 2023, Vol. 43
2023, Vol. 43文章信息
- 王苗苗, 王绍强, 陈斌, 张心怡, 赵健
- WANG Miaomiao, WANG Shaoqiang, CHEN Bin, ZHANG Xinyi, ZHAO Jian
- 2020-2050年CO2施肥效应促进全球陆地生态系统碳吸收
- CO2 fertilization promoting the carbon uptake of global terrestrial ecosystems in 2020-2050
- 生态学报. 2023, 43(6): 2408-2418
- Acta Ecologica Sinica. 2023, 43(6): 2408-2418
- http://dx.doi.org/10.5846/stxb202107051782
-
文章历史
- 收稿日期: 2021-07-05
- 网络出版日期: 2022-11-08





2. 中国科学院地理科学与资源研究所, 北京 100101
2. Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China
大气中CO2浓度升高导致的全球气候变暖是人类共同关注的问题, 根据未来不同CO2排放情景的预估表明大气中累积的CO2排放量将进一步增加, 年增长率约为0.002%[1—2], CO2浓度的增加将会促进植被生长并增加陆地生态系统生产力[3—4]。总初级生产力(Gross Primary Productivity, GPP)表示单位时间、单位面积内植物通过光合作用所固定的有机碳总量, 是陆地生态系统碳循环的基础[5—7], 预估未来气候变化条件下CO2施肥效应对全球陆地生态系统GPP的影响具有重大意义。
陆地生态系统模型是研究区域陆地生态系统生产力最重要的研究手段[6, 8]。叶面积指数(Leaf area index, LAI)是陆地生态系统模型的关键输入参数[9—10]。LAI定义为单位面积内总绿叶面积的一半[9], 控制着许多冠层尺度的生化过程, 如辐射吸收、光合作用、降水截留和蒸散等。陆地生态系统模型根据LAI的获取方式分为诊断模型和预测模型。诊断模型基于遥感的LAI作为模型输入数据, 并结合复杂程度各异的经验算法, 用于模拟区域乃至全球陆地生态系统碳水通量。而预测模型一般通过给定的气候和土壤条件模拟植被结构参数(LAI)和生长过程, 进而模拟陆地生态系统碳循环[11]。有研究表明, 相对于预测模型, 基于LAI驱动的诊断模型BEPS(Boreal Ecosystem Productivity Simulator)模型可以很好地用于模拟全球陆地生态系统生产力[12]。
许多研究表明区域和全球植被正在变绿, 大气CO2浓度增加被认为是全球变绿, 即LAI增加的主要原因之一[13—15]。比如, 大气CO2富集实验(FACE)表明CO2施肥效应对LAI影响大约为CO2每升高0.1%, LAI增加约0.6%—24.1%[16]。TRENDY2模型模拟结果表明CO2每升高0.1%, 全球LAI对应增加10.2%—20.7%[3]。2001—2017年CO2施肥效应对全球LAI的影响为CO2每升高0.1%, LAI大约增加13.4% (除热带常绿针叶林外)[17]。而CO2施肥效应对LAI的影响被认为是模型模拟陆地生态系统碳循环对气候变化影响的主要不确定性因子[18]。因此, 准确预估未来气候变化条件下CO2施肥效应对LAI的影响对全球陆地生态系统碳循环研究至关重要。但是未来气候变化条件下, CO2浓度增加对全球LAI的效应及其对GPP的影响仍不明晰。
陆地生态系统碳循环对大气CO2浓度增加的响应主要是通过叶片羧化速率的影响, 大气CO2浓度增加通过刺激叶片羧化速率增加GPP和NPP, 进而通过增加碳分配到叶片的量而增加LAI[19]。相对于积温法和碳动态平衡法, 利用模型直接估算CO2施肥效应对全球LAI的影响, 有利于减少未来全球陆地生态系统生产力的不确定性。其中, Farquhar模型通过羧化和电子传递能力间的共同限制模拟植被的光合作用, 可以用于模拟光照和CO2对光合作用的影响[18]。Wang等[17]研究表明Farquhar模型可以有效的用于模拟CO2施肥效应对全球LAI年际变化趋势的影响, 结合气候因子、土壤因子和Farquhar模型可以有效重构过去全球LAI的季节和年际变化趋势, 为有效地预估未来全球LAI时空格局提供了新思路, 并且可以为生态系统模型估算未来陆地生态系统生产力提供可靠的LAI输入源。Wang等[17]构建的方法成功的用于重构过去全球LAI季节和年际变化趋势, 暂未用于构造未来气候变化条件下全球陆地生态系统的LAI, 且未用于探讨CO2施肥效应导致的LAI增加量对未来GPP年际变化趋势的影响。因此, 本研究基于未来气候情景数据, 结合Farquhar模型, 采用Wang等[17]的全球LAI构造方法, 生成2020—2050年不同气候情景下全球月尺度的LAI数据, 并结合BEPS模型, 预估2020—2050年全球陆地生态系统GPP时空格局, 定量化研究未来气候变化条件下, CO2浓度增加对LAI的作用及CO2施肥效应导致的LAI增加量对GPP年际变化趋势的影响。
1 数据来源 1.1 气象数据研究中使用CMIP5(Coupled Model Intercomparison Project Phase 5)(https://esgf-node.llnl.gov/projects/cmip5/)中的MRI-CGCM3模型结果作为未来预估的气候驱动数据。在CMIP5界面的高级检索中选择MRI-CGCM3模型中对应的气象数据。为了对未来气候作出评估, IPCC为第五次评估报告制定了四个代表性的代表浓度路径(RCP:Representative concentration Pathways)[20]。研究中采用RCP2.6、RCP4.5和RCP8.5三个气候情景下的温度、降水、辐射和相对湿度来模拟未来全球陆地生态系统的LAI和GPP时空格局, 该数据集的空间分辨率1.125°(经度)×1.12148°(纬度)。本研究将所有的气候数据采用最邻近法统一重采样成0.5°×0.5°空间分辨率的栅格数据。
1.2 遥感LAI数据本研究使用的遥感LAI是全球GLOBMAP_v3数据(https://zenodo.org/badge/latestdoi/269540368)。其中, 2001—2018年的全球LAI数据是基于全球4尺度几何光学模型, 通过MOD09A1 C6地表反射率产品, 基于GLOBCARBON LAI算法的视角中反演得到的全球LAI数据产品[21]。GLOBMAP LAI数据集的空间分辨率为0.072727°×0.072727°, 时间分辨率为8 d。研究中心将LAI数据重采样成0.5°×0.5°空间分辨率数据。
2 研究方法 2.1 2020—2050年LAI计算方法Wang等[17]研究表明使用温度、降水、辐射和土壤湿度可以解释93.0%全球LAI季节变化趋势, 且Farquhar模型可以有效地用于模拟CO2施肥效应对全球LAI的影响。本研究采用Wang等[17]的研究方法, 结合温度、降水、辐射、土壤湿度和Farquhar模型, 构建2020—2050年月尺度全球陆地生态系统LAI数据集。研究按RCP2.6、RCP4.5和RCP8.5三个气候情景数据, 以及是否考虑CO2施肥效应对LAI年际变化趋势的影响, 共生成了6套不同的LAI产品, 分别为RCP2.6、RCP4.5和RCP8.5气候情景下对应的RLAI、RLAIco2数据集。
2.1.1 RLAI计算首先利用2001—2018年月尺度GLOBMAP LAI数据在不同季节分别建立LAI和气候因子(温度、降水、辐射和土壤湿度)的多元线性回归方程, 得到各个因子的回归系数。并利用得到的多元回归系数, 分别计算每个季节内对应月份的LAI值, 得到2020—2050年全球月尺度RLAI数据集。
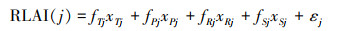
|
(1) |
其中RLAIj季节j内的仅由气候因子计算得到的月LAI值;xTj、xPj、xRj和xSj代表对应季节的月均大气温度、总降水量、总太阳辐射和月均土壤湿度;fTj、fPj、fRj和fSj为对应的回归系数, 详见Wang等[17]。
2.1.2 CO2施肥效应对LAI的影响2020—2050年RCP2.6、RCP4.5和RCP8.5三个气候情景下的CO2浓度分别升高至0.44%、0.48%和0.54%, 相对于2020年分别增加0.03%、0.07%和0.12%, 未来CO2年际变化趋势如图 1所示。研究将以2001—2018年LAI和GPP均值为基准, 探讨未来气候变化条件下, CO2浓度升高对LAI的作用及其对GPP的影响。

|
| 图 1 2020—2050年CO2浓度变化量 Fig. 1 The time series of CO2 concentrations during 2020—2050 |
研究采用Farquhar模型[22]探讨未来气候变化条件下CO2施肥效应对全球LAI的影响。
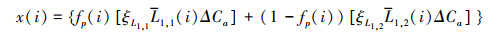
|
(2) |
式中, i代表年份, x(i)为大气CO2浓度对LAI的影响。fP为受电子传递速率限制的冠层光合作用的系数。ΔCa代表CO2浓度的年际变化量。L1, 1和L1, 2代表CO2浓度变化对LAI的影响系数, ξL1, 1和ξL1, 2分别为L1, 1和L1, 2日变化和季节变化的校正系数。各参数取值细节详见Ju等[22]。
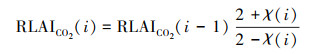
|
(3) |
将CO2施肥效应对LAI年际变化趋势的影响嵌入到RLAI中, 得到RLAICO2。其中RLAI在2001年的值作为RLAICO2第一年的值。关于CO2施肥效应对LAI的影响细节详见Ju等[23]和Wang等[17]。由于Wang等[17]研究显示热带常绿阔叶林LAI的季节性变化并不明显, 以及该区域常年多云多雨的特点, 并不适合基于逐像元的LAI构建。因此, 本研究不包括热带常绿阔叶林。
2.2 BEPS模型简介BEPS模型是基于FOREST-BGC模型发展起来的生态过程机理模型。BEPS模型最主要的特点是在模拟冠层尺度的光合作用时将冠层叶片分为阴叶和阳叶[24], 分别模拟对应的光合作用。BEPS模型已被逐渐推广应用于中国[25, 26]、北美[27, 28]以及全球[5, 9]。许多研究都使用通量站点观测数据验证BEPS模型模拟结果的准确性, 结果表明BEPS模型可以很好地用于模拟区域及全球陆地生态系统生产力[9, 29]。
BEPS模型中冠层的光合速率计算为:
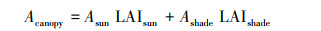
|
(4) |
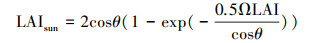
|
(5) |
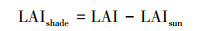
|
(6) |
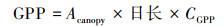
|
(7) |
式中, Acanopy、Asun和Ashade分别代表整个冠层、阳叶和阴叶的日光合速率;LAIsun和LAIshade分别代表阳叶和阴叶的叶面积指数;θ代表太阳高度角;Ω代表叶片聚集度(与地表覆盖类型有关);CGPP代表转换系数。
基于上述得到的2020—2050年全球月尺度LAI数据集, 结合RCP2.6、RCP4.5和RCP8.5气候情景下的温度、降水、辐射、相对湿度等数据, 运用BEPS模型分别模拟三种气候情景下2020—2050年全球陆地生态系统GPP的时空格局。
3 结果 3.1 2020—2050年全球LAI年际变化趋势2020—2050年仅有气候数据得到的LAI(RLAI)在RCP2.6、RCP4.5和RCP8.5气候情景下均无显著的变化趋势(P>0.5), 而当加入CO2浓度影响后得到的LAI(RLAIco2)年际变化趋势分别为0.002、0.003和0.005 m-2m-2a-1(图 2), 表明相对于未来气候变化, CO2施肥效应仍然是LAI增加的主要原因之一。

|
| 图 2 全球LAI年际变化趋势 Fig. 2 Area weighted annual averaged LAI time series |
2020—2050年在三种气候情景下, 仅基于气候因子计算得到的LAI的年际变化趋势在空间分布上均不明显(图 3)。部分显著增加区域的LAI年际变化趋势小于0.003 m-2m-2a-1, 且在南美洲东部的部分地区、印度部分地区以及北美洲部分地区, LAI有降低的趋势。当在加入CO2浓度的影响后, 在RCP2.6气候情景下, 全球大部分植被覆盖区的LAI呈现显著的增加趋势, 但年际变化趋势小于0.003 m-2m-2a-1。对于RCP4.5, 全球大部分植被覆盖区的LAI均具有显著增加趋势, 尤其在亚洲东南部、北美洲中东部、南美洲中部以及非洲中部地区, LAI的年均增加趋势大于0.006 m-2m-2a-1。对于RCP8.5, 全球LAI显著增加区域均高于RCP2.6和RCP4.5, 其中在亚洲东南部、欧洲部分区域、北美洲东南部以及南美洲的中南部地区, LAI的年际变化趋势大于0.009 m-2m-2a-1。

|
| 图 3 2020—2050年RLAI和RLAIco2年际变化趋势空间分布 Fig. 3 The spatial pattern of the trends in the global annual RLAI and RLAIco2 during 2020—2050 |
基于BEPS模型模拟结果表明2020—2050年在RCP2.6、RCP4.5和RCP8.5三种气候情景下, 仅由气候因子得到的LAI(RLAI)驱动BEPS模型得到的结果显示2020—2050年全球年均GPP值分别为68.8 PgC/a、70.5 PgC/a和70.6 PgC/a, 此时三种气候情景下GPP的年际变化趋势分别为0.14 PgC/a、0.20 PgC/a和0.27 PgC/a(图 4)。当使用考虑了CO2施肥效应对LAI影响后得到的RLAIco2驱动模型后得到的2020—2050年全球陆地生态系统在不同气候情景下的年均GPP值分别为73.2 PgC/a、72.9 PgC/a和76.7 PgC/a, 对应的GPP年际变化趋势分别为0.42 PgC/a、0.39 PgC/a和0.63 PgC/a。其中, CO2施肥效应导致的LAI的增加对GPP年际变化在三个不同气候情景下的贡献量分别为66.7%、48.7%和57.1%。说明对于未来气候变化条件下, CO2施肥效应导致的LAI增加量将显著促进全球陆地生态系统碳吸收。

|
| 图 4 不同气候情景下全球GPP年际变化趋势 Fig. 4 Aunnual GPP time series in different climate scenario |
2020—2050年全球GPP年际变化趋势的空间分布表明在RCP2.6、RCP4.5和RCP8.5气候情景下, 2020—2050年全球陆地生态系统大部分区域的GPP年际变化趋势均呈显著上升趋势(图 5)。在RCP2.6气候情景下, 2020—2050年全球陆地生态系统GPP的年际变化趋势的空间分布并不明显, 此时大部分GPP显著变化区域的GPP年均增量小于6 gC m-2 a-1。对于RCP4.5气候情景下, 当使用RLAIco2驱动模型得到的GPP较基于RLAI的模拟结果增加面积显著增多, 其中在南美洲北部、非洲中部地区以及北半球的中—高纬度地区GPP的增加趋势大于9 gC m-2 a-1。

|
| 图 5 2020—2050年全球陆地生态系统GPP年际变化趋势空间分布 Fig. 5 The spatial pattern of the trends in the global annual GPP during 2020—2050 |
在RCP8.5气候情景下, 由RLAI驱动的模拟结果表明, 在南美洲中北部、非洲中部地区、亚洲东南部、欧洲、北美洲中东部以及北半球高纬度部分地区, GPP的年际变化趋势大于12 gC m-2 a-1。基于RLAIco2驱动的模型结果表明, 在南美洲中北部、非洲中部、亚洲东南部、欧洲、北美洲和北半球高纬度部分地区的GPP年均增加量大于18 gC m-2 a-1, 较RCP2.6和RCP4.5显著提高。表明基于RLAIco2驱动的结果较基于RLAI驱动得到的结果在GPP显著增加趋势上有明显增加。
3.3 CO2施肥效应对全球LAI和GPP的影响对于全球LAI, 在RCP2.6、RCP4.5和RCP8.5气候情景下, CO2施肥效应导致的LAI相对于2001—2018年均值的增加量分别为0.04 m-2m-2、0.10 m-2m-2和0.15 m-2m-2(图 6)。CO2施肥效应对LAI的影响随着年份的增加而增加, 然而在2035年以后CO2施肥效应对LAI的影响有所减弱。在三个气候情景中, RCP8.5的CO2施肥效应对LAI的影响高于RCP4.5和RCP2.6, 结合对应气候情景下的CO2浓度增加量, CO2浓度增加对LAI的施肥效应为CO2每升高0.1%, LAI平均增加约8.1%—9.2%(图 6)。

|
| 图 6 CO2施肥效应对LAI的影响 Fig. 6 The effect of CO2 fertilization on LAI |
对于全球GPP, 在RCP2.6、RCP4.5和RCP8.5气候情景下, 2020—2050年CO2施肥效应导致的LAI增加量使GPP相对于2001—2018年均值的增加量分别为9.83 PgC、6.99 PgC和11.4 PgC(图 7)。2020—2050年三个气候变化情景下, CO2施肥效应导致的LAI增加量对GPP的影响随着年份的增大而增加, 三个气候情景CO2施肥效应相对大小为RCP2.6>RCP4.5>RCP8.5, CO2浓度每升高0.1%导致的LAI增加量使GPP对应增加7.9%—14.6%(图 7)。

|
| 图 7 CO2施肥效应对GPP的影响 Fig. 7 The effect of CO2 fertilization on GPP |
本研究结果表明在RCP2.6、RCP4.5和RCP8.5气候情景下, 2020—2050年由CO2施肥效应导致LAI的平均增加量分别为0.05、0.11和0.17 m2 m-2, CO2施肥效应为CO2每增加0.1%, LAI增加3.9%—9.2%。该研究结果与大气CO2富集实验(FACE)的0.6%—24.1% [16]、TRENDY2模型模拟结果10.2%—20.7% [3]等的研究结果相近。另外, 当研究未考虑CO2施肥效应时, 2020—2050年不同气候情景下的LAI并无显著变化趋势, 且由气候变化导致南美洲东部的部分地区、印度部分地区以及北美洲部分地区的LAI有降低趋势。当加入CO2施肥效应后的LAI(RLAIco2)具有显著变化趋势, 年际变化趋势分别为0.002、0.003和0.005 m-2m-2a-1, 表明CO2施肥效应仍是未来气候变化条件下导致全球变绿的主要原因之一。Piao等[14]研究均表明大气CO2浓度增加是全球陆地生态系统变绿的主要原因之一, 与本研究的结果一致。
另外, 本研究中预估的2020—2050年全球陆地生态系统LAI年际变化较为明显的区域出现在亚洲东南部、欧洲、北美洲东部、南美洲中部和非洲中部区域, 这也与2000—2017年全球LAI显著变化区域较为一致[9]。其中, Piao等[30]研究表明在亚洲东南部地区CO2施肥效应可以解释85.0%中国陆地生态系统生长季内年均LAI的年际变化趋势。而基于全球陆地生态系统模型模拟结果表明CO2施肥效应可以解释70.0%的全球LAI的年际增加趋势[3], 这些研究结果均可支持本研究的结果。
4.2 CO2施肥效应导致的LAI的增加促进全球陆地生态系统碳吸收本研究结果表明2020—2050年在RCP2.6、RCP4.5和RCP8.5气候情景下, 由RLAI驱动BEPS模型得到的2020—2050年GPP的年际变化趋势分别为0.14 PgC/a、0.20 PgC/a和0.27 PgC/a, 由RLAIco2驱动模型得到的GPP年际变化趋势分别为0.42 PgC/a、0.39 PgC/a和PgC/a(图 4)。三个气候情景下, CO2施肥效应导致的LAI增加量对全球陆地生态系统GPP年际变化趋势的贡献分别为66.7%、48.7%和57.1%。CO2每增加0.1%, 由LAI增加导致的GPP增加量约为7.9%—14.6%。朱再春等[31]基于CMIP5气候模式, 预估未来陆地生态系统生产力, 结果表明陆地生态系统NPP总量增加主要由大气CO2浓度上升驱动, 其他环境因子的影响相对较弱。Chen等[9]研究表明1981—2016年全球LAI的增加使陆地生态系统碳汇增强12.4%, 也与本研究结果相近。另外, Cao等[32]研究表明在全球范围内, CO2施肥效应随CO2浓度增加而降低, 当CO2浓度超过0.5%时, CO2浓度升高对光合作用的刺激作用减弱, 但依旧会促进光合作用, 这也支持研究中2035年以后, CO2施肥效应对LAI的影响有所减弱的结果。
4.3 不确定性分析本研究基于Farquhar模型和气候因子预估未来气候变化条件下CO2施肥效应对全球LAI的影响, 并探讨CO2施肥效应对LAI的影响对GPP的作用, 研究还存在一定的不确定性。首先, 遥感LAI数据显示2001—2018年LAI的年际变化趋势为0.008 m-2m-2a-1, 高于对未来LAI的模拟结果(图 2)。这主要是由于遥感的LAI包含了人为原因导致的LAI增加量, 本研究重点在于探讨CO2施肥效应和气候变化的影响, 暂未考虑人为因素的影响, 可能会低估未来LAI的年际变化趋势。而且未考虑或简化考虑生态系统管理(如森林管理、农业灌溉等)对碳循环的影响, 也会对研究结果造成一定的误差[33]。
其次, CO2施肥效应对GPP的增加中有一部分归因于叶水平效应, 即在恒定叶面积下发生的CO2直接诱导的GPP增加, 另一部分是由于CO2施肥效应导致LAI增加, 进而促使GPP增加[4, 34]。本研究侧重于探讨CO2施肥效应导致LAI增加量对全球GPP的影响, 并未考虑叶水平效应。另外, BEPS模型中也包含了CO2浓度增加对气孔导度的影响, 这方面也会影响陆地生态系统植被的光合作用, 但是本研究中暂时未分离出该部分对全球GPP的影响。
最后, 本研究仅基于MRI-CGCM3未来气候情景数据预估2020—2050年全球陆地生态系统的LAI和GPP, 未结合其它气候情景数据结果进行对比分析, 因此本研究仅代表MRI-CGCM3气候情景下的结果。
5 结论本研究基于2020—2050年RCP2.6、RCP4.5和RCP8.5三个气候情景下的温度、降水、辐射和土壤湿度数据, 结合Farquhar模型和BEPS模型, 探讨2020—2050年CO2施肥效应对全球陆地生态系统LAI和GPP的影响, 主要研究结论有:
(1) 2020—2050年仅有气候数据得到的LAI(RLAI)数据至2050年在三个气候情景下均无显著的变化趋势(P>0.5), 当加入CO2施肥效应的影响后得到的LAI(RLAIco2)在RCP2.6、RCP4.5和RCP8.5下, 年际变化趋势分别为0.002、0.003和0.005 m-2m-2a-1, 表明在未来气候变化情景下, CO2施肥效应仍然是全球LAI增加的主要原因之一。
(2) 2020—2050年三个气候情景下CO2施肥效应对LAI的影响为CO2每增加0.1%, LAI平均增加约8.1%—9.2%, 由此导致GPP对应增加7.9%—14.6%。表明未来气候变化条件下, CO2施肥效应仍是全球GPP增加的重要因素。
(3) CO2施肥效应导致的LAI的增加对GPP年际变化在三个不同气候情景下的贡献量分别为66.67%、48.72%和57.14%, 表明在未来气候变化条件下, CO2施肥效应导致的LAI增加量将显著促进全球陆地生态系统碳吸收。
| [1] |
Wang W L, Ciais P, Nemani R R, Canadell J G, Piao S, Sitch S, White M A, Hashimoto H, Milesi C, Myneni R B. Variations in atmospheric CO2 growth rates coupled with tropical temperature. Proceedings of the National Academy of sciences of the United States of America, 2013, 110(32): 13061-6. DOI:10.1073/pnas.1219683110 |
| [2] |
Friedlingstein P. Carbon cycle feedbacks and future climate change. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, 2015, 373(2054): 20140421. DOI:10.1098/rsta.2014.0421 |
| [3] |
Zhu Z C, Piao S L, Myneni R B, Huang M T, Zeng Z Z, Canadell J G, Ciais P, Sitch S, Friedlingstein P, Arneth A, Cao C X, Cheng L, Kato E, Koven C, Li Y, Lian X, Liu Y W, Liu R G, Mao J F, Pan Y Z, Peng S S, Peñelas J, Poulter B, Pugh T A M, Stocker B D, Viovy N, Wang X H, Wang Y P, Xiao Z Q, Yang H, Zaehle S, Zeng N. Greening of the Earth and its drivers. Nature Climate Change, 2016, 6(8): 791-795. DOI:10.1038/nclimate3004 |
| [4] |
Haverd V, Smith B, Canadell J G, Cuntz M, Mikaloff-Fletcher S, Farquhar G, Woodgate W, Briggs P R, Trudinger C M. Higher than expected CO2 fertilization inferred from leaf to global observations. Global Change Biology, 2020, 26(4): 2390-2402. DOI:10.1111/gcb.14950 |
| [5] |
Wang M M, Wang S Q, Zhao J, Ju W M, Hao Z. Global positive gross primary productivity extremes and climate contributions during 1982-2016. Science of the Total Environment, 2021, 774: 145703. DOI:10.1016/j.scitotenv.2021.145703 |
| [6] |
Sun Z Y, Wang X F, Zhang X R, Tani H, Guo E L, Yin S, Zhang T Y. Evaluating and comparing remote sensing terrestrial GPP models for their response to climate variability and CO2 trends. Science of the Total Environment, 2019, 668: 696-713. DOI:10.1016/j.scitotenv.2019.03.025 |
| [7] |
Beer C, Reichstein M, Tomelleri E, Ciais P, Jung M, Carvalhais N, Rödenbeck C, Arain M A, Baldocchi D, Bonan G B, Bondeau A, Cescatti A, Lasslop G, Lindroth A, Lomas M, Luyssaert S, Margolis H, Oleson K W, Roupsard O, Veenendaal E, Viovy N, Williams C, Woodward F I, Papale D. Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science, 2010, 329(5993): 834-838. DOI:10.1126/science.1184984 |
| [8] |
于贵瑞, 张黎, 何洪林, 杨萌. 大尺度陆地生态系统动态变化与空间变异的过程模型及模拟系统. 应用生态学报, 2021, 32(8): 2653-2665. DOI:10.13287/j.1001-9332.202108.040 |
| [9] |
Chen J M, Ju W M, Ciais P, Viovy N, Liu R G, Liu Y, Lu X H. Vegetation structural change since 1981 significantly enhanced the terrestrial carbon sink. Nature Communications, 2019, 10(1): 4259. DOI:10.1038/s41467-019-12257-8 |
| [10] |
White M A, Thornton P E, Running S W, Nemani R R. Parameterization and sensitivity analysis of the BIOME-BGC terrestrial ecosystem model: net primary production controls. Earth Interactions, 2000, 4(3): 1-85. DOI:10.1175/1087-3562(2000)004<0003:PASAOT>2.0.CO;2 |
| [11] |
Le Quéré C, Andrew R M, Friedlingstein P, Sitch S, Pongratz J, Manning A C, Korsbakken J I, Peters G P, Canadell J G, Jackson R B, Boden T A, Tans P P, Andrews O D, Arora V K, Bakker D C E, Barbero L, Becker M, Betts R A, Bopp L, Chevallier F, Chini L P, Ciais P, Cosca C E, Cross J, Currie K, Gasser T, Harris I, Hauck J, Haverd V, Houghton R A, Hunt C W, Hurtt G, Ilyina T, Jain A K, Kato E, Kautz M, Keeling R F, Klein Goldewijk K, Körtzinger A, Landschützer P, Lefèvre N, Lenton A, Lienert S, Lima I, Lombardozzi D, Metzl N, Millero F, Monteiro P M S, Munro D R, Nabel J E M S, Nakaoka S I, Nojiri Y, Padin X A, Peregon A, Pfeil B, Pierrot D, Poulter B, Rehder G, Reimer J, Rödenbeck C, Schwinger J, Séférian R, Skjelvan I, Stocker B D, Tian H Q, Tilbrook B, Tubiello F N, Van Der Laan-Luijkx I T, Van Der Werf G R, Van Heuven S, Viovy N, Vuichard N, Walker A P, Watson A J, Wiltshire A J, Zaehle S, Zhu D. Global carbon budget 2017. Earth System Science Data, 2018, 10(1): 405-448. DOI:10.5194/essd-10-405-2018 |
| [12] |
Sun Z Y, Wang X F, Yamamoto H, Tani H, Nie T Z. The effects of spatiotemporal patterns of atmospheric CO2 concentration on terrestrial gross primary productivity estimation. Climatic Change, 2020, 163(2): 913-930. DOI:10.1007/s10584-020-02903-2 |
| [13] |
Chen S Y, Zhang Y L, Wu Q L, Liu S H, Song C H, Xiao J F, Band L E, Vose J M. Vegetation structural change and CO2 fertilization more than offset gross primary production decline caused by reduced solar radiation in China. Agricultural and Forest Meteorology, 2021, 296: 108207. DOI:10.1016/j.agrformet.2020.108207 |
| [14] |
Piao S L, Wang X H, Park T, Chen C, Lian X, He Y, Bjerke J W, Chen A P, Ciais P, Tømmervik H, Nemani R R, Myneni R B. Characteristics, drivers and feedbacks of global greening. Nature Reviews Earth & Environment, 2020, 1(1): 14-27. |
| [15] |
Los S O. Analysis of trends in fused AVHRR and MODIS NDVI data for 1982-2006: indication for a CO2 fertilization effect in global vegetation. Global Biogeochemical Cycles, 2013, 27(2): 318-330. DOI:10.1002/gbc.20027 |
| [16] |
Norby R J, Delucia E H, Gielen B, Calfapietra C, Giardina C P, King J S, Ledford J, Mccarthy H R, Moore D J P, Ceulemans R, De Angelis P, Finzi A C, Karnosky D F, Kubiske M E, Lukac M, Pregitzer K S, Scarascia-Mugnozza G E, Schlesinger W H, Oren R. Forest response to elevated CO2 is conserved across a broad range of productivity. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(50): 18052-18056. DOI:10.1073/pnas.0509478102 |
| [17] |
Wang M M, Chen J M, Wang S Q. Reconstructing the seasonality and trend in global Leaf Area Index during 2001-2017 for prognostic modeling. Journal of Geophysical Research: Biogeosciences, 2020, 125(9): e2020JG005698. |
| [18] |
Huntzinger D N, Michalak A M, Schwalm C, Ciais P, King A W, Fang Y, Schaefer K, Wei Y, Cook R B, Fisher J B, Hayes D, Huang M, Ito A, Jain A K, Lei H, Lu C, Maignan F, Mao J, Parazoo N, Peng S, Poulter B, Ricciuto D, Shi X, Tian H, Wang W, Zeng N, Zhao F. Uncertainty in the response of terrestrial carbon sink to environmental drivers undermines carbon-climate feedback predictions. Scientific Reports, 2017, 7(1): 4765. DOI:10.1038/s41598-017-03818-2 |
| [19] |
Heimann M, Reichstein M. Terrestrial ecosystem carbon dynamics and climate feedbacks. Nature, 2008, 451(7176): 289-292. DOI:10.1038/nature06591 |
| [20] |
Yukimoto S, Adachi Y, Hosaka M, Sakami T, Yoshimura H, Hirabara M, Tanaka T Y, Shindo E, Tsujino H, Deushi M, Mizuta R, Yabu S, Obata A, Nakano H, Koshiro T, Ose T, Kitoh A. A new global climate model of the meteorological research institute: MRI-CGCM3——model description and basic performance. Journal of the Meteorological Society of Japan, 2012, 90A: 23-64. |
| [21] |
Liu Y, Liu R G, Chen J M. Retrospective retrieval of long-term consistent global leaf area index (1981-2011) from combined AVHRR and MODIS data. Journal of Geophysical Research: Biogeosciences, 2012, 117(G4): G04003. |
| [22] |
Farquhar G D, von Caemmerer S, Berry J A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta, 1980, 149(1): 78-90. DOI:10.1007/BF00386231 |
| [23] |
Ju W M, Chen J M, Harvey D, Wang S. Future carbon balance of China's forests under climate change and increasing CO2. Journal of Environmental Management, 2007, 85(3): 538-562. DOI:10.1016/j.jenvman.2006.04.028 |
| [24] |
Chen J M, Liu J, Cihlar J, Goulden M L. Daily canopy photosynthesis model through temporal and spatial scaling for remote sensing applications. Ecological Modelling, 1999, 124(2/3): 99-119. |
| [25] |
Wang M M, Zhao J, Wang S Q, Chen B, Li Z P. Detection and attribution of positive net ecosystem productivity extremes in China's terrestrial ecosystems during 2000-2016. Ecological Indicators, 2021, 132: 108323. DOI:10.1016/j.ecolind.2021.108323 |
| [26] |
Liu Y B, Ju W M, He H L, Wang S Q, Sun R, Zhang Y D. Changes of net primary productivity in China during recent 11 years detected using an ecological model driven by MODIS data. Frontiers of Earth Science, 2013, 7(1): 112-127. DOI:10.1007/s11707-012-0348-5 |
| [27] |
Sprintsin M, Chen J M, Desai A, Gough C M. Evaluation of leaf-to-canopy upscaling methodologies against carbon flux data in North America. Journal of Geophysical Research: Biogeosciences, 2012, 117(G1): G01023. |
| [28] |
Schwalm C R, Williams C A, Schaefer K, Baldocchi D, Black T A, Goldstein A H, Law B E, Oechel W C, Paw K T, Scott R L. Reduction in carbon uptake during turn of the century drought in western North America. Nature Geoscience, 2012, 5(8): 551-556. DOI:10.1038/ngeo1529 |
| [29] |
He H L, Wang S Q, Zhang L, Wang J B, Ren X L, Zhou L, Piao S, Yan H, Ju W M, Gu F X, Yu S Y, Yang Y H, Wang M M, Niu Z, Ge R, Yan H M, Huang M, Zhou G Y, Bai Y F, Xie Z Q, Tang Z Y, Wu B F, Zhang L M, He N P, Wang Q F, Yu G R. Altered trends in carbon uptake in China's terrestrial ecosystems under the enhanced summer monsoon and warming hiatus. National Science Review, 2019, 6(3): 505-514. DOI:10.1093/nsr/nwz021 |
| [30] |
Piao S L, Yin G D, Tan J G, Cheng L, Huang M T, Li Y, Liu R G, Mao J F, Myneni R B, Peng S S, Poulter B, Shi X Y, Xiao Z Q, Zeng N, Zeng Z Z, Wang Y P. Detection and attribution of vegetation greening trend in China over the last 30 years. Global Change Biology, 2015, 21(4): 1601-1609. DOI:10.1111/gcb.12795 |
| [31] |
朱再春, 刘永稳, 刘祯, 朴世龙. CMIP5模式对未来升温情景下全球陆地生态系统净初级生产力变化的预估. 气候变化研究进展, 2018, 14(1): 31-39. |
| [32] |
Cao M K, Woodward F I. Dynamic responses of terrestrial ecosystem carbon cycling to global climate change. Nature, 1998, 393(6682): 249-252. DOI:10.1038/30460 |
| [33] |
朴世龙, 何悦, 王旭辉, 陈发虎. 中国陆地生态系统碳汇估算: 方法、进展、展望. 中国科学: 地球科学, 2022, 52(6): 1010-1020. |
| [34] |
Wang S H, Zhang Y G, Ju W M, Chen J M, Ciais P, Cescatti A, Sardans J, Janssens I A, Wu M S, Berry J A, Campbell E, Fernández-Martínez M, Alkama R, Sitch S, Friedlingstein P, Smith W K, Yuan W P, He W, Lombardozzi D, Kautz M, Zhu D, Lienert S, Kato E, Poulter B, Sanders T G M, Krüger I K, Wang R, Zeng N, Tian H Q, Vuichard N, Jain A K, Wiltshire A, Haverd V, Goll D S, Peñuelas J. Recent global decline of CO2 fertilization effects on vegetation photosynthesis. Science, 2020, 370(6522): 1295-1300. DOI:10.1126/science.abb7772 |