 2023, Vol. 43
2023, Vol. 43文章信息
- 张馨文, 安慧, 杜忠毓, 刘姝萱, 薛斌, 刘小平, 文志林
- ZHANG Xinwen, AN Hui, DU Zhongyu, LIU Shuxuan, XUE Bin, LIU Xiaoping, WEN Zhilin
- 氮添加对宁夏荒漠草原植物初级生产力的影响机制
- Effects and mechanism of nitrogen addition on primary productivity in a desert grassland of Ningxia, China
- 生态学报. 2023, 43(6): 2476-2487
- Acta Ecologica Sinica. 2023, 43(6): 2476-2487
- http://dx.doi.org/10.5846/stxb202203100573
-
文章历史
- 收稿日期: 2022-03-10
- 网络出版日期: 2022-11-04





2. 盐池县草原实验站, 盐池 751506
2. Grassland Experiment Station of Yanchi, Yanchi 751506, China
全球变化对生态系统功能和服务以及人类福祉有至关重要的影响。而全球变化导致生物多样性的减少可能会危害生态系统, 因此了解生物多样性与生态系统功能之间的关系成为生态学家研究的热点[1]。大气中N沉降已成为国际上备受关注的全球变化问题之一, N沉降能通过改变植物物种代谢和生理机能直接影响生态系统功能, 也可以通过改变生物多样性间接影响生态系统功能[2]。N沉降主要通过提高土壤养分含量从而促进了内蒙古半干旱典型草原初级生产力[3];但关于N沉降如何通过改变生物多样性进而间接影响生态系统功能的研究较少。物种多样性和功能多样性是生物多样性的重要组成部分。功能多样性是指影响生态系统功能过程的种间功能特征变化的范围, 可以由多个指数反映, 包括Rao指数、功能离散度、功能均匀度及群落性状加权平均值等[4]。与物种多样性相比, 功能多样性考虑了植物群落内的冗余性和种间互补机制, 并把植物功能性状与生态系统功能相联结[5], 可用多个植物功能性状来解释生态系统功能[6]。N添加降低了祁连山亚高山草地群落物种多样性和典型草原功能多样性[7—8];对内蒙古极度退化草地恢复进程中物种多样性无显著影响[9]。但关于物种多样性与功能多样性之间呈何种关系仍存在较大的争议。已有研究表明, 物种多样性与功能多样性的关系可呈正相关、负相关、不相关或呈S形曲线[10—14]。因此, 物种多样性与功能多样性之间的关系随着扰动强度及环境胁迫而发生改变。
生物多样性与初级生产力的关系存在两种解释机制, 即“生态位互补假说”[15]和“生物量比率假说”[16]。生态位互补一般泛指物种之间的自然资源综合利用效率之间存在着较大的差异, 当该假说占优势时, 初级生产力变化用多样性指数来表征。生物量比率假说指出生态系统功能对物种丰富度的变化相对不敏感, 但在很大程度上取决于优势种, 通常用群落性状加权平均值表征该假说。生物量比率假说能更好的解释位于我国东部的亚热带次生林和黄土丘陵退耕还草区初级生产力变异[17—18];而生态位互补假说在解释阔叶红松林群落内生产力的变异中占主导地位[19]。Cardinale等[20]通过整合分析研究了生物多样性与初级生产力的关系, 结果表明生物量比率假说和生态位互补假说对初级生产力变化的解释一致。青藏高原高寒草甸植物群落物种丰富度与地上生产力之间呈负相关关系[21]。功能多样性通过群落中植物子叶类型、植株高度等性状的多样化程度的增加, 从而增加了植物物种从不同的时空尺度、用不同的方式对有限资源进行最大程度的互补性利用率, 减少资源浪费, 进而对植物初级生产力产生影响, 使生态系统功能得以高效的发挥[22]。因此, 研究植物群落物种多样性、功能多样性与初级生产力的关系, 对于揭示物种多样性、功能多样性变化对生态系统功能的潜在影响具有重要意义。
荒漠草原作为中国西北生态屏障的重要组成部分, 是草原向荒漠过渡的旱生化草地生态系统, 是干旱、半干旱地区生态系统的主要组成部分。过量的N沉降不仅影响陆地生态系统碳氮循环, 也对生态系统的物种组成、生物多样性及初级生产力产生影响[23]。同时过量的N沉降导致荒漠草原退化、物种丰富度减少及生态系统服务功能下降[24]。近年来, 关于N添加对荒漠草原的影响主要集中在物种多样性、初级生产力及物种组成的影响等方面[25—26]。认为适量N添加改变了物种组成和物种多样性, 从而增加荒漠草原植物初级生产力。然而, 目前关于N添加对荒漠草原植物功能多样性的影响及其与初级生产力的关系的研究较少, 植物功能多样性对氮素添加的响应机制有待进一步研究。因此, 本研究以宁夏盐池县荒漠草原为研究对象, 主要研究荒漠草原物种多样性、功能多样性及初级生产力对不同N添加量的响应, 探讨N添加下植物多样性对荒漠草原初级生产力的维持机制。为半干旱地区退化荒漠草原恢复重建以及退耕还草提供理论依据和实践指导。
1 材料与方法 1.1 研究区概况本研究在宁夏荒漠草原生态系统定位观测研究站进行(37°31′ N, 106°93′ E, 海拔1523 m)。定位观测研究站位于宁夏回族自治区盐池县, 该地区气候类型为典型的中温带大陆性气候, 年均降水量约300 mm, 降水多集中于7—9月份, 占全年降水的70%;年潜在蒸发量为2384 mm;年平均气温7.6℃, 其中1月平均气温-8.7℃, 7月平均气温22.4℃。地带性土壤主要是灰钙土, 非地带性土壤主要是风沙土和草甸土等。土壤结构较为松散, 养分匮乏。土壤pH为8.4, 弱碱性。土壤有机碳含量为6.17 g/kg, 全氮含量为0.63 g/kg, 全磷含量为0.30 g/kg, 全钾含量为14.35 g/kg, 碱解氮含量为36.14 mg/kg, 速效磷含量为4.25 mg/kg, 速效钾含量为174.10 mg/kg。植被类型主要有草原、沙地植被以及荒漠化草原植被, 主要物种有草原石头花(Gypsophila davurica)、长芒草(Stipa bungeana)、达乌里胡枝子(Lespedeza daurica)、糙隐子草(Cleistogenes squarrosa)、短翼岩黄耆(Hedysarum brachypterum)等。该地区干旱少雨且基质较差, 植物生长矮小, 群落层片结构多呈单层[27—28]。
1.2 实验设计2017年8月选择地势平坦的荒漠草原进行围封并建立N添加试验样地, 自2018年5月开始进行N添加处理。N肥施用量参考荒漠草原N添加实验[29]和区域大气N沉降水平[30]。实验处理包括对照和4个N添加水平(5、10、20和40 g m-2 a-1), 共5个N处理:CK、N5、N10、N20和N40。每一处理4个重复, 共计20个小区。实验采用完全随机区组设计, 每个小区面积为6 m×6 m, 小区间均设置有2 m宽的缓冲带。N肥为缓释型尿素[(NH2)2CO](含纯N量为46.4%), 为了保证N是唯一的限制性元素, 除了对照之外, 其他N添加处理都添加1.5 g m-2 a-1磷肥, 磷肥为过磷酸钙(Ca(H2PO4)2)[31]。每年5月初将缓释型尿素和磷肥于降雨前撒施。
1.3 植物群落调查与采样2018年至2021年每年8月中旬, 在每个小区选取1 m×1 m的固定样方进行植物群落调查。调查样方内植物的物种数并且分物种测定株高和密度(株丛数), 之后设置2个0.2 m×1 m的小样方用于测定植物地上生物量。将小样方内植物按照物种分类, 齐地面剪下其地上部分, 带回实验室烘干至恒重(65℃, 48 h), 按照物种进行称重测定地上生物量(精度为0.01 g)。样方内所有物种的地上生物量之和即为群落初级生产力(PP, g m-2 a-1)。
1.4 生物多样性计算采用R 4.0.2 vegan包的diversity函数和FDiversity包分别计算物种多样性及功能多样性(Rao指数、功能均匀度、功能离散度和群落性状加权平均值)。
1.4.1 物种多样性物种多样性用Shannon-Wiener指数(H′)和物种丰富度(S)表征:
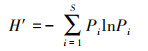
|
式中, P为物种个体数占样方中所有物种个体的比率, P=Ni/N, Ni为物种i的个体数, N为样方中所有物种个体数之和;S为样方内的物种数。
1.4.2 功能多样性本研究筛选与初级生产力显著相关的6个功能性状, 包括2个定量功能性状(植物地上生物量、植株高度)和4个定性功能性状(生活周期、生长类型、子叶类型和植物类群)来计算功能多样性[32], 定性功能性状赋值详见表 1。用实测值标准化的方法来计算定量性状[33]。本文计算了多性状(6个性状)的功能多样性值。
| 定性性状 Qualitative functional traits |
赋值 Assignment |
| 生活周期 Life cycle |
0=非多年生 1=多年生 |
| 生长类型 Growth form |
1=散生 2=丛生 3=密丛生 |
| 子叶类型 Cotyledon type |
0=单子叶 1=双子叶 |
| 植物类群 Plant groups |
1=禾草科 2=豆科 3=杂类草 |
使用SPSS 22.0—单因素方差和Duncan′s多重比较, 分析物种多样性、功能多样性和初级生产力对N添加的响应。对初级生产力数据进行对数转换, 并通过SPSS 20.0—Correlate和Regression分析功能多样性、物种多样性与初级生产力三者之间的相关性。以上统计检验的显著水平为P < 0.05。使用SigmaPlot 14.0软件作图。
结构方程模型利用R 4.1.2 lavaan包完成。本研究结构方程模型的构建选取与初级生产力相关性较高的变量, 包括物种丰富度[38]、功能离散度[39]及群落加权平均值-株高[40]。利用卡方检验的p值、近似误差平方根(RMSEA)、标准化残差均方根(SRMR)、相对匹配度(CFI)、拟合优势度(GFI)评价模型的优度[41]。拟合优度的临界值为p>0.05, RMSEA < 0.06, SRMS < 0.09, CFI>0.95, GFI>0.90。
2 结果与分析 2.1 N添加对荒漠草原物种多样性和功能多样性的影响实验处理第一年(2018年), N添加对荒漠草原物种多样性(Shannon-Wiener指数、物种丰富度)没有显著的影响;随着处理年限的增加, N添加对荒漠草原物种多样性产生显著影响(图 1)。2019年, 荒漠草原植物Shannon-Wiener指数和物种丰富度随N添加量增加呈先增加后降低的趋势, N10和N20处理的Shannon-Wiener指数和物种丰富度均显著高于对照(P < 0.05)。2020年和2021年荒漠草原植物Shannon-Wiener指数与物种丰富度随着N添加量增加而呈下降趋势, 且高N添加量(N20和N40)处理的Shannon-Wiener指数与物种丰富度显著低于对照(P < 0.05)。其中2021年高N添加量(N20和N40)处理下Shannon-Wiener指数和物种丰富度分别比对照减少了80.5%、71.0%和79.4%、61.29%。

|
| 图 1 N添加对荒漠草原Shannon-Wiener指数和物种丰富度的影响 Fig. 1 Effects of N addition on species richness and Shannon-Wiener index in desert grassland 不同字母表示差异显著(P < 0.05) |
不同实验年份, 各N添加量对荒漠草原功能多样性的影响不同。第一年和第二年(2018年、2019年), N添加对功能多样性(Rao指数)、功能均匀度及群落加权平均值-株高影响不显著(图 2);而第三年和第四年(2020年、2021年)N添加对功能多样性(Rao指数)、功能均匀度及群落加权平均值-株高的影响显著。2018年N添加对功能离散度无显著影响, 而后三年N10、N40处理对功能离散度的影响显著(P < 0.05)。随着实验年限的增加, 2020年和2021年N添加显著提高了荒漠草原功能多样性(Rao指数)、功能离散度和群落加权平均值-株高(图 2, P < 0.05);而功能均匀度随着N添加量的增加呈先增加后降低的趋势, 在N10时达到最大值(0.74、0.79)。N添加显著影响了群落加权平均值-地上生物量。2020年和2021年高N处理(N20、N40)群落加权平均值-地上生物量显著高于对照(图 2), 而2019年高N处理地上群落加权平均值-地上生物量与对照差异不显著。

|
| 图 2 N添加对荒漠草原功能多样性及初级生产力的影响 Fig. 2 Effects of N addition on functional diversity and primary productivity in desert grassland |
N添加显著影响荒漠草原植物初级生产力(图 2, P < 0.05)。2018年N添加处理的荒漠草原植物初级生产力均显著高于对照。2019年至2021年, 低N处理(N5)的荒漠草原植物初级生产力与对照差异不显著, 而高N添加量(N20、N40)处理的荒漠草原植物初级生产力显著高于对照。N10处理时, 2019年初级生产力没有显著增加, 而2020年和2021年初级生产力显著增加。
2.3 物种多样性、功能多样性及初级生产力的关系由于Shannon-Wiener指数和物种丰富度在N添加处理间的变化一致, 因此仅以Shannon-Wiener指数说明物种多样性与功能多样性的关系。功能多样性(Rao指数)与功能离散度、群落加权平均值-株高呈显著正相关;功能离散度与群落加权平均值-株高也呈显著正相关(表 2)。物种多样性与功能多样性(Rao指数)、功能离散度呈极显著负相关关系(图 3, P < 0.01), 与群落加权平均值-株高呈显著正相关(P < 0.05)。不同N添加量的物种多样性与功能多样性(Rao指数)、功能离散度呈显著负相关, 而物种多样性与功能均匀度、群落加权平均值-株高不相关(表 3)。荒漠草原初级生产力与群落加权平均值-株高呈显著正相关性(图 4, P < 0.05), 而与功能多样性(Rao指数)、功能离散度呈显著负相关性(P < 0.05), 与物种丰富度、Shannon-Wiener指数及功能均匀度不相关(图 4)。
| 名称 Name |
R代码 Argument |
公式 Formula |
说明 Note |
| 功能多样性(FD)[34] Rao′s quadratic entropy |
dbFD (X, comm)$RaoQ | 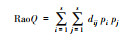 |
S为永久样方内的物种数;pi、pj分别为第i、j个物种的相对密度。dij为欧氏距离, 表示物种i、j在一组性状空间中的相异性。 |
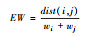 |
|||
| 功能均匀度[35] Functional evenness |
dbFD (X, comm) ¥FEve | 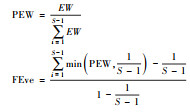 |
EW为加权均匀度;dist(i, j)是物种i和物种j的欧氏距离;PEW为偏加权均匀度;S是物种数。 |
| 功能离散度[36] Functional dispersion |
dbFD (X, comm)$FDis | 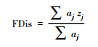 |
aj为物种j的多度, zj为物种j到加权质心的距离。 |
| 群落性状加权平均值[37] community weighted mean |
functcomp (X, comm)$CWM | 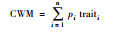 |
pi是物种i的多度, traiti为物种i的性状值, n为物种个数。 |
| X为性状数据矩阵, 6列×16行, 6列为6个性状, 16行是16个物种的性状矩阵;comm为多度数据矩阵, 16列×20行, 16列是16个物种的多度矩阵, 20行为20个样方 | |||

|
| 图 3 物种多样性与功能多样性的关系 Fig. 3 Relationships between species diversity and functional diversity |
| RaoQ | FEve | FDis | CWM-height | |
| RaoQ | 0.147 | 0.587** | 0.946** | |
| FEve | 0.242 | 0.143 | ||
| FDis | 0.510** | |||
| RaoQ、FEve、FDis和CWM-height分别代表功能多样性、功能均匀度、功能离散度及群落加权平均值-株高;**, < 0.05 | ||||

|
| 图 4 物种多样性、功能多样性与植物初级生产力(对数尺度) 的关系 Fig. 4 Relationships between species diversity, functional diversity and primary productivity (logarithmic scale) |
| 氮添加处理 N addition |
RaoQ-H′ | FEve-H′ | FDis-H′ | CWM-height-H′ | |||||||
| r | P | r | P | r | P | r | P | ||||
| CK | -0.341 | >0.05 | 0.007 | >0.05 | -0.262 | >0.05 | -0.087 | >0.05 | |||
| N5 | -0.608 | < 0.05 | -0.296 | >0.05 | -0.571 | < 0.05 | 0.463 | >0.05 | |||
| N10 | -0.700 | < 0.01 | -0.463 | >0.05 | -0.648 | < 0.05 | 0.486 | >0.05 | |||
| N20 | -0.835 | < 0.01 | 0.265 | >0.05 | -0.803 | < 0.01 | 0.375 | >0.05 | |||
| N40 | -0.829 | < 0.01 | 0.121 | >0.05 | -0.779 | < 0.01 | 0.311 | >0.05 | |||
| H′、RaoQ、FEve、FDis和CWM-height分别代表Shannon-Wiener指数、功能多样性、功能均匀度、功能离散度及群落加权平均值-株高 | |||||||||||
结构方程的分析结果发现N添加对荒漠草原群落初级生产力直接影响不显著(图 5)。各变量解释初级生产力变异的45.8%, 群落加权平均值-株高对初级生产力有显著的正效应, 路径系数为0.63;功能离散度和物种丰富度对初级生产力有显著的负效应, 路径系数分别为-0.29、-0.38。

|
| 图 5 N添加对植物初级生产力影响途径的结构方程模型 Fig. 5 Structural equation model analyses of pathways of N addition on productivity Chisq=0.169, df=1, pvalue=0.681, GFI=0.999, CFI=1.000, RMR=0.137, SRMR=0.011, RMSEA=0.000;实线表示变量之间作用关系显著(P < 0.05), 虚线表示变量之间作用关系不显著(P>0.05)。正负数值表示路径系数;**, < 0.05;***, < 0.01 |
物种多样性和功能多样性已经成为生态系统初级生产力的主要驱动力, 是评价生态系统稳定性的重要指标。连续四年N添加显著降低了物种丰富度和Shannon-Wiener指数, 与在典型草原的研究结果较为相似[8]。随着N添加量的增加, 植物对N的利用效率不同, 将引起生态位分化以及增强植物种间对资源的竞争, 导致对光资源和磷等养分竞争能力差的物种被排斥, 以致物种多样性降低[42]。此外, 长期或高量N添加致使土壤酸化, 从而排斥了部分耐酸能力弱的物种, 也是物种多样性降低的原因之一[43]。植物功能均匀度越大, 群落内物种功能特征分布越均匀, 物种对有效资源的利用效率越高;功能离散度越大, 生态位重叠的效应越弱, 生态系统中的资源竞争越弱, 由于资源利用效率较高, 反过来可以增加生态系统的功能[44]。许多研究表明, 施N后, 物种的减少会降低植物的功能多样性[44—45]。但本研究结果表明N添加第三年和第四年功能多样性(Rao指数)、功能离散度及群落加权平均值-株高均显著增加。N添加导致功能多样性的变化与两个相反的机制有关[46]:一是N添加使物种间竞争加剧, 排斥作用会引起功能多样性减少[47];二是共存物种间的生态位分化促使性状趋异, 从而引起功能多样性增加[48]。对照和N添加处理下群落内共有物种较多, 共存物种生态位分化概率较大。此外, N添加改变了不同植物物种对光照、养分等资源的竞争, 同时增加了植物盖度, 减缓了土壤表面的热辐射, 地表水分的蒸发量有所减少, 导致物种间生态位发生分化、功能多样性较高。N添加显著增加了群落加权平均值-株高和群落加权平均值-地上生物量, 这与在典型草原的研究结果相似[49]。N添加缓解了土壤中的养分限制, 促进植物地上部分的生长, 从而增加了植物地上生物量。
功能多样性与物种多样性的关系是研究生物多样性与生态系统功能的关键, 两者关系有助于揭示植物多样性作用机制及确定生态系统的管理政策[50]。本研究结果表明功能多样性与物种多样性呈负相关或不相关, 这与de Bello等[51]和臧岳铭等[10]的研究结果相似。功能多样性与物种多样性的关系既是性状依赖的, 又与环境因素及人为干扰有关, 这与Mayfield等[52]关于两者之间关系是性状依赖的观点一致, 说明二者共变的趋势不是必然的, 即物种多样性与功能多样性不一定等同, 二者之间呈何种关系是有条件的。干旱、养分匮乏等环境选择压力增强时, 群落内组成物种的性状将被限制在适应该选择压力的范围内[2], 物种多样性的降低阻止了生态位分化, 增加了种间性状差异, 功能多样性没有降低甚至增加。功能多样性对物种多样性的依赖程度与物种性状库的幅度及物种在可用生态位的空间分布有关[51]。功能多样性与物种多样性之间的关系会因性状、人为干扰和资源条件的不同而变化。因此, 功能多样性是不等同于物种多样性的一个生物多样性组分, 基于群落水平的物种多样性与基于物种性状的功能多样性之间可能存在互补性, 而不是绝对的替代关系[53]。
3.2 N添加对荒漠草原初级生产力的影响大小及机制初级生产力是陆地生态系统结构和功能的综合体现。本研究结果显示, N添加显著促进了荒漠草原初级生产力, 这一结果与武倩[54]和景明慧[55]的研究较为一致。植物组织中结构蛋白和酶蛋白的合成离不开N素, 在受N限制的草地生态系统中N添加可以促进植物CO2同化作用、植物组织形成和叶片生长[56], 从而提高初级生产力。本研究中, N处理第三年和第四年(2020年和2021年)初级生产力明显低于前两年(2018年和2019年), 这可能是由于受年际间降水量的影响, 2020和2021年降水量(160.8 mm—232.3 mm)低于2018年和2019年的降水量(321.9 mm—364.5 mm)。近年来, 许多研究试图阐明物种多样性、功能多样性对初级生产力的效应。物种多样性、功能多样性和初级生产力之间可表现出正相关、负相关或无相性, 这些相互矛盾的结果反映出生物多样性对初级生产力效应的复杂性[53, 57]。本研究探讨N添加对荒漠草原功能多样性、物种多样性与初级生产力之间关系的影响, 结果表明初级生产力与功能多样性指数(Rao指数、功能离散度)呈极显著负相关关系, 与群落加权平均值-株高呈极显著正相关关系, 而与物种多样性无相关性。与物种多样性相比, 高寒草甸植物功能多样性与初级生产力的关系更为密切[58], 这与本研究结果相同。物种多样性所包含的关于物种的特征信息缺乏或者忽略了物种间功能性状的相似性和差异性。不同物种具有不同的功能性状, 物种多样性不能解释物种间不同的功能特性, 而功能多样性可以综合不同物种的特性;其次, 功能性状可以表征植物从环境中利用资源的效率。功能多样性比物种多样性更好地反映不同物种的生态位, 从而能够更有效地揭示初级生产力的变化[59]。因此, 探讨初级生产力和生物多样性关系时, 应同时考虑物种多样性和功能多样性, 两者对初级生产力的影响可能不同。N添加改变某些植物的生长反应, 调整群落结构及物种组成, 使得群落多样性格局及物种多样性、功能多样性-初级生产力关系发生变化, 最终对整个生态系统的结构和功能产生影响。
本研究发现表征功能多样性的两个指数(功能多样性指数、群落加权平均值-株高)对荒漠草原植物初级生产力的影响相反, 这与Xu等研究结果相似[56]。N添加对初级生产力有间接的积极的影响, 主要是在N匮乏的环境下, N添加提高了物种的整体适应性, 植物高度也会随之发生变化, 植物高度的增加会提高初级生产力, 因此初级生产力对高度的加权平均值更加敏感。此外本研究中, 群落加权平均值-株高、功能离散度及物种丰富度均与初级生产力显著相关, 群落加权平均值-株高的直接效应(0.63)高于功能离散度和物种丰富度的直接效应(0.29、0.38)。因此荒漠草原初级生产力主要是由群落内优势物种的功能性状决定的, 此时生物量比率假说更为重要, 在有生境过滤和扩散限制的群落中, 优势物种的影响可能更大, 生态系统过程主要取决于群落中优势物种的功能性状[60]。我们的研究结果与Xu等在典型草原研究结果相似[56], 而与王玉冰等在典型草原略有不同[8], 其原因可能与选择的功能多样性指数不同有关, Xu等选用功能离散度, 王玉冰采用功能性状多样性指数(FAD指数), 功能性状多样性指数、功能离散度等功能多样性指数表示的意义不同, 不同功能多样性指数对结果产生的影响不同。此外Xu等是选取自样地建立以来的10年数据, 而王玉冰等选取N添加后第18年的数据。本研究采用的是自样地建立以来的4年数据。因此, 多样性影响初级生产力的机制(生物量比率假说、生态位互补假说)可能与功能多样性指数和实验数据的选取有关。
4 结论N添加处理改变了荒漠草原植物物种多样性和功能多样性, 但不同年份的物种多样性和功能多样性对N添加的响应趋势不同。中高N添加显著增加了荒漠草原植物群落初级生产力, 但N添加对荒漠草原群落生产力没有直接影响。N添加主要通过改变物种多样性指数和群落加权平均值-株高进而导致初级生产力变异, 且物种多样性指数和群落加权平均值-株高对初级生产力的作用方向不同, 具体表现为物种多样性指数对初级生产力的影响是负效应, 而群落加权平均值-株高对初级生产力的影响是正效应。
由群落加权性平均值-株高计算的功能多样性, 能更好地预测荒漠草原初级生产力的变化, 表明生物量比率假说是N添加下荒漠草原初级生产力的重要维持机制, 强调了优势物种的性状对N沉降影响生态系统功能的重要性。
| [1] |
Mensah S, Veldtman R, Assogbadjo A E, Kakaï R G, Seifert T. Tree species diversity promotes aboveground carbon storage through functional diversity and functional dominance. Ecology and Evolution, 2016, 6(20): 7546-7557. DOI:10.1002/ece3.2525 |
| [2] |
Díaz S, Lavorel S, De Bello F, Quétier F, Grigulis K, Robson T M. Incorporating plant functional diversity effects in ecosystem service assessments. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(52): 20684-20689. DOI:10.1073/pnas.0704716104 |
| [3] |
嘎玛达尔基, 杨泽, 谭星儒, 王珊珊, 李伟晶, 游翠海, 王彦兵, 张兵伟, 任婷婷, 陈世苹. 凋落物输入变化和氮添加对半干旱草原群落生产力及功能群组成的影响. 植物生态学报, 2020, 44(8): 791-806. |
| [4] |
黄小荣. 广西马尾松林植物功能多样性与生产力的关系. 生物多样性, 2018, 26(7): 690-700. |
| [5] |
Petchey O L, Gaston K J. Functional diversity: back to basics and looking forward. Ecology Letters, 2006, 9(6): 741-758. DOI:10.1111/j.1461-0248.2006.00924.x |
| [6] |
Schleuter D, Daufresne M, Massol F, Argillier C. A user's guide to functional diversity indices. Ecological Monographs, 2010, 80(3): 469-484. DOI:10.1890/08-2225.1 |
| [7] |
曹丰丰, 刘瑞雪, 黄国柱, 吴泓瑾, 赵传燕, 李伟斌. 短期氮添加对祁连山亚高山草地生产力及植物多样性的影响. 生态学报, 2021, 41(12): 5034-5044. |
| [8] |
王玉冰, 孙毅寒, 丁威, 张恩涛, 李文怀, 迟永刚, 郑淑霞. 长期氮添加对典型草原植物多样性与初级生产力的影响及途径. 植物生态学报, 2020, 44(1): 22-32. |
| [9] |
杨倩, 王娓, 曾辉. 氮添加对内蒙古退化草地植物群落多样性和生物量的影响. 植物生态学报, 2018, 42(4): 430-441. |
| [10] |
臧岳铭, 朱志红, 李英年, 王文娟, 席博. 高寒矮嵩草草甸物种多样性与功能多样性对初级生产力的影响. 生态学杂志, 2009, 28(6): 999-1005. |
| [11] |
Sasaki T, Okubo S, Okayasu T, Jamsran U, Ohkuro T, Takeuchi K. Two-phase functional redundancy in plant communities along a grazing gradient in Mongolian rangelands. Ecology, 2009, 90(9): 2598-2608. DOI:10.1890/08-1850.1 |
| [12] |
陈超, 朱志红, 李英年, 姚天华, 潘石玉, 卫欣华, 孔彬彬, 杜家丽. 高寒草甸种间性状差异和物种均匀度对物种多样性与功能多样性关系的影响. 生态学报, 2016, 36(3): 661-674. |
| [13] |
董世魁, 汤琳, 张相锋, 刘世梁, 刘全儒, 苏旭坤, 张勇, 武晓宇, 赵珍珍, 李钰, 沙威. 高寒草地植物物种多样性与功能多样性的关系. 生态学报, 2017, 37(5): 1472-1483. |
| [14] |
刘旻霞, 南笑宁, 张国娟, 李博文, 徐璐, 穆若兰, 李亮, 于瑞新. 高寒草甸不同坡向植物群落物种多样性与功能多样性的关系. 生态学报, 2021, 41(13): 5398-5407. |
| [15] |
Tilman D. Distinguishing between the effects of species diversity and species composition. Oikos, 1997, 80(1): 185-185. DOI:10.2307/3546532 |
| [16] |
Grime J P. Benefits of plant diversity to ecosystems: immediate, filter and founder effects. Journal of Ecology, 1998, 86(6): 902-910. DOI:10.1046/j.1365-2745.1998.00306.x |
| [17] |
Ali A, Yan E R, Chang S X, Cheng J Y, Liu X Y. Community-weighted mean of leaf traits and divergence of wood traits predict aboveground biomass in secondary subtropical forests. Science of the Total Environment, 2017, 574: 654-662. DOI:10.1016/j.scitotenv.2016.09.022 |
| [18] |
唐学娟, 赵峰侠, 张瑞, 闫丽飞, 冀雯霞. 晋西北黄土丘陵退耕还草区优势种功能性状对群落生产力的影响. 干旱区资源与环境, 2020, 34(9): 164-170. |
| [19] |
温纯, 金光泽. 功能多样性对典型阔叶红松林生产力的影响. 植物生态学报, 2019, 43(2): 94-106. |
| [20] |
Cardinale B J, Matulich K L, Hooper D U, Byrnes J E, Duffy E, Gamfeldt L, Balvanera P, O'Connor M I, Gonzalez A. The functional role of producer diversity in ecosystems. American Journal of Botany, 2021, 98(3): 572-592. |
| [21] |
张杰琦, 李奇, 任正炜, 杨雪, 王刚. 氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响. 植物生态学报, 2010, 34(10): 1125-1131. DOI:10.3773/j.issn.1005-264x.2010.10.001 |
| [22] |
江小雷, 张卫国. 功能多样性及其研究方法. 生态学报, 2010, 30(10): 2766-2773. |
| [23] |
Xu Z W, Wan S Q, Ren H Y, Han X G, Li M H, Cheng W X, Jiang Y. Effects of water and nitrogen addition on species turnover in temperate grasslands in northern China. PLoS One, 2012, 7(6): e39762. DOI:10.1371/journal.pone.0039762 |
| [24] |
白永飞, 黄建辉, 郑淑霞, 潘庆民, 张丽霞, 周华坤, 徐海量, 李玉霖, 马健. 草地和荒漠生态系统服务功能的形成与调控机制. 植物生态学报, 2014, 38(2): 93-102. |
| [25] |
何玉惠, 刘新平, 谢忠奎. 氮素添加对黄土高原荒漠草原草本植物物种多样性和生产力的影响. 中国沙漠, 2015, 35(1): 66-71. |
| [26] |
黄菊莹, 余海龙. 四种荒漠草原植物的生长对不同氮添加水平的响应. 植物生态学报, 2016, 40(2): 165-176. |
| [27] |
杜忠毓, 安慧, 文志林, 王波, 张馨文. 荒漠草原植物群落结构及其稳定性对增水和增氮的响应. 生态学报, 2021, 41(6): 2359-2371. |
| [28] |
张馨文, 安慧, 刘小平, 文志林, 王波, 杜忠毓. 短期氮添加对荒漠草原植物群落组成及稳定性的影响. 生态学杂志, 2021, 40(8): 2400-2409. |
| [29] |
王攀, 朱湾湾, 牛玉斌, 樊瑾, 余海龙, 赖江山, 黄菊莹. 氮添加对荒漠草原植物群落组成与微生物生物量生态化学计量特征的影响. 植物生态学报, 2019, 43(5): 427-436. |
| [30] |
顾峰雪, 黄玫, 张远东, 闫慧敏, 李洁, 郭瑞, 钟秀丽. 1961-2010年中国区域氮沉降时空格局模拟研究. 生态学报, 2016, 36(12): 3591-3600. |
| [31] |
Shi Y, Sheng L X, Wang Z Q, Zhang X Y, He N P, Yu Q. Responses of soil enzyme activity and microbial community compositions to nitrogen addition in bulk and microaggregate soil in the temperate steppe of Inner Mongolia. Eurasian Soil Science, 2016, 49(10): 1149-1160. |
| [32] |
姚天华, 朱志红, 李英年, 潘石玉, 孔彬彬, 卫欣华, 杜家丽. 功能多样性和功能冗余对高寒草甸群落稳定性的影响. 生态学报, 2016, 36(6): 1547-1558. |
| [33] |
孔彬彬, 卫欣华, 杜家丽, 李英年, 朱志红. 刈割和施肥对高寒草甸物种多样性和功能多样性时间动态及其关系的影响. 植物生态学报, 2016, 40(3): 187-199. |
| [34] |
Lepš J, De Bello F, Lavorel S, Berman S. Quantifying and interpreting functional diversity of natural communities: Practical considerations matter. Preslia, 2006, 78(4): 481-501. |
| [35] |
Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301. |
| [36] |
Laliberté E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305. |
| [37] |
Lavorel S, Grigulis K, Mcintyre S, Williams N S G, Garden D, Dorrough J, Berman S, Quétier F, Thébault A, Bonis A. Assessing functional diversity in the field-methodology matters!. Functional Ecology, 2008, 22(1): 134-147. |
| [38] |
Isbell F, Craven D, Connolly J, Loreau M, Schmid B, Beierkuhnlein C, Bezemer T M, Bonin C, Bruelheide H, De Luca E, Ebeling A, Griffin J N, Guo Q F, Hautier Y, Hector A, Jentsch A, Kreyling J, Lanta V, Manning P, Meyer S T, Mori A S, Naeem S, Niklaus P A, Polley H W, Reich P B, Roscher C, Seabloom E W, Smith M D, Thakur M P, Tilman D, Tracy B F, Van Der Putten W H, Van Ruijven J, Weigelt A, Weisser W W, Wilsey B, Eisenhauer N. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature, 2015, 526(7574): 574-577. |
| [39] |
Cadotte M W, Carscadden K, Mirotchnick N. Beyond species: functional diversity and the maintenance of ecological processes and services. Journal of Applied Ecology, 2011, 48(5): 1079-1087. |
| [40] |
Reich P B. The world-wide 'fast-slow' plant economics spectrum: a traits manifesto. Journal of Ecology, 2014, 102(2): 275-301. |
| [41] |
Howard A L. Handbook of structural equation modeling. Structural Equation Modeling: A Multidisciplinary Journal, 2013, 20(2): 354-360. |
| [42] |
毛庆功, 鲁显楷, 陈浩, 莫江明. 陆地生态系统植物多样性对矿质元素输入的响应. 生态学报, 2015, 35(17): 5884-5897. |
| [43] |
杨祥祥, 李梦琦, 何兴东, 尤万学, 余殿, 张彩华, 陈娜. 沙地植物群落功能多样性对物种多样性的影响. 南开大学学报: 自然科学版, 2020, 53(4): 75-80. |
| [44] |
Hooper D U, Chapin Ⅲ F S, Ewel J J, Hector A, Inchausti P, Lavorel S, Lawton J H, Lodge D M, Loreau M, Naeem S, Schmid B, Setälä H, Symstad A J, Vandermeer J, Wardle D A. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecological Monographs, 2005, 75(1): 3-35. |
| [45] |
Moonen A C, Bàrberi P. Functional biodiversity: an agroecosystem approach. Agriculture, Ecosystems & Environment, 2008, 127(1/2): 7-21. |
| [46] |
Niu K C, Choler P, De Bello F, Mirotchnick N, Du G Z, Sun S C. Fertilization decreases species diversity but increases functional diversity: a three-year experiment in a Tibetan alpine meadow. Agriculture, Ecosystems & Environment, 2014, 182: 106-112. |
| [47] |
Mayfield M M, Levine J M. Opposing effects of competitive exclusion on the phylogenetic structure of communities. Ecology Letters, 2010, 13(9): 1085-1093. |
| [48] |
Mason N W H, De Bello F, Doležal J, Lepš J. Niche overlap reveals the effects of competition, disturbance and contrasting assembly processes in experimental grassland communities. Journal of Ecology, 2011, 99(3): 788-796. |
| [49] |
梁潇洒, 马望, 王洪义, 雒文涛, 王正文. 呼伦贝尔草原3种植物的功能性状对氮磷添加的响应. 中国草地学报, 2019, 41(1): 61-67. |
| [50] |
张全国, 张大勇. 生物多样性与生态系统功能: 最新的进展与动向. 生物多样性, 2003, 11(5): 351-363. |
| [51] |
De Bello F, Lepš J, Sebastià M T. Variations in species and functional plant diversity along climatic and grazing gradients. Ecography, 2006, 29(6): 801-810. |
| [52] |
Mayfield M M, Boni M F, Daily G C, Ackerly D. Species and functional diversity of native and human-dominated plant communities. Ecology, 2005, 86(9): 2365-2372. |
| [53] |
吕亭亭, 王平, 燕红, 张稳, 廖桂项, 姜海波, 邹畅林, 盛连喜. 草甸和沼泽植物群落功能多样性与生产力的关系. 植物生态学报, 2014, 38(5): 405-416. |
| [54] |
武倩. 长期增温和氮素添加对荒漠草原植物群落稳定性的影响[D]. 呼和浩特: 内蒙古农业大学, 2019.
|
| [55] |
景明慧, 贾晓彤, 张运龙, 曹娟, 周伟, 王杰, 庾强. 长期氮添加对内蒙古典型草原植物地上、地下生物量及根冠比的影响. 生态学杂志, 2020, 39(10): 3185-3193. |
| [56] |
Xu Z W, Li M H, Zimmermann N E, Li S P, Li H, Ren H Y, Sun H, Han X G, Jiang Y, Jiang L. Plant functional diversity modulates global environmental change effects on grassland productivity. Journal of Ecology, 2018, 106(5): 1941-1951. |
| [57] |
Griffin J N, Méndez V, Johnson A F, Jenkins S R, Foggo A. Functional diversity predicts overyielding effect of species combination on primary productivity. Oikos, 2009, 118(1): 37-44. |
| [58] |
李晓刚, 朱志红, 周晓松, 袁芙蓉, 樊瑞俭, 许曼丽. 刈割、施肥和浇水对高寒草甸物种多样性、功能多样性与初级生产力关系的影响. 植物生态学报, 2011, 35(11): 1136-1147. |
| [59] |
Petche O L, Hector A, Gaston K J. How do different measures of functional diversity perform?. Ecology, 2004, 85(3): 847-857. |
| [60] |
Finegan B, Peña-Claros M, De Oliveira A, Ascarrunz N, Bret-Harte M S, Carreño-Rocabado G, Casanoves F, Díaz S, Velepucha P E, Fernandez F, Licona J C, Lorenzo L, Negret B S, Vaz M, Poorter L. Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses. Journal of Ecology, 2015, 103(1): 191-201. |