1.0 m×15.0 m)进行规模化培养, 在指数阶段收集蓝藻后备用。
野外接种。2018年8月初, 在野外建立面积为3.0 m2(1.5 m×2.0 m)的试验样地(流动沙丘推平后建立的试验样地), 将收集的新鲜蓝藻均匀的喷洒在试验区地表, 用量200 g/m2(约10 g干重, 0.2 mm厚度), 然后用无纺布、防尘网和遮阳网进行覆盖, 共设置5个处理, 每个处理4个重复。具体处理为:1)新鲜蓝藻覆盖两层无纺布(NWF);2)新鲜蓝藻覆盖防尘网(DPN);3)新鲜蓝藻覆盖遮阳网(SSN);4)新鲜蓝藻先覆盖两层无纺布, 再覆盖防尘网(NWF+DPN);5)新鲜蓝藻先覆盖两层无纺布, 再覆盖遮阳网(NWF+SSN)。实验布设方法详见Zhao等[17]。
人工植被区为宁夏中卫中国科学院沙坡头沙漠试验站2010年建植, 配置类型为柠条和油蒿混合种植, 株行距分别为1.9 m×1.5 m, 0.9 m×1.5 m。经过多年固沙实践证明该区最适宜的固沙物种包括柠条与油蒿, 而这两种植物搭配种植不仅在地上部分可以有效控制风蚀, 而且地下部分由于根系长度的不同能够有效利用土壤水分与养分等, 因此通过柠条与油蒿搭配种植后可以有效的控制沙丘的移动, 并经过多年演替后, 形成了一个复杂的人工—天然的荒漠生态系统, 因此本研究选择柠条和油蒿混合种植作为对照组[16]。
2.2 样品采集大量研究表明, BSCs土壤酶活性夏季高于其它季节[6—7]。因此本研究于2021年7月, 分别在人工蓝藻结皮样地、2010年的人工植被区(柠条和油蒿混合种植)自然发育的蓝藻结皮和流沙区(样地基本概况见表 1), 采集0—2 cm土层样品约500g, 每个处理三个重复, 共采集样品21份。在实验室中对样品进行预处理, 剔除样品中的杂草, 自然风干后过1 mm筛, 用于测土壤酶。BSCs叶绿素a(Chlorophyll a, Chl-a)与胞外多糖(Exopolysaccharide, EPS)采样方法:用一个不锈钢圆形(内径7.08 mm)的器具随机取每个处理中的BSCs样品6份, 其中3份用于测叶绿素a, 另外3份用于测定胞外多糖。将采集的样品放入离心管中, 带回实验室进行测定。
| 样地 Sample plot |
生物土壤结皮特征 Characteristics of biological soil crusts |
主要草本植物物种 Main herb species |
植被盖度/% Vegetation coverage |
| 人工蓝藻结皮 Artificial cyanobacterial crust |
人工培育的蓝藻结皮, 盖度81.7%—85.6%, 厚度大约在2.0—2.5 mm左右 | 小画眉草Eragrostis poaeoides 砂蓝刺头Echinops gmelinii 虎尾草Chloris virgata 狗尾草Setaria viridis |
21.7 |
| 自然发育蓝藻结皮 Naturally-developing cyanobacterial crusts |
自然发育的蓝藻结皮总盖度在80.0%以上, 平均厚度为2.74 mm | 砂蓝刺头Echinops gmelinii 刺蓬Salsola tragus 小画眉Eragrostis poaeoides |
17.8 |
| 流沙Sand | 无生物土壤结皮覆盖 | 砂蓝刺头Echinops gmelinii 虫实Corispermum declinatum |
1.5 |
本研究选择了与土壤碳循环相关的4种土壤酶, 包括1种氧化还原酶和3种水解酶, 测定方法如下:土壤过氧氢化酶(Soil catalase, S-CAT)采用高锰酸钾滴定法测定, 土壤蔗糖酶(Soil sucrase, S-SC)、淀粉酶(Soil amylase, S-AL)、纤维素酶(Soil cellulase, S-CL)采用3, 5-二硝基水杨酸比色法测定[1]。
BSCs叶绿素a含量采用乙醇法测定, 上述样品带回实验室后向土壤中加入10 mL无水乙醇, 80℃水浴5min, 冷却30min后过滤。最后用分光光度计在665nm(A665)波长下测量提取溶液的吸光值, 计算公式[17]:

|
BSCs中的3种胞外多糖包括松散结合胞外多糖(LB-EPS)、紧密结合胞外多糖(TB-EPS)、糖被胞外多糖(G-EPS), 提取方法详见Rossi等[18], 每种胞外多糖含量测定采用苯酚-浓硫酸法[19]。
2.4 土壤酶恢复速率及恢复年限评价
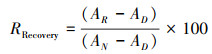
|
RRecovery是指人工蓝藻结皮相对于人工植被区自然发育的蓝藻结皮土壤酶活性的恢复速率, A指土壤酶活性, 下标R、N、D分别指人工蓝藻结皮、人工植被下自然发育的蓝藻结皮和流沙土壤酶活性[20]。
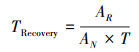
|
TRecovery是指接种3年后的人工蓝藻结皮土壤酶活性相对于自然发育蓝藻结皮的恢复时间, T指人工植被重建后的恢复时间, 本研究中为11年。
2.5 数据处理使用软件Excel 2020与SPSS 25.0进行数据处理和统计分析, 用Origin 2021作图。
3 结果与分析 3.1 人工蓝藻结皮的发育特征图 1显示, 不同处理的人工蓝藻结皮与人工植被区自然发育的蓝藻结皮在叶绿素a含量上无显著性差异(P>0.05), 人工蓝藻结皮分泌的3种胞外多糖含量有G-EPS>TB-EPS>LB-EPS的总趋势。LB-EPS和TB-EPS两种胞外多糖含量大小都表现为DPN>NWF>NWF+SSN>SSN>NWF+DPN>人工植被, 在LB-EPS方面, DPN覆盖方式显著高于其它处理(P<0.05), 在TB-EPS方面, DPN、NWF、NWF+SSN处理组显著高于其它处理(P<0.05);G-EPS含量大小表现为DPN>NWF+SSN>NWF>SSN>NWF+DPN>人工植被, 其中DPN和NWF+SSN显著高于其它处理(P<0.05)。从图 1中可以看出人工蓝藻结皮处理组的NWF、NWF+SSN、DPN的3种胞外多糖的总和显著高于人工植被下自然发育的蓝藻结皮(P<0.05)。

|
| 图 1 蓝藻结皮的发育特征 Fig. 1 Developmental characteristics of cyanobacterial crust NWF: 无纺布Non-woven fabric; DPN: 防尘网Dust-proof net; SSN: 遮阳网Sun-shading net; LB-EPS:松散结合胞外多糖loosely bound exopolysaccharide;TB-EPS:紧密结合胞外多糖tightly bound exopolysaccharide;G-EPS:糖被胞外多糖glycocalix exopolysaccharide; 平均值±标准误, 不同小写字母表示差异显著性, 显著水平为P<0.05 |
因此, 从两种指标来看, 接种3年后的人工蓝藻结皮在发育程度上已经超过自然发育时间为11年的蓝藻结皮, 说明人工蓝藻结皮技术可以缩短BSCs的发育时间。
3.2 人工蓝藻结皮对土壤酶活性及恢复速率的影响由图 2可知, 人工蓝藻结皮可以显著提高土壤表层蔗糖酶、过氧化氢酶以及纤维素酶活性(P<0.05)。其中人工蓝藻结皮处理组中土壤蔗糖酶活性为13.03—20.51 mg d-1 g-1, 与流沙相比, 提高了8.6—13.5倍;土壤纤维素酶活性在45.60—47.20 mg d-1 g-1之间, 是流沙的1.06—1.10倍;土壤过氧化氢酶活性为12.43—23.31 μmol d-1 g-1, 是流沙的1.8—3.3倍;人工蓝藻结皮下土壤淀粉酶与流沙相比, 虽然提高了淀粉酶活性1.4—2.4倍, 但只有DPN覆盖方式与流沙之间差异显著(P<0.05)。同时4种土壤酶中, 除了淀粉酶以外, 人工蓝藻结皮中的大多数处理组与自然发育的蓝藻结皮表层土壤酶活性无显著差异(P>0.05), 因此接下来我们计算了人工蓝藻结皮相对于自然发育蓝藻结皮的恢复速率以及恢复时间, 如表 2所示。

|
| 图 2 不同处理下土壤酶活性 Fig. 2 Soil enzyme activities under different treatments |
| 不同处理 Different treatment |
土壤蔗糖酶 Soil sucrase |
土壤纤维素酶 Soil cellulase |
土壤过氧化氢酶 Soil catalase |
土壤淀粉酶 Soil amylase |
|||||||
| 恢复速率/% Recovery rate |
恢复时间/a Recovery time |
恢复速率/% Recovery rate |
恢复时间/a Recovery time |
恢复速率/% Recovery rate |
恢复时间/a Recovery time |
恢复速率/% Recovery rate |
恢复时间/a Recovery time |
||||
| NWF+DPN | 80.2±3.2 | 9.01 | 109.5±8.8 | 11.08 | 77.7±1.8 | 9.27 | 23.6±20.0 | 4.81 | |||
| NWF | 118.1±9.1 | 12.82 | 91.9±4.2 | 10.93 | 96.7±15.3 | 10.74 | 15.3±6.5 | 4.13 | |||
| NWF+SSN | 117.8±9.8 | 12.79 | 70.1±2.3 | 10.74 | 58.6±11.6 | 7.80 | 21.9±1.1 | 4.67 | |||
| SSN | 71.6±6.4 | 8.14 | 114.0±7.1 | 11.12 | 31.9±4.7 | 5.73 | 14.5±12.1 | 4.07 | |||
| DPN | 108.9±7.9 | 11.89 | 98.4±15.0 | 10.99 | 43.7±5.4 | 6.65 | 49.1±11.2 | 4.88 | |||
| 平均值Mean | 99.3 | 10.93 | 100.0 | 10.97 | 61.7 | 8.04 | 24.9 | 4.91 | |||
| NWF, 无纺布Non-woven fabric; DPN, 防尘网Dust-proof net; SSN, 遮阳网Sun-shading net | |||||||||||
从表 2可以看出土壤蔗糖酶和纤维素酶在4种土壤酶中恢复速率最快, 前者在NWF、NWF+SSN、DPN覆盖处理上恢复速率已经超过人工植被(>100.0%), 总体平均值已达到99.3%, 后者在NWF+DPN和SSN覆盖方式上恢复速率也已超过人工植被(>100.0%), 并且总体平均值为100.0%;土壤过氧化氢酶恢复速率次之, 平均恢复速率为61.7%;土壤淀粉酶恢复速率最慢, 均低于50.0%, 除了DPN覆盖方式的恢复速率为49.1%, 其余处理组的恢复速率在14.5%—23.6%之间, 总体恢复速率平均值为24.9%。同时将接种培育3年的人工蓝藻结皮与建植11年的自然发育的蓝藻结皮表层土壤酶相比, 结果表明, 恢复速率较快的两种土壤酶已达到自然发育蓝藻结皮11年左右的水平, 后两种土壤酶也分别达到了自然发育蓝藻结皮8年和5年左右的水平。由此可见, 人工蓝藻结皮可以加快表层土壤酶活性的恢复, 与自然发育的蓝藻结皮相比缩短了恢复时间。
3.3 人工蓝藻结皮基本特征与土壤酶活性相关性为了揭示4种土壤酶与人工蓝藻结皮基本特征之间的关系, 我们将4种土壤酶活性与叶绿素a、LB-EPS、TB-EPS、G-EPS含量进行了回归分析(图 3)。

|
| 图 3 人工蓝藻结皮基本特征(Chl a、LB-EPS、TB-EPS、G-EPS)与土壤酶活性的回归分析 Fig. 3 Regression analysis of artificial cyanobacterial crust properties(Chlorophyll a, LB-EPS, TB-EPS, G-EPS)and soil enzymes activity 显著水平为P<0.05 |
由图 3可知, 土壤蔗糖酶和纤维素酶活性与人工蓝藻结皮中叶绿素a的含量呈显著正相关(R2=0.366, P<0.05;R2=0.683, P<0.05), 土壤过氧化氢酶与淀粉酶与叶绿素a的含量呈正相关但不显著(R2=0.106, P>0.05;R2=0.112, P>0.05);在胞外多糖方面, 4种酶与LB-EPS、TB-EPS、G-EPS均显著线性正相关(P<0.05), 其中蔗糖酶与3种胞外多糖的相关性最强, R2分别为0.812、0.802、0.768。由此可见, 土壤酶的活性随着人工蓝藻结皮的发育逐渐增强, 并且恢复速率较快的土壤蔗糖酶和纤维素酶与人工蓝藻结皮基本特征的关系更加显著。
4 讨论BSCs组成中的一些生物体不仅可以通过光合作用固定碳, 还能够分泌胞外多糖从而增加土壤碳库储量[3]。因此, 本文选取与土壤碳循环相关的4种土壤酶, 其中土壤蔗糖酶、纤维素酶、淀粉酶为水解酶类, 可以将土壤中的有机质分解转化, 为植物和微生物提供养分和能量, 土壤过氧化氢酶为氧化酶类, 可以促进过氧化氢的分解, 消除其对生物体的毒害作用, 同时也与土壤有机质含量也有关[1—2]。本文研究结果显示发育早期的人工蓝藻结皮能够提高表层土壤酶的活性, 但不同的处理以及土壤酶种类之间存在差异, 其中以土壤蔗糖酶与流沙差异最大, 是流沙的8.6—13.5倍。人工蓝藻结皮增加土壤酶活性可以从三个方面进行分析:1)BSCs通过增加土壤微生物生物量以及促进植被生长进而增加土壤酶的来源[21—22];2)大量研究表明土壤酶与土壤养分具有较强的相关性, 而BSCs可以增加土壤C、N、P的含量[23];3)BSCs还可以改变土壤结构以及土壤周围的环境条件如土壤水分、土壤温湿度等, 从而有利于对土壤酶的吸附和酶促反应的发生[24]。
已有研究表明, 随着BSCs的发育, 表层土壤酶的活性逐渐增强, 可以归因于BSCs的发育改变了土壤理化性质并且对植被产生影响, 此时形成的BSCs—土壤—植被系统相互影响相互作用, 有利于土壤酶活性的增加[24—25]。王凯等[9]在腾格里沙漠东南缘也发现BSCs的基本特征与土壤酶活性显著相关, 我们的研究同样支持BSCs的发育特征与土壤酶活性相关, 本研究表明恢复速率较快的土壤蔗糖酶和纤维素酶与叶绿素a呈显著线性正相关。叶绿素a含量表示人工BSCs所包含的藻类生物量, 而这些藻类可以分泌多种胞外酶, 因此, 随着叶绿素a含量的提高, 土壤蔗糖酶和纤维素酶的活性也增加, Xiao等[21]也发现接种人工藓类结皮后可以增加土壤微生物生物量, 从而有利于土壤酶的增加[26]。将3种胞外多糖与土壤酶进行回归分析发现, 4种土壤酶与LB-EPS、TB-EPS、G-EPS都呈显著线性正相关, Zhang等[27]研究表明表层土壤酶的活性与土壤微生物生物量、胞外多糖和有机碳有关, 一方面, 胞外多糖是土壤碳库的重要来源, 是土壤酶发生催化反应的底物[28];另一方面, BSCs分泌的胞外多糖有利于水分的保留, 增加土壤表层水分, 促进土壤酶活性的保持与酶促反应的发生[18], 因此随着BSCs分泌的胞外多糖含量增多, 土壤酶的活性也逐渐提高。由此可以看出, 人工蓝藻结皮基本特征与土壤酶的关系和自然发育的BSCs规律一致[9], 两者具有很好的相关关系, 人工蓝藻结皮的发育能够促进表层土壤酶活性的增加。
本研究中, 除了土壤淀粉酶外, 人工蓝藻结皮表层土壤蔗糖酶、纤维素酶、过氧化氢酶与人工植被区无显著差异(P>0.05), 并且前两种酶的活性已达到了自然发育的蓝藻结皮表层土壤酶活性11年左右的水平, 土壤过氧化氢酶也达到了自然发育的蓝藻结皮表层土壤酶活性8年左右的恢复水平, 这说明人工蓝藻结皮的应用与人工植被的建植相比, 可以加快土壤酶活性的恢复。4种土壤酶的恢复速率也具有一样的趋势, 总体来看, 土壤蔗糖酶和纤维素酶恢复速率较快, 土壤过氧化氢酶次之, 土壤淀粉酶恢复速率最慢, 这与张鹏等[29]的研究结果一致, 其通过对人工植被重建后不同植被配置以及演替阶段的土壤酶调查发现, 总体上水解类的土壤酶恢复速率高于氧化还原酶类。但在本研究的同一地区也有相反的结论, Zhang等[20]通过对建立50多年的人工植被下的土壤酶研究发现氧化还原酶类恢复速率较水解酶类快, 并且研究表明氧化酶类的恢复与灌木相关;而水解酶类与草本植物和BSCs联系密切, 前者木质素含量较低, 可以增加土壤微生物基质有效性, 后者通过分泌胞外多糖等底物而改变基质养分的有效性。人工BSCs接种后前几年基本上是以草本植物和BSCs为主[30], 在本实验样地也具有一样的植物生活型特征, 因此在本实验中水解酶类的恢复速率快于氧化还原酶类。而土壤淀粉酶恢复速率最低的原因可能也与人工蓝藻结皮试验区的植物以草本植物为主有关, 植物的根系分泌物也是干旱地区土壤酶的来源之一, 根据植物种类的不同, 土壤酶的活性也会有所差异[31], 此时的人工植被以灌木为主, 两者地下部分的根系不同, 分泌的酶也具有差异, 因此淀粉酶活性较低, 恢复速率较慢。
5 结论综上所述, 人工蓝藻结皮的拓殖与发育可以显著增加表层土壤蔗糖酶、纤维素酶、淀粉酶和过氧化氢酶活性, 且与人工蓝藻结皮的基本特征显著相关。与自然发育的蓝藻结皮相比, 人工蓝藻结皮的拓殖与发育加快了土壤酶的恢复速率, 恢复速率较快的土壤蔗糖酶和纤维素酶在短期内达到了自然发育蓝藻结皮11年的水平, 缩短了土壤酶的恢复时间, 促进了荒漠土壤生态系统功能的恢复。本文通过测定与碳相关的4种土壤酶活性来评价人工蓝藻结皮对沙化土地的贡献, 并且通过与已知年限的自然发育蓝藻结皮比较, 量化了土壤酶的恢复时间, 为人工蓝藻结皮在沙区大规模推广提供了支撑。
但人工蓝藻结皮只是目前人工BSCs的其中一种, 不同的BSCs类型会对土壤酶活性产生不同的影响, 因此今后可开展人工地衣结皮和人工藓类结皮对土壤酶活性的影响, 并且本文仅考虑了人工蓝藻结皮基本特征对表层土壤酶活性恢复的影响, 没有涉及土壤微生物以及土壤水分、pH和电导、土壤碳氮磷含量等方面, 如果综合考虑这些因素, 并对土壤酶活性进行季节性的监测, 将有助于更好的理解人工BSCs对土壤功能恢复的影响。
| [1] |
关松荫. 土壤酶及其研究法. 北京: 农业出版社, 1986.
|
| [2] |
周礼恺, 张志明, 曹承绵. 土壤酶活性的总体在评价土壤肥力水平中的作用. 土壤学报, 1983, 20(4): 413-418. |
| [3] |
Belnap J, Lange O L. Biological Soil Crusts: Structure, Function, and Management. Heidelberg: Springer, 2003.
|
| [4] |
李新荣, 谭会娟, 回嵘, 赵洋, 黄磊, 贾荣亮, 宋光. 中国荒漠与沙地生物土壤结皮研究. 科学通报, 2018, 63(23): 2320-2334. |
| [5] |
樊瑾, 李诗瑶, 余海龙, 黄菊莹. 毛乌素沙地不同类型生物结皮与下层土壤酶活性及土壤碳氮磷化学计量特征. 中国沙漠, 2021, 41(4): 109-120. |
| [6] |
Liu Y M, Yang H Y, Li X R, Xing Z S. Effects of biological soil crusts on soil enzyme activities in revegetated areas of the Tengger Desert, China. Applied Soil Ecology, 2014, 80: 6-14. DOI:10.1016/j.apsoil.2014.03.015 |
| [7] |
Zhang B C, Zhou X B, Zhang Y M. Responses of microbial activities and soil physical-chemical properties to the successional process of biological soil crusts in the Gurbantunggut Desert, Xinjiang. Journal of Arid Land, 2015, 7(1): 101-109. DOI:10.1007/s40333-014-0035-3 |
| [8] |
Miralles I, Domingo F, Cantón Y, Trasar-Cepeda C, Leirós M C, Gil-Sotres F. Hydrolase enzyme activities in a successional gradient of biological soil crusts in arid and semi-arid zones. Soil Biology and Biochemistry, 2012, 53: 124-132. DOI:10.1016/j.soilbio.2012.05.016 |
| [9] |
王凯, 厉萌萌, 刘德权, 张凌基, 刘可欣, 许华, 唐亮, 何明珠, 张珂. 腾格里沙漠不同组成生物结皮特征及其对土壤酶活性的影响. 生态学报, 2022, 42(14): 5859-5868. |
| [10] |
李新荣, 回嵘, 赵洋. 中国荒漠生物土壤结皮生态生理学研究. 北京: 高等教育出版社, 2016.
|
| [11] |
Rossi F, Li H, Liu Y D, De Philippis R. Cyanobacterial inoculation (cyanobacterisation): perspectives for the development of a standardized multifunctional technology for soil fertilization and desertification reversal. Earth-Science Reviews, 2017, 171: 28-43. DOI:10.1016/j.earscirev.2017.05.006 |
| [12] |
Zhao Y, Jia R L, Wang J. Towards stopping land degradation in drylands: water-saving techniques for cultivating biocrusts in situ. Land Degradation & Development, 2019, 30(18): 2336-2346. |
| [13] |
Zhou X B, Zhao Y G, Belnap J, Zhang B C, Bu C F, Zhang Y M. Practices of biological soil crust rehabilitation in China: experiences and challenges. Restoration Ecology, 2020, 28(S2): S45-S55. |
| [14] |
唐东山, 王伟波, 李敦海, 胡春香, 刘永定. 人工藻结皮对库布齐沙地土壤酶活性的影响. 水生生物学报, 2007, 31(3): 339-344. |
| [15] |
吴易雯, 饶本强, 刘永定, 李根保, 李敦海. 不同生境对人工结皮发育及表土氮、磷含量及其代谢酶活性的影响. 土壤, 2013, 45(1): 52-59. |
| [16] |
李新荣, 张志山, 刘玉冰, 李小军, 杨昊天. 中国沙区生态重建与恢复的生态水文学基础. 北京: 科学出版社, 2016.
|
| [17] |
Zhao Y, Wang N, Zhang Z S, Pan Y X, Jia R L. Accelerating the development of artificial biocrusts using covers for restoration of degraded land in dryland ecosystems. Land Degradation & Development, 2021, 32(1): 285-295. |
| [18] |
Rossi F, Mugnai G, De Philippis R. Complex role of the polymeric matrix in biological soil crusts. Plant and Soil, 2018, 429(1/2): 19-34. |
| [19] |
Dubois M, Gilles K A, Hamilton J K, Rebers P A, Smith F. Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 1956, 28(3): 350-356. |
| [20] |
Zhang Z S, Dong X J, Liu Y B, Li X R, Jia R L, Hu Y G, He M Z, Huang L. Soil oxidases recovered faster than hydrolases in a 50-year chronosequence of desert revegetation. Plant and Soil, 2012, 358(1/2): 275-287. |
| [21] |
Xiao B, Veste M. Moss-dominated biocrusts increase soil microbial abundance and community diversity and improve soil fertility in semi-arid climates on the Loess Plateau of China. Applied Soil Ecology, 2017, 117-118: 165-177. |
| [22] |
杨航宇, 刘艳梅, 王廷璞, 回嵘. 生物土壤结皮对荒漠区土壤微生物数量和活性的影响. 中国沙漠, 2017, 37(5): 950-960. |
| [23] |
高丽倩, 赵允格, 许明祥, 孙会, 杨巧云. 生物土壤结皮演替对土壤生态化学计量特征的影响. 生态学报, 2018, 38(2): 678-688. |
| [24] |
Niu J P, Yang K, Tang Z J, Wang Y T. Relationships between soil crust development and soil properties in the desert region of north China. Sustainability, 2017, 9(5): 725. |
| [25] |
吴永胜, 尹瑞平, 田秀民, 哈斯. 毛乌素沙地南缘人工植被区生物结皮发育特征. 中国沙漠, 2018, 38(2): 339-344. |
| [26] |
Ghiloufi W, Seo J, Kim J, Chaieb M, Kang H. Effects of biological soil crusts on enzyme activities and microbial community in soils of an arid ecosystem. Microbial Ecology, 2019, 77(1): 201-216. |
| [27] |
Zhang K, Li M M, Wang K, Gao N, Liu D Q, Chen Y L. Depth-related response of soil enzymes to cyanobacteria-dominated crusts along a precipitation gradient. Land Degradation & Development, 2021, 32(15): 4183-4192. |
| [28] |
Hernández D L, Hobbie S E. The effects of substrate composition, quantity, and diversity on microbial activity. Plant and Soil, 2010, 335(1/2): 397-411. |
| [29] |
张鹏, 赵洋, 黄磊, 胡宜刚, 韩旭. 植被重建对露天煤矿排土场土壤酶活性的影响. 生态学报, 2016, 36(9): 2715-2723. |
| [30] |
Lan S B, Zhang Q Y, Wu L, Liu Y D, Zhang D L, Hu C X. Artificially accelerating the reversal of desertification: cyanobacterial inoculation facilitates the succession of vegetation communities. Environmental Science & Technology, 2014, 48(1): 307-315. |
| [31] |
杨文彬, 耿玉清, 王冬梅. 漓江水陆交错带不同植被类型的土壤酶活性. 生态学报, 2015, 35(14): 4604-4612. |