 2023, Vol. 43
2023, Vol. 43文章信息
- 王菲, 曹永强, 周姝含, 范帅邦, 姜雪梅
- WANG Fei, CAO Yongqiang, ZHOU Shuhan, FAN Shuaibang, JIANG Xuemei
- 黄河流域生态功能区植被碳汇估算及其气候影响要素
- Estimation of vegetation carbon sink in the Yellow River Basin ecological function area and analysis of its main meteorological elements
- 生态学报. 2023, 43(6): 2501-2514
- Acta Ecologica Sinica. 2023, 43(6): 2501-2514
- http://dx.doi.org/10.5846/stxb202203110591
-
文章历史
- 收稿日期: 2022-03-11
- 网络出版日期: 2022-11-03





2. 天津师范大学京津冀生态文明发展研究院, 天津 300387;
3. 东北财经大学公共管理学院, 大连 116025
2. Institute of Beijing-Tianjin-Hebei Ecological Civilization Development, Tianjin Normal University, Tianjin 300387, China;
3. School of Public Administration, Dongbei University of Finance and Economics, Dalian 116025, China
政府间气候变化专门委员会(IPCC)第六次评估报告指出:气候变暖造成陆地生态系统前所未有的变化:物种的灭亡、频繁的火灾以及植被碳储量的减少极易造成陆地碳的大量释放, 从而引发不断加剧的恶性循环。陆地生态系统碳循环系统作为全球碳循环的重要组成部分, 其沟通了大气圈、水圈和土壤圈, 是区域气候特征和人类活动的反映和指示[1—2]。目前, 学者们已对不同区域(世界[3]、北半球[4]、中国[5]、流域[6]、省份[7])的主要碳库(森林[8]、草地[9]、农田[10—11])的固碳大小[12]、碳源/汇属性[13]及其对气候变化的响应[14]进行了探究。然而, 陆地生态系统与气候变化是一个复杂的互馈过程, 如何在全球变暖、极端气候频率加快及人类活动强度增加的背景下, 揭示区域碳循环的过程机制、变化特征、归因分析, 是目前面临的一大难题。
中国陆地生态系统固碳潜力巨大, 每年CO2吸收量占全球的8%—11%[15—16], 但具体到中国各区域而言, 植被固碳能力大小不一且应对气候变化反馈机制不同, 导致区域间生态系统固碳量的差异。黄河流域是我国生态文明建设的主战场, 承担着重要的生态屏障功能, 但近年来, 随着经济、人口的进一步发展, 黄河流域生态破环成为制约流域可持续发展的瓶颈要素[17], 同时, 黄河流域半湿润半干旱的大陆性气候决定了其生态环境自我调节能力有限。目前, 有关黄河流域森林碳储量[18]、植被归一化指数(Normalized Difference Vegetation Index, NDVI)[19]、植被净初级生产力(Net Primary Productivity, NPP)[20]的研究较为丰富, 大致掌握了该流域植被生长状况, 但进一步探究植被碳源/汇的研究相对较少, 存在流域尺度碳源/汇的属性界定不清楚、生态功能区固碳能力估算结果的不统一、植被对气候变化响应不一致等情况, 且大多是对整个流域或分市县进行, 该划分方式破坏了生态系统的完整性。因此, 有必要选择适宜的模型对该流域进行碳储量估算及碳源/汇的确定。
本文以生态功能区与生态系统碳循环的紧密联系为切入点, 探究朱文泉教授改进后的CASA模型及土壤微生物呼吸量模型在黄河流域的适用性, 并在此基础上, 对黄河流域各生态功能区碳源/汇进行估算, 进而确定了该地区的碳汇属性及与主要气象要素的关系。以期为生态系统管理部门制定科学合理的土地利用决策提供参考依据。
1 材料与方法 1.1 研究区概况黄河干流全长5464 km, 流域总面积79.5万km2(含内流区面积4.2万km2), 位于95°53′45″—119°12′48″E、32°9′33″—41°50′20″N。区域内气候的差异性显著, 大部分属于干旱半干旱区, 年日照时数2000—3000 h, 季节温差明显, 降水年际变化大, 年均蒸发量1100 mm, 多冰雹、沙暴等天气, 地势西高东低。黄河是世界含沙量最大的河流, 流域内水土流失问题严重, 生态恢复工程方兴未艾。根据中国植被类型空间分布数据显示, 黄河流域东部的广大地区主要植被类型为草地, 占流域总面积的55%以上, 碳储量潜力巨大, 林地主要集中在黄河上游水源涵养区、秦岭等山地地区, 耕地主要分布于中下游华北平原区, 流域森林面积约9.52万km2, 植被碳循环对气候变化反应极为敏感。黄河流域生态区涉及到中国3大生态大区(Ⅰ:东部季风大区、Ⅱ:西部干旱大区、Ⅲ:青藏高寒生态大区)内的11个生态区(图 1)。

|
| 图 1 黄河流域生态功能区划方案 Fig. 1 Ecological function zoning scheme of the Yellow River Basin |
(1) NEP估算模型
净生态系统生产力(Net Ecosystem Productivity, NEP)是指净初级生产力(Net Primary Productivity, NPP)中减去异养生物呼吸损耗(土壤呼吸)光合产物(Heterotrophic Respiration, Rh)之后的部分, 即植被的固碳量[21]。其公式为:
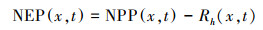
|
(1) |
式中, NEP(x, t)表示像元x在t月的植被净生态系统生产力(gC/m2), NPP(x, t)表示像元x在t月的植被净初级生产力(gC/m2), Rh(x, t)表示像元x在t月的土壤微生物呼吸量(gC/m2)。当NEP>0时, 被固定的碳量多于异养呼吸消耗的碳量, 此生态系统为碳汇;当NEP < 0时, 表示植被固定的碳量少于异养呼吸消耗的碳量, 此生态系统为碳源。NPP是指绿色植被在扣除自身呼吸消耗以外, 单位面积单位时间内所累积的有机物的数量[22]。本文通过朱文泉等[23—24]改进的CASA模型模拟植被生长状况, 该模型充分考虑了植被生长特性和环境条件, 能够更为准确的判定研究区生态系统的碳源/汇属性。其表达式为:
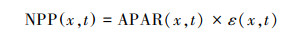
|
(2) |
式中, APAR(x, t)表示x像元在t月吸收的光合有效辐射(gC m-2 month-1), ε(x, t)表示像元x在t月的光能利用率(gC/MJ)。
对土壤微生物呼吸量的确定是估算区域碳源/汇的关键。国内外学者大多借助机理模型或经验模型拟合土壤微生物呼吸量(Rh)与土壤呼吸量(Rs)值的回归关系模型, Bond_Lamberty和Raich等[25—27]基于全球森林土壤特质提出了Rh与Rs的关系公式;石志华[28]推导出适合陕西省Rh与Rs的月尺度关系, 但具有较强的区域性。裴志永等[29]基于青海省五道梁实验点实测数据建立了气温、降水与碳排放的土壤呼吸回归模型, 并在西北干旱区[30]、甘肃省[31]、青海高原[32]等地区应用取得理想效果。鉴于本文研究区面积较大, 目前尚未建立普适模型, 在综合对比三种模型结果基础上, 本文选用裴志永等建立的关系模型计算土壤异养呼吸Rh, 该模型公式如下:
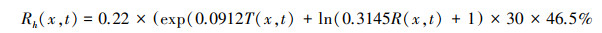
|
(3) |
式中, T(x, t)为像元x在第t个月的平均气温(℃);R(x, t)表示像元x在第t个月的月平均降水量(mm)。相关公式及参数见参考文献[33—34]。
(2) 多元回归模型
为探究多个变量对NEP的相对贡献率, 利用多元回归模型对2000—2020年NEP与年均气温、年累计降水和年总辐射量进行回归计算:
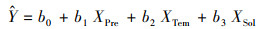
|
(4) |
式中, Xpre, XTem, XSol分别表示降水, 气温和总辐射的时间序列样本, 再将回归系数进行标准化处理, 首先对NEP、降水、气温和总辐射求取标准差, 以降水为例:
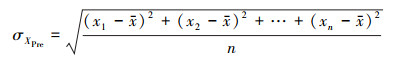
|
(5) |
而后对系数进行标准化:
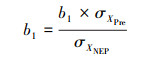
|
(6) |
(3) 本文采用一元线性回归法分析NPP年际变化率[35];利用ArcGIS栅格计算器计算出NEP与气象要素的空间相关性, 并进行显著性检验[36]。
1.3 数据来源本文归一化植被指数(NDVI)来自于中国科学院资源环境科学数据中心(http://www.resdc.cn)提供的1 km数据集;植被类型数据为寒区旱区科学数据中心的《基于多源数据融合方法的中国1公里土地覆盖图》, 该数据集在中国地区的分类准确度超过70%, 对中国地区涉及植被类型研究的课题具有较高的参考价值;本文将植被类型归纳为9类用地:耕地、林地、草地、建筑用地、水域、沼泽地、裸沙地、戈壁、风蚀劣地[34];月均温、月降水量和月总太阳辐射数据来源于中国气象数据网(http://data.cma.cn/), 时间尺度为2000—2020年, 为保证空间插值精度, 选取35个流域及周围站点(图 2), 其中气温和降水的插值应用澳大利亚的ANUSPLIN软件, 太阳辐射数据选用反距离权重法进行插值[37], 因本文研究区较大, 故将所有数据重采样统一为1 km分辨率。生态功能区划方案采用中国生态系统评估与生态安全数据库[32](http://www.ecosystem.csdb.cn/), 将黄河流域分为11个生态分区。

|
| 图 2 黄河流域土地利用及监测站点分布 Fig. 2 Distribution of land use and monitoring stations in the Yellow River Basin |
本文通过分别验证NPP和Rh两个参数结果, 进而判断区域NEP估算结果的科学性与准确性。一般来说, 估算值验证的方式有:与实测值验证、与其它研究成果对比、与其它遥感数据对比等[31], 本文因研究区较广, 大面积实测数据获取难度非常困难, 故选取与其他研究成果和与其他遥感产品进行对比的方式验证估算值。
2.1.1 NPP模拟结果验证为保证模型结果的准确性, 对CASA模型模拟的NPP值与MOD17A3 NPP产品值进行验证对比(图 3), 结果显示本文模拟的NPP值与MOD17A3 NPP产品值拟合优度R2均大于0.7(P < 0.01), 具有一定的统计学意义;同时本文模拟黄河流域NPP年均值为317.18 gC m-2 a-1, 该结果与王娟等[38]对黄河流域NPP值模拟结果相似(多年平均值281.39 gC m-2 a-1), 略高于张丹丹[39](228.2 gC m-2 a-1)的模拟结果, 这可能是研究年限不同造成;此外, 本文年均NPP最高值出现在2018年, 朱莹莹[40]对黄河流域的模拟最高值出现在2019年, 变化趋势相一致。因此认为本文模拟结果具有较高可靠性。

|
| 图 3 黄河流域模拟NPP值与MODIS NPP值对比 Fig. 3 Comparison of simulated NPP values and MODIS NPP values in the Yellow River Basin NPP:植被净初级生产力Net primary productivity |
黄河流域土壤微生物呼吸量估算结果与其他研究结果对比见表 1, 本文月均Rh估算值(3.44—30.89 gC m-2 a-1)与刘春雨[31]、牟银娟[41]应用裴志永公式对甘肃省、石羊河流域土壤微生物呼吸量估算结果相一致, 但明显低于采用Bond_Lamberty和Raich公式(30.15—82.99 gC m-2 a-1)和田琴等[42]实测数据(97.88 gC m-2 a-1)估算的结果, 这可能因为研究区土壤特性差别过大所致。研究年段、气象数据的插值处理以及模型参数的调整、实验误差等都会导致土壤微生物呼吸量计算结果的差异, 总体来说, 可以认为采用裴志永公式估算的黄河流域土壤呼吸量结果基本可靠。
| 区域 Area |
时间 Time |
月均Rh/(gC m-2 a-1) Monthly average |
方法 Method |
| 甘肃省[31]Gansu province | 2000—2010年 | 15.19—28.00 | 裴志永公式[29] |
| 石羊河流域[41]Shiyang river Basin | 2000—2015年 | 11.71—38.47 | 裴志永公式 |
| 河西走廊[33]Hexi corridor | 2001—2016年 | 30.15—82.99 | Bond_Lamberty和Raich公式 |
| 黄土丘陵区[42Loess hilly area | 2013年8月 | 97.88 | 实测数据 |
| 本文This study | 2000—2020年 | 3.44—30.89 | 裴志永公式 |
| Rh:土壤微生物呼吸量Heterotrophic respiration | |||
根据CASA模型模拟黄河流域2000—2020年植被NPP变化趋势见图 4, 黄河流域多年平均NPP值为317.18 gC m-2 a-1, 年际NPP整体呈波动上升趋势, 表明黄河流域植被状况得到不断改善。上升趋势主要可以分为两个阶段, 第一阶段为2000—2011年低水平大幅度波动, 24.848/10a的速度上升, 2000年为近18年的最低值(260.39 gC m-2 a-1), 第二阶段为高水平小幅度波动, 自2011年开始植被NPP出现飞跃增长, 此后呈现持续波动状态, 变化速度为18.128/10a, 十年间仅2017年低于平均水平, 2018年达到最高值, 为356.65 gC m-2 a-1。自2010年开始我国低碳试点城市建设持续推进, “三北”防护林、“天然林保护工程”、“退耕还林”等多项生态修复工程取得显著成效, 有力提高了固碳减排能力[17]。黄河流域不同生态功能区植被NPP差异明显, 秦巴山地落叶与常绿阔叶林生态区年均NPP值最高, 为576.55 gC m-2 a-1, 其次为燕山—太行山山地落叶阔叶林生态区(506.37 gC m-2 a-1), 内蒙古高原中部草原化荒漠生态区NPP值最低, 为88.78 gC m-2 a-1。

|
| 图 4 2000—2020年黄河流域NPP变化趋势 Fig. 4 Change trend of NPP in the Yellow River Basin from 2000 to 2020 |
黄河流域不同植被类型NPP差异显著(图 5), 其中, 林地>耕地>沼泽>草地, 水体、戈壁、风蚀劣地和裸沙地的NPP总体处于较低值;NPP四季变化为:夏季>秋季>春季>冬季, 夏季NPP整体处于高位, 8月份最高, 为58.07 gC m-2 a-1;春季和秋季NPP值相差无几;冬季NPP年均值最低, 极低值出现在1月份, 为5.548 gC m-2 a-1, 这与植被生长规律相一致。在3—4月, 随着气温逐渐回升, 降水逐渐增加, 植物开始萌芽, 初级生产力呈上升趋势;5—9月气温大幅回升降水迅速增加, 在这种条件下植被的水热耦合作用达到了最佳组合, 是植被生长的最佳时期, 植被初级生产力达到最高值。10—11月气温回落, 降水减少, 自然植被生长缓慢, 且绝大多数农作物都已收割, 12月为一年中气温最低时期, 植被处于枯萎期, 植被生产能力弱。

|
| 图 5 黄河流域季节及不同植被类型NPP变化趋势 Fig. 5 Variation trend of monthly NPP in the Yellow River Basin |
黄河流域植被NPP受水热组合条件的限制, 空间差异显著, 呈现出南高北低, 平原、山地高, 高原、盆地低的分布特点(图 6)。NPP高值区集中在黄土高原农业与草原生态区(Ⅰ—12), 素有“塞上粮仓”、“塞北江南”的河套平原土壤肥沃, 灌溉系统发达, 农作物覆盖率高, NPP值相对较高;汾渭盆地农业生态区(Ⅰ—11)的秦岭北麓沿线、陕西中部渭北高原区以及东部接壤的山西省地区, 随着废弃露天矿山生态修复项目、汾河中上游山水林田湖草生态保护修复工程基本完成[43], 植被NPP值普遍提高;辽东-山东丘陵落叶阔叶林生态区(Ⅰ—8)植被状态良好, NPP值较高;在内蒙古西部大区中(Ⅱ)由于受到海陆分布和降水条件的限制, 植被主要以草甸及荒漠为主, 土壤贫瘠, 自然生态环境恶劣, 植被覆盖率低[41]。黄河流域植被NPP在2000—2020年约有47.13%的区域NPP呈增加趋势, 其中, 极显著增加面积占35.86%;无显著变化的区域占总面积的1/2, NPP减少的区域占比很小, 仅有3.2%。增加区域主要位于黄土高原农业与草原生态区(Ⅰ—12), 该生态区是我国“三北防护林”建设的主要实施地, 以政府为主导的生态恢复制度效果显著, 森林固碳效果明显。

|
| 图 6 2000—2020年黄河流域NPP空间分布变化 Fig. 6 Spatial variation of NPP in the Yellow River Basin from 2000 to 2020 |
通过计算2000—2020年黄河流域逐年和逐月Rh平均值, 得到本研究区21年土壤微生物呼吸量值的变化曲线(图 7)。由图 7黄河流域Rh年均值可知, 21年来黄河流域Rh极大值出现2018年, 为189.90 gC m-2 a-1, 这与NPP最大值年份一致, 生物生产力的提高导致土壤微生物的数量激增, 最小值出现在2004年, 为172.06 gC m-2 a-1, 极差为17.84 gC m-2 a-1。研究区Rh值大致可以分为两个阶段, 第一阶段是2000—2013年, 整体Rh值较低, 以0.1507 gC m-2 a-1速度微弱增长, 但波动较大;第二阶段为2013—2020年, 整体Rh值较高, 以0.6448 gC m-2 a-1的速度增长。总体而言, 黄河流域Rh呈显著增加趋势, 年总Rh2000年174.915 gC/a增加到2020年的181.335 gC/a。

|
| 图 7 2000—2020年黄河流域Rh时间变化趋势 Fig. 7 Temporal variation trend of Rh in the Yellow River Basin from 2000 to 2020 |
黄河流域月Rh年内变化呈单峰状态, Rh分层比较明显, 从2月开始Rh呈上升趋势, 其中5—9月Rh总量占到了全年总量的70.123%, 7月Rh值为最高, 达到30.885 gC m-2 a-1;冬季(12月—次年2月)Rh整体偏低, 12月达到最低值3.435 gC m-2 a-1。Rh最大值变化与均值变化一致, 最大值出现在7月, 达56.171 gC m-2 a-1, 超过了同月NPP均值, 出现土壤微生物呼吸量大于植被净生产能力的“超负荷”现象。
由2000—2020年黄河流域Rh年均值空间分布图(图 8)可知, 黄河流域Rh总体上呈现从东南往西北、从东往西依次递减的空间分布格局。整体而言, Rh高值主要分布在汾渭盆地农业生态区(Ⅰ—11), 陕西省和山西省秦岭沿线土壤微生物呼吸量最高, 该地区森林茂密, 气候温湿, 生物量丰富。

|
| 图 8 2000—2020年黄河流域Rh空间变化趋势 Fig. 8 Spatial variation trend of Rh in the Yellow River Basin from 2000 to 2020 |
其次是华北平原农业生态区(Ⅰ—13)及辽东-山东丘陵落叶阔叶林生态区(Ⅰ—8), 华北平原是我国重要的农业生产区, 地表农作物覆盖率高;位于黄河三角洲的山东省东营市近些年来致力于打造“国际湿地城市”, 截止2020年, 该地区通过蓄淡压碱修复退化湿地近200 km2, 以生态补偿方式完成退耕还湿面积140 km2[44];土壤微生物呼吸量低值区主要位于海拔较高的祁连山森林与高寒草原生态区(Ⅲ—1)和江河源区-甘南高寒草甸草原生态区(Ⅲ—4)。黄河流域在2000—2020年93%的区域Rh值呈增加趋势, 尤其在内蒙古高原中部-陇中荒漠草原生态区(Ⅱ—2)的宁夏地区, 增长趋势明显, 主要是由于该地区近些年来实施了多项生态修复政策, 主要有针对贺兰山区域的贺兰山生态保护与修复重点工程[45], 针对“十年九旱, 生态焦黄”西海固地区的退耕还林、荒山造林[46]、生态移民[47]等多项生态工程;Rh值趋于减少的区域仅出现在黄土高原农业与草原生态区(Ⅰ—12)的关中平原地区以及内蒙古高原中东部典型草原生态区(Ⅱ—1)的鄂尔多斯高原[48], 该地区是中国最典型的生态敏感区, 水资源短缺, 土地沙漠化, 加之人类活动频繁, 是我国重要的能源化工基地[49], 导致该地区生态环境极端脆弱。
2.4 黄河流域碳源/汇估算结果分析在2000—2020年, 黄河流域呈碳汇属性的区域约占研究区面积的60%以上(图 9), 分布广泛, 并随着年份的推移, 呈碳汇属性的面积逐渐增大, 由2000年的64.22%增长到2020年的76.32%, 植被碳汇能力明显增强, 碳汇区年固碳量均值197.13 gC m-2 a-1, 年均碳汇量121.37 MgC/a;增长趋势以黄土高原农业与草原生态区(Ⅰ—12)为中心向北向东北方向扩张, 该地区是秦岭生态保护和修复工程及黄土高原水土流失综合治理工程重点区域。碳源区域呈不断缩小趋势, 主要集中在内蒙古高原中部-陇中荒漠草原生态区(Ⅱ—2)和内蒙古高原中部草原化荒漠生态区(Ⅱ—3), 陇中地区主要为黄河高原水土流失区, 内蒙古中南部毛乌素沙地沙漠化严重[50]。在2000—2020年黄河流域年均NEP值为137.83 gC m-2 a-1, 年均固碳量约为111.02 MgC/a;年排碳量均值为-52.47 gC m-2 a-1, 年均排碳量为-9.96 MgC/a, 年均净碳汇总量约为111.41 MgC/a。21年间黄河流域植被累积NEP平均值为2857.17 gC m-2 a-1, 年均固碳量约为2301.33 MgC/a。

|
| 图 9 黄河流域碳源/汇空间分布 Fig. 9 Spatial distribution of carbon source/sink in the Yellow River Basin |
总体来看, 黄河流域主要呈碳汇属性, 但碳汇区域存在不稳定性, 这可能与该流域土地利用变化及林地面积不足(12%)有关, 该研究区土地利用类型以草地和耕地为主, 相较于林地, 草地与耕地更易受到气候变化和人类活动的影响, 从而影响研究区碳源/汇属性[51]。
黄河流域在2000—2020年累积净碳汇总量为2339.56 MgC(表 2)。其中, 黄土高原农业与草原生态区(Ⅰ—12)位于黄河流域中部, 生态区面积最大, 区内汾渭平原是黄河流域重要的农业区, 光热水分匹配度高, 植被以碳汇作用为主, 是黄河流域净碳汇量最高的生态区;其次为江河源区-甘南高寒草甸草原生态区(Ⅲ—4), 该区位于黄河流域的上游地区, 国内外多条大河的发源地, 植被以高山草原草甸为主, 区内湟水谷地生态环境优越, 农业条件发达, 是世界屋脊上的“隐秘江南”, 耕地面积占青海省的56%, 属于黄河流域强碳汇区, 21年累积净碳汇量超过500 MgC;秦巴山地落叶与常绿阔叶林生态区(Ⅰ—15)及燕山—太行山山地落叶阔叶林生态区(Ⅰ—10)区内山地植被生长茂盛, 累积NEP平均值较高, 但生态区面积相对较小, 总固碳量不突出;西部干旱大区(Ⅱ)位于黄河流域北部地区, 该区植被类型以草原、荒漠为主, 植被覆盖率低, 水土流失严重, 蒙古高原中部-陇中荒漠草原生态区(Ⅱ—2)和内蒙古高原中部草原化荒漠生态区(Ⅱ—3)整体呈碳源区;内蒙古高原中东部典型草原生态区(Ⅱ—1)呈弱碳汇区。总体而言, 黄河流域各生态功能区以碳汇作用为主, 碳汇储量呈现中部高, 南北低的空间分布格局。
| 黄河流域生态功能区 Yellow River Basin Ecological Function Area |
碳汇区Carbon sink area | 碳源区 | 总净碳汇量 Net Carbon Sink/MgC |
|||
| 累积NEP均值 Cumulative NEP mean value /(gC/m2) |
总固碳量 Total carbon sequestration /MgC |
累积NEP均值 Cumulative NEP mean value /(gC/m2) |
总排碳量 Total carbon emissions /MgC |
|||
| Ⅰ—8 | 3782.473 | 37.80 | -1389.22 | -1.88 | 35.93 | |
| Ⅰ—10 | 6330.09 | 210.48 | -1210.22 | -1.84 | 208.63 | |
| Ⅰ—11 | 5217.68 | 257.88 | -1573.7 | -7.78 | 250.1 | |
| Ⅰ—12 | 4074.87 | 847.78 | -565.06 | -12.88 | 834.91 | |
| Ⅰ—13 | 3561.66 | 52.4 | -2378.72 | -9.43 | 42.97 | |
| Ⅰ—15 | 7775.98 | 304.54 | -1606.04 | -2.85 | 301.69 | |
| Ⅱ—1 | 1491.46 | 57.83 | -838.83 | -41.54 | 16.29 | |
| Ⅱ—2 | 1711.81 | 47.01 | -1253.14 | -109.63 | -51.79 | |
| Ⅱ—3 | 816.94 | 0.43 | -1942.47 | -14.56 | -14.13 | |
| Ⅲ—1 | 3678.79 | 119.03 | -680.46 | -0.43 | 118.60 | |
| Ⅲ—4 | 3779 | 605 | -727.26 | -6 | 599 | |
| 黄河流域Yellow River Basin | 4139.77 | 2548.70 | -1101.90 | -209.14 | 2339.56 | |
| Ⅰ—8:辽东-山东丘陵落叶阔叶林生态区、Ⅰ—10:燕山—太行山山地落叶阔叶林生态区、Ⅰ—11:汾渭盆地农业生态区、Ⅰ—12:黄土高原农业与草原生态区、Ⅰ—13:华北平原农业生态区、Ⅰ—15:秦巴山地落叶与常绿阔叶林生态区、Ⅱ—1:内蒙古高原中东部典型草原生态区、Ⅱ—2:蒙古高原中部-陇中荒漠草原生态区、Ⅱ—3:内蒙古高原中部草原化荒漠生态区、Ⅲ—1:祁连山森林与高寒草原生态区、Ⅲ—4:江河源区-甘南高寒草甸草原生态区 | ||||||
碳源/汇的形成是多种气象要素综合作用的结果, 本文探讨影响黄河流域碳源/汇形成的三个主要气象要素(降水、气温、太阳辐射)(图 10)。黄河流域植被碳源/汇与降水整体呈现正相关, 正相关面积占研究区面积的78.08%, 负相关面积占据21.92%。负相关区域主要位于江河源区-甘南高寒草甸草原生态区(Ⅲ—4)、祁连山森林与高寒草原生态区(Ⅲ—1)以及黄土高原农业与草原生态区(Ⅰ—12)、秦巴山地落叶与常绿阔叶林生态区(Ⅰ—15)的南部地区, 显著负相关主要集中在江河源区-甘南高寒草甸草原生态区(Ⅲ—4)的西南部以及祁连山森林与高寒草原生态区(Ⅲ—1)的西北部地区, 该地区平均海拔较高, 地形以高原、山地为主, 多草地, 受降水影响大, 降水的减少, 在一定程度上抑制了土壤微生物呼吸作用, 植被NEP反而提升;甘肃以东的中下游段主要呈正相关, 显著正相关区域主要集中在黄土高原农业与草原生态区(Ⅰ—12)的西部和北部地区、内蒙古高原中部-陇中荒漠草原生态区(Ⅱ—2)的西部、北部地区以及内蒙古高原中东部典型草原生态区(Ⅱ—1)的大部分地区, 该地区是黄河流域主要的碳源区, 碳汇的增长与降水量呈正相关, 主要原因在于该地区多荒漠, 属于干旱半干旱地区, 降水量不足以满足植被生长的需要, 成为限制当地经济和生态发展的主要因素。

|
| 图 10 2000—2020年黄河流域NEP与气象要素相关性 Fig. 10 Correlation between NEP and meteorological elements in the Yellow River Basin from 2000 to 2020 |
黄河流域植被碳源/汇与气温的正负相关面积占比相差不大, 负相关面积占总面积的45.26%, 正相关面积占总面积的54.74%, 相关系数在-0.84—0.83之间浮动。在植被碳源区, 基本上呈现负相关, 在植被碳汇区, 大部分区域为正相关, 显著正相关区域主要位于江河源区-甘南高寒草甸草原生态区(Ⅲ—4)的青海省部分;显著负相关主要位于汾渭盆地农业生态区(Ⅰ—11)的南部秦岭山地。植被碳源/汇与太阳辐射整体呈现正相关, 正相关区域占研究区面积的78.3%, 负相关区域占21.7%, 负相关区域主要分布于黄土高原农业与草原生态区(Ⅰ—12)和内蒙古高原中部-陇中荒漠草原生态区(Ⅱ—2)的西部地区, 即黄河流域的中游区域, 上游和下游则以正相关为主。
总体而言, 降水和太阳辐射对黄河流域的影响最大, 共计影响面积87.56%, 气温的影响面积约占12.44%。黄河流域上游地区的江河源区-甘南高寒草甸草原生态区(Ⅲ—4)、祁连山森林与高寒草原生态区(Ⅲ—1)受太阳辐射和气温的双重控制;中游的北部和东部主要受太阳辐射的控制, 集中于汾渭盆地农业生态区(Ⅰ—11)、内蒙古高原中东部典型草原生态区(Ⅱ—1)、内蒙古高原中部草原化荒漠生态区(Ⅱ—3);中游的中部黄土高原农业与草原生态区(Ⅰ—12)和内蒙古高原中部-陇中荒漠草原生态区(Ⅱ—2)以及下游华北平原农业生态区(Ⅰ—13)主要受降水控制。
分别对降水、气温、太阳辐射进行函数拟合, 得到图 11。可以看出, 当月降水量低于50 mm时, NEP在碳源/汇边界变化, 月降水量达到100—150 mm时NEP值达到最高值, 月降水量高于150 mm时, 随着降水量的增加, NEP呈缓慢下降趋势。同时, 根据统计, 黄河流域在2000—2020年月总降水量低于100 mm的月份占比90%, 植被生长受到降水量的严重制约;随着气温的升高, NEP值呈上升趋势, 在0—20℃范围内NEP变化不大, 基本在弱碳源—弱碳汇范围内浮动, 当升温至30℃左右时, NEP分布范围最广, 且达到最大值, 说明增温在一定程度上促进植被生长发育, 植被固碳能力增强[52];太阳辐射对NEP的影响呈近“正态”分布变化, 当月辐射量在550—600 MJ/m2范围内时, 植被NEP保持高位波动。综上分析, 当气温为30℃, 降水达到150 mm, 太阳辐射约为550 MJ/m2时, 植被固碳能力效果最强。

|
| 图 11 黄河流域NEP与气象要素拟合函数 Fig. 11 Fit function between NEP and meteorological elements in the Yellow River Basin |
(1) 黄河流域植被NEP变化与降水和太阳辐射的相关性最为显著, 气温影响面积最少, 这与Jiang F等[53]、曹明奎等[54]认为中国陆地生态系统碳汇与温度呈强相关的研究结果有所出入。这可能与研究区范围大小不一致有关。黄河流域跨越半湿润、半干旱、干旱多种气候区, 植被生长对水分依赖性极强, 同时区内地形起伏较大, 受降水限制的同时极易受到太阳辐射的影响;黄河流域处于中纬度地带, 南北跨越少, 温差小、气温相对稳定, 植被NEP对气温敏感性相对较低。气候变化对黄河流域植被碳汇的影响是复杂的, 本文仅探究了主要气象要素对植被生长影响, 更需要结合土壤、地形等下垫面要素以及土地利用变化等人类活动进一步开展机理研究。
(2) 模型的验证是估算大尺度区域生物量的关键, 本文借助修正后的CASA模型和土壤微生物呼吸量模型对黄河流域各生态功能区碳汇储量进行了估算, 但仍存在以下不确定性:一是本文采取与其他研究成果对比和与其他遥感数据对比的方式验证NPP和Rh的精度, 未获取实测数据进行验证;二是本文在对NPP和Rh结果验证的基础上间接验证NEP, 未能直接验证NEP的精度;三是本文对CASA模型参数进行了本地化修正, 但未进行长时间尺度的观测, 模型的适用性尚需进行多方验证。
(3) 本文按照黄河流域各生态功能区进行分区研究植被碳源/汇, 有助于更好地体现生态功能区的价值, 有针对性的解决生态区植被碳汇不足的问题。西部干旱大区(Ⅱ)是黄河流域主要的碳源及弱碳汇区域, 该地区植被生长主要受降水的限制, 同时面临严重草原退化问题, 针对这一特点, 可开展春季休牧政策, 同时配合划区轮牧管理措施[55], 最大程度保障天然草场的恢复工作;华北平原农业生态区总体也呈弱碳汇, 该地区人类活动强度大[56], 黄河入海口大量林地和草地转化为水库、盐场等, 导致植被NEP大幅萎缩[57], 因此要控制人类活动对生态环境的干预, 在追求经济价值的同时保护生态环境, 可改良盐碱荒地为农田, 因地制宜的种植耐盐碱植物, 增强区域碳汇能力[58]。
3.2 结论(1) 黄河流域NPP值在时间尺度上整体呈波动上升趋势, 多年平均NPP值为317.18 gC m-2 a-1, 秦巴山地落叶与常绿阔叶林生态区年均NPP值最高, 为576.55 gC m-2 a-1;林地>耕地>沼泽>草地, 水体、戈壁、风蚀劣地和裸沙地的NPP总体处于较低值;夏季>秋季>春季>冬季;空间尺度上, 黄河流域NPP呈现出南高北低的分布特点, 高值区集中在黄土高原农业与草原生态区与汾渭盆地农业生态区交界处, 空间差异显著。
(2) 黄河流域在2000—2020年Rh值变化趋势大致与NPP一致, 极大值出现2018年, 为189.90 gC m-2 a-1, 最小值出现在2004年为172.06 gC m-2 a-1;空间上呈现从东南往西北、从东往西依次递减的空间分布格局, 高值主要分布在汾渭盆地农业生态区, 低值主要位于祁连山森林与高寒草原生态区和江河源区-甘南高寒草甸草原生态区。
(3) 黄河流域总体呈碳汇属性, 碳汇区域由2000年的64.22%增长到2020年的76.31%, 增长趋势以黄土高原农业与草原生态区为中心向北向东北方向扩张。21年间黄河流域植被累积NEP平均值为2857.17 gC m-2 a-1, 年均固碳量约为2301.33 MgC/a, 累积净碳汇总量2339.56 MgC。
(4) 黄河流域植被碳汇的形成与降水和太阳辐射的相关性最大, 共计影响面积87.56%, 且以正相关为主, 气温的影响面积约占12.44%;当气温为30℃, 降水达到150 mm, 太阳辐射约为550 MJ/m2时, 植被固碳能力效果最强。
| [1] |
於琍, 朴世龙. IPCC第五次评估报告对碳循环及其他生物地球化学循环的最新认识. 气候变化研究进展, 2014, 10(1): 33-36. DOI:10.3969/j.issn.1673-1719.2014.01.007 |
| [2] |
赵荣钦, 李志萍, 韩宇平, Milind Kandlikar, 张战平, 丁明磊. 区域"水-土-能-碳"耦合作用机制分析. 地理学报, 2016, 71(9): 1613-1628. |
| [3] |
Piao S, Huang M, Liu Z, Wang X, Ciais P, Canadell J G, Wang K, Bastos A, Friedlingstein P, Houghton R A, Le Quéré C, Liu Y, Myneni R B, Peng S, Pongratz J, Sitch S, Yan T, Wang Y, Zhu Z, Wu D, Wang T. Lower land-use emissions responsible for increased net land carbon sink during the slow warming period. Nature Geoscience, 2018, 11(10): 739-743. DOI:10.1038/s41561-018-0204-7 |
| [4] |
Wofsy S C, Goulden M L, Munger J W, Fan S M, Bakwin P S, Daube B C, Bassow S L, Bazzaz F A. Net exchange of CO2 in a mid-latitude forest. Science, 1993, 260(5112): 1314-1317. DOI:10.1126/science.260.5112.1314 |
| [5] |
赵宁, 周蕾, 庄杰, 王永琳, 周稳, 陈集景, 宋珺, 丁键浠, 迟永刚. 中国陆地生态系统碳源/汇整合分析. 生态学报, 2021, 41(19): 7648-7658. |
| [6] |
王川, 刘永昌, 李稚. 塔里木河下游生态输水对植被碳源/汇空间格局的影响. 干旱区地理, 2021, 44(3): 729-738. |
| [7] |
赖碧海. 江西省近二十年碳源碳汇变化研究[D]. 南昌: 南昌大学, 2021.
|
| [8] |
Woodall C W, Walters B F, Coulston J W, D'Amato A W, Domke G M, Russell M B, Sowers P A. Monitoring Network Confirms Land Use Change is a Substantial Component of the Forest Carbon Sink in the eastern United States. Scientific Reports, 2015, 5: 17028. DOI:10.1038/srep17028 |
| [9] |
周伟, 何顺兵, 杨晗, 何绪刚, 牟凤云. 基于BIOME-BGC模型的三峡库区不同草地群落碳源/汇的动态变化研究. 山地学报, 2018, 36(1): 13-22. |
| [10] |
于伟咏, 漆雁斌, 明辉, 任丹, 邓鑫, 傅丽. 中国农业系统碳汇和碳源的省域差异及影响因素. 西部林业科学, 2016, 45(5): 1-7. |
| [11] |
方精云, 郭兆迪, 朴世龙, 陈安平. 1981-2000年中国陆地植被碳汇的估算. 中国科学: D辑: 地球科学, 2007, 37(6): 804-812. |
| [12] |
王喜, 鲁丰先, 秦耀辰, 孙艳芳. 河南省碳源碳汇的时空变化研究. 地理科学进展, 2016, 35(8): 941-951. |
| [13] |
张璐, 王静, 施润和. 2000-2010年东北三省碳源汇时空动态遥感研究. 华东师范大学学报: 自然科学版, 2015(4): 164-173. |
| [14] |
Cox P M, Betts R A, Jones C D, Spall S A, Totterdell I J. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature, 2000, 408(6809): 184-187. DOI:10.1038/35041539 |
| [15] |
He H L, Wang S Q, Zhang L, Wang J B, Ren X L, Zhou L, Piao S L, Yan H, Ju W M, Gu F X, Yu S Y, Yang Y H, Wang M M, Niu Z E, Ge R, Yan H M, Huang M, Zhou G Y, Bai Y F, Xie Z Q, Tang Z Y, Wu B F, Zhang L M, He N P, Wang Q F, Yu G R. Altered trends in carbon uptake in China's terrestrial ecosystems under the enhanced summer monsoon and warming hiatus. National Science Review, 2019, 6(3): 505-514. |
| [16] |
于贵瑞, 李轩然, 王秋凤, 李胜功. 中国陆地生态系统的碳储量及其空间格局. Journal of Resources and Ecology, 2010, 1(2): 97-109. |
| [17] |
马明娟, 李强, 周文瑞. 碳中和视域下黄河流域碳生态补偿研究. 人民黄河, 2021, 43(12): 5-11. |
| [18] |
贾松伟. 黄河流域森林植被碳储量分布特征及动态变化. 水土保持研究, 2018, 25(5): 78-82, 88. |
| [19] |
颜明, 贺莉, 王随继, 郑明国, 孙莉英, 许炯心. 基于NDVI的1982-2012年黄河流域多时间尺度植被覆盖变化. 中国水土保持科学, 2018, 16(3): 86-94. |
| [20] |
郭睿妍, 田佳, 杨志玲, 杨泽康, 苏文瑞, 刘文娟. 基于GEE平台的黄河流域森林NPP时空变化特征. 生态学报, 2022(13): 1-9. DOI: http://kns.cnki.net/kcms/detail/11.2031.Q.20220316.1720.030.html
|
| [21] |
巩杰, 张影, 钱彩云. 甘肃白龙江流域净生态系统生产力时空变化. 生态学报, 2017, 37(15): 5121-5128. |
| [22] |
孙从建, 乔鹏, 王佳瑞, 王红阳, 孙九林. 2000年来吕梁连片贫困区植被净初级生产力时空变化特征. 生态学报, 2022, 42(1): 277-286. |
| [23] |
朱文泉, 潘耀忠, 张锦水. 中国陆地植被净初级生产力遥感估算. 植物生态学报, 2007, 31(3): 413-424. |
| [24] |
朱文泉, 潘耀忠, 何浩, 于德永, 扈海波. 中国典型植被最大光利用率模拟. 科学通报, 2006, 51(6): 700-706. |
| [25] |
Bond-Lamberty B, Wang C K, Gower S T. A global relationship between the heterotrophic and autotrophic components of soil respiration?. Global Change Biology, 2004, 10(10): 1756-1766. |
| [26] |
Bond-Lamberty B, Thomson A. Temperature-associated increases in the global soil respiration record. Nature, 2010, 464(7288): 579-582. |
| [27] |
Raich J W, Potter C S, Bhagawati D. International variability in global soil respiration, 1980-94. Global Change Biology, 2002, 8(8): 800-812. |
| [28] |
石志华. 基于CASA与GSMSR模型的陕西省植被碳汇时空模拟及影响因素研究[D]. 杨凌: 西北农林科技大学, 2015.
|
| [29] |
裴志永, 周才平, 欧阳华, 杨文斌. 青藏高原高寒草原区域碳估测. 地理研究, 2010, 29(1): 102-110. |
| [30] |
潘竟虎, 文岩. 中国西北干旱区植被碳汇估算及其时空格局. 生态学报, 2015, 35(23): 7718-7728. |
| [31] |
刘春雨. 省域生态系统碳源/汇的时空演变及驱动机制——以甘肃省为例[D]. 兰州: 兰州大学, 2015.
|
| [32] |
刘凤, 曾永年. 2000-2015年青海高原植被碳源/汇时空格局及变化. 生态学报, 2021, 41(14): 5792-5803. |
| [33] |
范也平. 河西走廊植被碳源/汇时空变化及其对气候变化的响应[D]. 兰州: 西北师范大学, 2018.
|
| [34] |
贠银娟, 赵军. 基于MODIS-NDVI数据的植被碳汇空间格局研究——以石羊河流域为例. 山地学报, 2018, 36(4): 644-653. |
| [35] |
王强, 张廷斌, 易桂花, 陈田田, 别小娟, 何奕萱. 横断山区2004-2014年植被NPP时空变化及其驱动因子. 生态学报, 2017, 37(9): 3084-3095. |
| [36] |
曹永强, 王菲. 河北省植被时空变化及其对气象干旱的响应. 南水北调与水利科技: 中英文, 2021, 19(4): 708-719, 757. |
| [37] |
朱玉果, 杜灵通, 谢应忠, 刘可, 宫菲, 丹杨, 王乐. 不同气象插值方法精度评估及对草地NPP估算的影响. 水土保持研究, 2018, 25(6): 160-167. |
| [38] |
王娟, 何慧娟, 董金芳, 郭斌. 黄河流域植被净初级生产力时空特征及自然驱动因子. 中国沙漠, 2021, 41(6): 213-222. |
| [39] |
张丹丹. 基于多源数据的黄河流域植被NPP时空变化及其影响因子分析[D]. 郑州: 郑州大学, 2019.
|
| [40] |
朱莹莹. 1992-2015年黄河流域植被净初级生产力遥感估算及其对气候变化的响应[D]. 西安: 长安大学, 2019.
|
| [41] |
贠银绢. 2000-2015年石羊河流域植被碳汇时空变化及影响因子研究[D]. 兰州: 西北师范大学, 2018.
|
| [42] |
田琴, 牛春梅, 谷口武士, 山中典和, 时伟宇, 杜盛. 黄土丘陵区植被类型与土壤微生物区系及生物量的关系. 生态学报, 2017, 37(20): 6847-6854. |
| [43] |
赵冰清. 半干旱黄土区大型露天煤矿植被演替规律研究[D]. 北京: 中国地质大学(北京), 2019.
|
| [44] |
刘建涛. 黄河三角洲典型地表类型遥感协同提取方法及生态环境遥感评价研究[D]. 北京: 中国科学院大学(中国科学院遥感与数字地球研究所), 2018.
|
| [45] |
袁丽丽, 杨玉萍. 贺兰山自然保护区生态环境保护成效及存在问题与对策建议. 内蒙古林业, 2020(11): 17-21. |
| [46] |
周侃, 樊杰, 王亚飞, 李九一. 干旱半干旱区水资源承载力评价及空间规划指引——以宁夏西海固地区为例. 地理科学, 2019, 39(2): 232-241. |
| [47] |
李婷婷. 宁夏生态移民居住文化变迁研究——以西海固生态移民为例[D]. 临汾: 山西师范大学, 2018.
|
| [48] |
白壮壮, 崔建新, 丁晓辉. 1986-2015年鄂尔多斯高原沙漠化及其驱动因素研究. 干旱区研究, 2020, 37(3): 749-756. |
| [49] |
何航. 黄河流域上中游植被覆盖变化及驱动因素研究[D]. 兰州: 西北师范大学, 2020.
|
| [50] |
杨洁, 谢保鹏, 张德罡. 基于InVEST和CA-Markov模型的黄河流域碳储量时空变化研究. 中国生态农业学报: 中英文, 2021, 29(6): 1018-1029. |
| [51] |
杨季. 试论草原碳源与碳汇的对立统一关系及草原碳汇的作用. 国家林业和草原局管理干部学院学报, 2019, 18(2): 8-12. |
| [52] |
Cao M, Woodward F I. Dynamic responses of terrestrial ecosystem carbon cycling to global climate change. Nature, 1998, 393(6682): 249-252. |
| [53] |
Fang J Y, Yu G R, Liu L L, Hu S J, Chapin F S Ⅲ. Climate change, human impacts, and carbon sequestration in China. PNAS, 2018, 115(16): 4015-4020. |
| [54] |
曹明奎, 陶波, 李克让, 邵雪梅, Stephen DPRIENCE. 1981-2000年中国陆地生态系统碳通量的年际变化. Acta Botanica Sinica, 2003, 45(5): 552-560. |
| [55] |
张全民. 锡林郭勒草原固碳潜力估算与碳增汇功能区划分[D]. 呼和浩特: 内蒙古大学, 2019.
|
| [56] |
张心茹, 曹茜, 季舒平, 陈浩, 张廷靖, 刘建. 气候变化和人类活动对黄河三角洲植被动态变化的影响. 环境科学学报, 2022, 42(1): 56-69. |
| [57] |
池源, 石洪华, 孙景宽, 李捷, 杨帆, 付战勇. 近30年来黄河三角洲植被净初级生产力时空特征及主要影响因素. 生态学报, 2018, 38(8): 2683-2697. |
| [58] |
董红云, 朱振林, 李新华, 杨丽萍, 张正. 山东省盐碱地分布、改良利用现状与治理成效潜力分析. 山东农业科学, 2017, 49(5): 134-139. |