 2023, Vol. 43
2023, Vol. 43文章信息
- 汪正祥, 熊蔚, 邓龙强, 林丽群, 张志麒, 李亭亭
- WANG Zhengxiang, XIONG Wei, DENG Longqiang, LIN Liqun, ZHANG Zhiqi, LI Tingting
- 大九湖沼泽湿地大泥炭藓种群分布特征及其制约因素
- Distribution characteristics of Sphagnum palustre L. population and its influencing factors in Sphagnum wetland of Dajiu Lake
- 生态学报. 2023, 43(6): 2388-2396
- Acta Ecologica Sinica. 2023, 43(6): 2388-2396
- http://dx.doi.org/10.5846/stxb202102250528
-
文章历史
- 收稿日期: 2021-02-25
- 网络出版日期: 2022-11-03





2. 湖北大学资源环境学院, 武汉 430062;
3. 神农架国家公园管理局, 神农架 442421
2. Faculty of Resources and Environmental Science, Hubei University, Wuhan 430062, China;
3. National Park Management Bureau of Shennongjia, Shennongjia 442421, China
泥炭地总面积仅相当于陆地面积的3%, 但其碳储量约为全球土壤碳储量的三分之一, 是最大的陆地土壤碳汇[1—2]。而泥炭藓属物种是泥炭地中重要的造碳植物[3]。泥炭藓湿地是以泥炭藓属植物占优势的沼泽湿地, 其在固碳[4]、维持生物多样性[5]、净化污染物[6—7]等方面具有重要的生态价值, 而且泥炭藓湿地通常发育于江河源头地区[8—10], 具有涵养水源、水文调节功能[11—12], 因此对泥炭藓湿地进行保护和研究尤为重要。
种群的内分布格局指其个体在生活空间的相对位置, 包括均匀型、随机型、集群型[13—14]。泥炭沼泽中泥炭藓种群在不同群落内呈现明显的聚集型分布[15]。大量研究表明, 泥炭沼泽的种群分布格局与四个主要梯度有关:地下水埋深梯度、养分有效性梯度、酸碱度梯度、沼泽边缘—沼泽扩展梯度[16—18]。当地下水埋深、养分、酸碱度任一梯度具有明显变化时, 物种种群分布也具有明显变化[17, 19]。而关于沼泽边缘—沼泽扩展梯度, 有学者认为其背后的生态原因没有明确界定, 具有复杂性, 并且因地而异[16, 20]。
大九湖泥炭藓湿地是华中地区保存最完好的亚热带亚高山泥炭沼泽湿地[21]。大九湖湿地包含九个人工湖泊, 由南向北依次排开, 并由沟渠串联在一起[22], 靠近其中几个湖泊边缘处存在若干发育良好的泥炭藓沼泽湿地斑块。前期我们在大九湖泥炭藓湿地植被调查中发现, 沿垂直湖岸带向外距离的增加, 维管植物群落类型由草本群落(苔草群落→藜芦群落)到灌木群落(绣线菊群落)发生明显变化。与此同时, 大九湖泥炭藓湿地关键物种—大泥炭藓(Sphagnum palustre L.)种群的生长和分布也发生着变化, 这可能会进一步影响到湿地的演替以及湿地生态功能的发挥, 这也显示对大泥炭藓种群分布特征及其制约因素进行研究具有重要意义。早期, Green研究了毛壁泥炭藓(Sphagnum imbricatum)种群的空间分布特征[20]。就国内而言, 仅陈旭对长白山哈尼泥炭地多种苔藓植物分布与环境因子的关系进行研究[23], 并未涉及大泥炭藓, 而该研究区域的自然环境及植物群落类型与亚热带区域差异也较大。本文以亚热带山地大九湖泥炭藓湿地为研究区域, 通过设置样带和样方进行植被和环境因子的调查、取样和测定分析, 探讨大泥炭藓(S. palustre)种群的分布特征及其制约因素, 为亚热带山地泥炭藓湿地的保护和恢复提供理论支撑。
1 材料与方法 1.1 研究区概况神农架大九湖湿地发育于大巴山东段的岩溶盆地之中, 海拔1730—1800m, 年均温7.4℃, 全年日照1000h左右, 年降水丰富, 可达1528mm, 相对湿度大于80%[21]。泥炭藓沼泽是大九湖湿地的重要组成部分, 本文以二号湖(31°28′N, 110°01′E, 海拔1759m)西南侧泥炭藓沼泽为研究区, 前期调查显示, 该区域植物群落的分布和生长状况沿垂直湖岸带方向具有明显的变化规律, 即从湖边向外依次为:拂子茅-大泥炭藓群丛(Ass. Calamagrostis epigeios-Sphagnum palustre)、丝叶苔草-大泥炭藓群丛(Ass. Carex capilliformis-Sphagnum palustre)、湖北大戟+丝叶苔草-大泥炭藓群丛(Ass. Euphorbia hylonoma+Carex capilliformis-Sphagnum palustre)、分株紫萁-大泥炭藓群丛(Ass. Osmunda cinnamomea-Sphagnum palustre)、丛毛羊胡子草-大泥炭藓群丛(Ass. Eriophorum comosum-Sphagnum palustre)、藜芦-大泥炭藓群丛(Ass. Veratrum nigrum-Sphagnum palustre)、拂子茅+地榆-大泥炭藓群丛(Ass. Calamagrostis epigeios+Sanguisorba officinalis-Sphagnum palustre)、地榆+丝叶苔草-大泥炭藓群丛(Ass. Sanguisorba officinalis+Carex capilliformis-Sphagnum palustre)、红穗苔草+地榆-大泥炭藓群丛(Ass. Carex argyi+Sanguisorba officinalis-Sphagnum palustre)、地榆+丝叶苔草群丛(Ass. Sanguisorba officinalis+Carex capilliformis)、绣线菊-地榆群丛(Ass. Spiraea salicifolia-Sanguisorba officinalis)。
1.2 实验方法 1.2.1 野外调查与样品采集2020年8月, 在对大九湖泥炭藓湿地植被进行野外调查的基础上, 于二号湖西南侧泥炭藓沼泽区选取调查样地(见图 1)。根据群落外貌特征, 在样地内沿垂直于湖岸带方向设置3条5m宽样带, 在样带内依据优势种选取11种植物群丛类型, 每类型设置3个1m×1m调查样方(编号:1—33号)。用手持GPS定位仪(UniStrong MG711北京)测量样方中心点到湖岸线的垂直距离。记录每一样方内出现的所有植物种类及每种植物的株数、株高、盖度等;此外, 选取样方内代表性区域放置10cm×10cm的采样框, 统计框内的大泥炭藓头状枝数量, 随后采集框内全部大泥炭藓植株装于塑封袋密封。共采集泥炭藓样品27份(26—31号样方未生长大泥炭藓)。在调查植物的同时, 每个1m×1m样方使用穿孔PVC管法[24]测量地下水埋深, 使用针管吸取法采集水样[25];用环刀法采集土样, 在上述10cm×10cm采样框内完成全部大泥炭藓植株采集并去除其他维管植物后, 使用环刀采集框内表层土壤。将所有样品带回实验室, 进行相关指标测定。

|
| 图 1 研究区域示意图 Fig. 1 Schematic diagram of study area |
大泥炭藓鲜重于采集后2h内测定, 随后室内阴干再70℃恒温烘干48h, 测其干重。分别将土样和大泥炭藓头状枝干燥后研磨过100目筛, 随后进行各理化指标测定。土样测定指标包括含水量、容重、孔隙度、固相比、液相比、气相比、pH、总氮(TN)、总磷(TP)、总有机碳(TOC)、碱解氮(AHN)、有效磷(AP);大泥炭藓测定指标包括总氮、总磷、总碳(TC)、总叶绿素(a+b)、类胡萝卜素;水样测定指标包括pH、总氮、总磷。具体测定方法见表 1。
| 测定指标 Measurement indicators |
测定方法及仪器型号 Measurement method and instrument model |
| 土样、水样pH Soil pH and water pH | pH电位计测定(pHS-3E型, 上海) |
| 土样 TN Soil TN | 酸溶(H2SO4-HCLO4)-靛酚蓝比色法 |
| 土样、藓样 TP Soil TP and capitula TP in S. palustre | 酸溶(H2SO4-HCLO4)-钼锑抗吸光法 |
| 土样 TOC Soil TOC | 重铬酸钾容量法-外加热法 |
| 土样 AHN Soil AHN | 碱解扩散法 |
| 土样 AP Soil AP | 0.5mol/L NaHCO3浸提法 |
| 水样 TN、TP Water TN and TP | 碱性过硫酸钾消解法 |
| 藓样 TC、TN Capitula TC and TN in S. palustre | Elementar元素分析仪(vario MACRO CUBE, Germany) |
| 藓样总叶绿素、类胡萝卜素 Total chlorophyll and carotenoids in capitula of S. palustre | 乙醇浸提法-分光光度法 |
| TN: 总氮 Total nitrogen content; TP: 总磷 Total phosphorus content; TOC: 总有机碳 Total organic carbon content; AHN: 碱解氮 | |
采用多种回归模型检验垂直湖岸带距离与各环境因子(土壤含水量、容重、孔隙度、固相比、液相比、气相比、pH、TN、TP、TOC、AHN、AP、TOC/TN、TOC/TP、TN/TP, 地下水埋深, 沼泽水的pH、TN、TP、TN/TP)、各生理因子(大泥炭藓TN、TP、TC、TC/TN、TC/TP、TN/TP、总叶绿素、类胡萝卜素)及大泥炭藓三生长指标(头状枝数量、盖度、生物量)之间的相关性, 并筛选出最优模型进行分析。使用多重线性逐步回归分析分别检验影响大泥炭藓生长的主导环境因子和生理指标;随后使用Pearson相关性分析检验环境因子与植物生理指标的相关性。所有数据分析使用软件SPSS 20.0和Excel 2010完成;图形制作使用软件Excel 2010完成。
2 结果与分析 2.1 环境因子对距离的响应回归分析表明除土壤TP、AP、地下水TN/TP与距离无显著相关性外, 其余环境因子与距离均显著相关(图 2)。其中, 气相比沿垂直湖岸带距离的增加持续增加;土壤容重、固相比、pH、TOC/TN沿距离呈倒单峰模式, 最小值出现在距湖岸60m左右;土壤含水量、孔隙度、液相比、TN、TOC、AHN、TOC/TP、TN/TP、地下水埋深、地下水pH、TN、TP沿距离呈单峰模式, 其中土壤液相比在距离40m左右出现最大值, 土壤含水量、孔隙度、TN、TOC、AHN、TOC/TP、TN/TP及地下水pH在距湖岸60m左右出现最大值, 地下水TN、TP在距湖岸80m左右出现最大值。

|
| 图 2 各环境因子与垂直湖岸带距离的回归分析 Fig. 2 Regression analysis between each environmental factor and vertical distance from lakeshore |
回归分析表明, 除大泥炭藓TN、TC/TN、总叶绿素、类胡萝卜素与距离无显著相关性外, 其余生理指标与距离均显著相关(图 3)。其中, TC、TC/TP、TN/TP沿距离呈单峰模式, 最大值出现在距湖岸60m左右;TP沿距离呈倒单峰模式, 最小值出现在距湖岸60m左右。

|
| 图 3 大泥炭藓各生理指标与垂直湖岸带距离的回归分析 Fig. 3 Regression analysis between each physiological indicator of S. palustre and vertical distance from lakeshore |
回归分析结果表明大泥炭藓三生长指标(头状枝数、盖度、生物量)对距离的响应具有一致性, 随着垂直湖岸带距离的增加均呈现先增加后减小的趋势(图 4), 且最大值都出现在距湖岸40m左右。

|
| 图 4 大泥炭藓三生长指标与垂直湖岸带距离的回归分析 Fig. 4 Regression analysis between each of three growth indicators of S. palustre and vertical distance from lakeshore |
多重线性逐步回归分析表明, 在20个环境因子(土壤含水量、容重、孔隙度、固相比、液相比、气相比、pH、TN、TP、TOC、AHN、AP、TOC/TN、TOC/TP、TN/TP;地下水埋深;水样pH、TN、TP、TN/TP)中, 仅地下水埋深(X1)对生物量(Y)影响显著, 且二者正相关(表 2), 说明地下水埋深是影响大泥炭藓生长最主要的环境因子。回归方程如下:
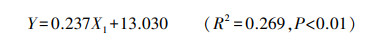
|
| 大泥炭藓TC TC in S. palustre |
土壤TOC TOC in soil |
土壤TOC/TP TOC/TP in soil |
|
| 大泥炭藓 TC TC in S. palustre | 1 | ||
| 土壤 TOC TOC in soil | 0.763** | 1 | |
| 土壤 TOC/TP TOC/TP in soil | 0.501** | 0.902** | 1 |
| ** 分别表示在0.01水平上显著相关 | |||
基于上述分析, 在去除地下水埋深后对剩下19个环境因子进行与生物量的多元线性逐步回归分析, 发现土壤TOC/TP(X2)对生物量(Y)影响显著。回归方程如下:
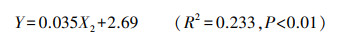
|
因此, 对大泥炭藓生长影响最大的环境因子是地下水埋深, 其次是土壤TOC/TP。
2.5 大泥炭藓生理指标对其生长的影响多重线性逐步回归分析表明, 在大泥炭藓的8个生理指标(TN、TC、TP、TC/TN、TC/TP、TN/TP、总叶绿素、类胡萝卜素)中, 仅TC(X)对生物量(Y)影响显著, 且二者正相关。回归方程如下:
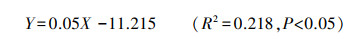
|
基于上述环境因子、生理指标对大泥炭藓生物量影响的分析, 进一步对土壤TOC/TP、土壤TOC与大泥炭藓组织TC进行Pearson相关分析(表 2)。结果显示, 土壤TOC(r=0.763, P<0.01)、土壤TOC/TP(r=0.501, P<0.01)与大泥炭藓组织TC呈极显著正相关。
3 讨论本研究以头状枝数量、盖度和生物量三指标反映大泥炭藓的生长情况, 并以生物量代表大泥炭藓的最终生产力。多重线性回归分析表明, 地下水埋深是影响大泥炭藓生长最主要的环境因子, 而地下水埋深随着垂直湖岸带距离的增加先增加后降低(图 2)。因此, 研究区域大泥炭藓生长随着远离湖岸带方向先增加后降低可能是地下水埋深的相应变化导致的结果。前人大量研究表明, 地下水埋深是影响泥炭藓生长的最重要环境因子[24, 26—27], 并且在一定范围内地下水埋深与生物量显著正相关[28]。本文也证实了上述结论, 在-60—-10cm范围内, 两者显著正相关, 大泥炭藓生物量随地下水埋深的降低持续减小(Y=0.237X1+13.030);并且在地下水埋深低于-40cm时不再生长大泥炭藓, 这与部分学者的研究结果一致[29—30]。由于泥炭藓缺乏真正的根和内部导水组织[26], 对被动水分运输的依赖使其对水分变化十分敏感[27]。当地下水埋深较低时将阻碍毛细水向泥炭藓头状枝(光合作用最活跃)部分的运输, 进一步影响泥炭藓的光合作用和呼吸作用[31], 最终导致生产力的降低[28]。
本研究中, 地下水埋深沿垂直湖岸带距离增加呈先增高后降低趋势, 这可能是受到湿地内微地形影响。在距湖岸60m出现最高地下水埋深-10cm(图 2), 此时大泥炭藓也生长的最好(图 4), 这与Tuittila等[31]和李亭亭等[24]研究发现0—-10cm左右为大泥炭藓生长的最佳水位条件的结论相一致。
除地下水埋深外, 土壤TOC/TP是影响大泥炭藓生物量的第二重要环境因子(Y=0.035X2+2.69)。经分析发现土壤TOC、大泥炭藓组织TC以及大泥炭藓生物量三者之间均显著正相关, 但土壤TP、组织TP与生物量之间均无显著相关性(P>0.05)。这可能表明:主要是土壤中的有机碳能够被大泥炭藓吸收同化到自身组织, 进而对其生长产生影响。因此, 从这个角度, 也可能是随着垂直湖岸带距离的增加带来的土壤TOC先增加后降低的变化(图 2)导致大泥炭藓的生长沿着此方向呈现出先增加后降低的趋势(图 4)。本研究中, 大泥炭藓随着组织TC含量(327.57—429.67mg/g范围内)的增加生长的更好(Y=0.05X-11.215), 显示了C对大泥炭藓的积极作用。这种积极作用在其他泥炭藓物种的研究中已经被发现:Smolders[32]发现组织中的C能使泥炭藓产生坚硬、直立的茎干, 促使其垂直向上生长, 进一步形成了密集紧凑的聚合体, 这种策略创造了有利于自身生长的微环境, 以此来避免减小与其他物种进行竞争。另一方面, 泥炭藓组织中以C为主的碳水化合物能够赋予细胞壁更大的持水能力和强度[33], 且过量的C会转化为萜类、单宁等化合物, 这类化合物使泥炭藓能够更好地抵御食草动物和病原体[33], 均有利于泥炭藓自身的生长。
尽管地下水埋深、大泥炭藓TC和大泥炭藓生物量沿垂直湖岸带距离的变化均呈现较为一致的单峰趋势(图 2, 图 3, 图 4), 但前两者与生物量的峰值却出现少许“错位”。地下水埋深与大泥炭藓TC峰值均出现在距湖岸60m左右(图 2, 图 3), 而大泥炭藓生物量峰值出现在距湖岸40m左右(图 4)。造成这种“错位”的原因可能有两方面:一是本研究中在距湖岸40m左右没有设置样方, 距该处最近的样方设置在距湖岸50m左右, 因此野外实际峰值与模型模拟的峰值存在少许误差;二是可能受到样地内维管植物的影响。本研究着重对非生物因子进行了分析, 但生物因子(如植物的种间作用)对大泥炭藓的生长可能也起着不可忽视的作用。前人多项研究表明, 一定条件下维管植物, 尤其是莎草科植物, 能够通过遮阴或提供结构支撑促进泥炭藓的生长[34—35]。通常维管植物在盖度小于60%左右时对泥炭藓产生有利影响[35], 当盖度过大时则会产生不利影响。本研究中, 在距湖岸50m左右处, 样方优势种为莎草科植物丛毛羊胡子草(Eriophorum comosum), 盖度约60%;而在距湖岸60m左右处, 样方优势种为藜芦(Veratrum nigrum), 盖度约65%。与莎草类植物不同, 藜芦茎粗壮、叶大, 幼时叶片在基部生长紧密, 挤占泥炭藓的生存空间, 而且遮光率较大, 不利于泥炭藓生长。因此, 可能是模型拟合产生的误差以及维管植物对大泥炭藓的种间作用共同导致了大泥炭藓在40m(而非60m)左右生长的最好, 这需要今后进一步试验来验证。
大九湖湿地是北亚热带较为罕见的亚高山沼泽湿地, 特别是其发育的泥炭藓沼泽是在较高海拔山地冷湿气候条件下形成。但大九湖湿地在历史上以排涝除渍为主的开发活动中修建起庞大的人工排水系统, 在大九湖填湖造田、围泽植草的同时, 也重创了大九湖的湿地生态系统。其直接的生态后果是原有的沼泽湿地植被特别是泥炭藓湿地植被退化, 形成较大面积杂草类草甸以及较旱化的金发藓藓丘。随着神农架国家公园的建设, 大九湖泥炭藓湿地的保护和恢复也迫在眉睫。本研究表明, 地下水埋深是影响大泥炭藓生长的至关重要的环境因子, 而-10cm左右的地下水埋深最有利于大泥炭藓种群的生长。该研究结果对重建大九湖湿地水文系统, 恢复大九湖泥炭藓湿地沼泽生态系统具有重要指导意义。
4 结论在前期调查发现大九湖泥炭藓湿地的植被沿垂直湖岸带方向呈现明显的带状分布现象的基础上, 本研究进一步对大泥炭藓种群生长情况及环境因子进行调查采样, 探究大泥炭藓种群生长及分布特征对垂直湖岸带距离的响应及其制约因素。结论如下:
(1) 随着垂直湖岸带距离的增加, 大泥炭藓种群生产力呈现出先增加后降低的趋势, 在垂直湖岸带40m左右生长最好。
(2) 地下水埋深和土壤TOC含量是影响大泥炭藓种群生长及分布特征的主要环境因子;地下水埋深通过影响头状枝光合和呼吸作用, 土壤TOC通过影响大泥炭藓吸收同化到组织中的TC, 来影响其生长。在本研究范围内, 地下水埋深、土壤TOC、组织TC均对大泥炭藓生长产生促进作用。
(3) 除了非生物因子, 生物因子(例如维管植物的种间作用), 也可能是影响泥炭藓种群生长及分布特征的重要因子。
| [1] |
Gorham E. Northern Peatlands: role in the carbon cycle and probable responses to climatic warming. Ecological Applications, 1991, 1(2): 182-195. DOI:10.2307/1941811 |
| [2] |
Binet P, Rouifed S, Jassey V E J, Toussaint M L, Chiapusio G. Experimental climate warming alters the relationship between fungal root symbiosis and Sphagnum litter phenolics in two peatland microhabitats. Soil Biology and Biochemistry, 2017, 105: 153-161. DOI:10.1016/j.soilbio.2016.11.020 |
| [3] |
Clymo R S. Sphagnum-dominated peat bog: a naturally acid ecosystem. Philosophical Transactions of the Royal Society B: Biological Sciences, 1984, 305(1124): 487-499. |
| [4] |
庞毓雯, 黄雨馨, 俞立鹏, 问静怡, 吴玉环, 徐俊锋. 基于MODIS数据的神农架大九湖泥炭藓沼泽植被指数变化研究. 生态学报, 2019, 39(13): 4872-4882. |
| [5] |
卢建利, 吴法清, 郑炜. 湖北二仙岩亚高山泥炭藓沼泽湿地两栖爬行动物资源调查. 四川动物, 2007, 26(2): 374-376. |
| [6] |
胡鸿兴, 何伟, 刘巧玲, 胡万辉, 沈虹, 任晓迪, 石宇鹏, 杨阳, 田艳萍, 雷卫国, 彭博, 张五州, 刘玲, 钟鑫, 陈侈, 秦蓓蓓, 张南南, 毛娜, 江丽, 贾明元, 沈娟, 古南辉, 陈文方. 大九湖泥炭藓湿地对磷、铜污染物净化作用的模拟研究. 长江流域资源与环境, 2008, 17(6): 920-926. |
| [7] |
胡鸿兴, 张岩岩, 何伟, 田蓉, 钟鑫, 韩世松, 李思思, 王俊杰, 陈文方, 杨阳, 陈侈, 邓晗, 文英, 崔雅婷, 李茜, 王璇, 彭菁菁, 高鑫, 唐义. 神农架大九湖泥炭藓沼泽湿地对镉(Ⅱ)、铜(Ⅱ)、铅(Ⅱ)、锌(Ⅱ)的净化模拟. 长江流域资源与环境, 2009, 18(11): 1050-1057. |
| [8] |
汪正祥, 雷耘, 刘胜祥, 方元平, 满金山, 彭宗林, 张柳, 马广礼. 湖北七姊妹山自然保护区发现亚高山泥炭藓湿地. 华中师范大学学报: 自然科学版, 2005, 39(3): 387-388. |
| [9] |
Li T T, Lei Y, Dai C, Yang L F, Li Z Q, Wang Z X. Effects of both substrate and nitrogen and phosphorus fertilizer on Sphagnum palustre growth in subtropical high-mountain regions and implications for peatland recovery. Wetlands Ecology and Management, 2018, 26(4): 651-663. |
| [10] |
Li T T, Liu T, Lei Y, Li Z Q, Dai C, Wang Z X. Effects of the substrate and planting method on Sphagnum palustre growth in subtropical high-mountain regions and the underlying mechanisms. Wetlands, 2019, 39(4): 879-893. |
| [11] |
Price J S, Edwards T W D, Yi Y, Whittington P N. Physical and isotopic characterization of evaporation from Sphagnum moss. Journal of Hydrology, 2009, 369(1/2): 175-182. |
| [12] |
黄咸雨, 张志麒, 王红梅, 陈旭, 朱宗敏, 顾延生, 秦养民, 刘金铃, 汪迎春. 神农架大九湖泥炭湿地关键带监测进展. 地球科学, 2017, 42(6): 1026-1038. |
| [13] |
宋永昌. 植被生态学. 上海: 华东师范大学出版社, 2001.
|
| [14] |
伊力塔, 韩海荣, 程小琴, 康峰峰, 张志杰. 灵空山林区辽东栎(Quercus liaotungensis)种群空间分布格局. 生态学报, 2008, 28(7): 3254-3261. |
| [15] |
卜兆君. 东北贫营养泥炭沼泽几种植物的种群生态学研究[D]. 长春: 东北师范大学, 2004.
|
| [16] |
Pellerin S, Lagneau L A, Lavoie M, Larocque M. Environmental factors explaining the vegetation patterns in a temperate peatland. Comptes Rendus Biologies, 2009, 332(8): 720-731. |
| [17] |
Jeglum J K, He F L. Pattern and vegetation-environment relationships in a boreal forested wetland in northeastern Ontario. Canadian Journal of Botany, 1995, 73(4): 629-637. |
| [18] |
Marini L, Nascimbene J, Scotton M, Klimek S. Hydrochemistry, water table depth and related distribution patterns of vascular plants in a mixed mire. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology, 2008, 142(1): 79-86. |
| [19] |
Vitt D H, Chee W L. The relationships of vegetation to surface water chemistry and peat chemistry in fens of Alberta, Canada. Vegetatio, 1990, 89(2): 87-106. |
| [20] |
Wheeler B D, Proctor M C F. Ecological gradients, subdivisions and terminology of north-west European mires. Journal of Ecology, 2000, 88(2): 187-203. |
| [21] |
杜耘, 蔡述明, 王学雷, 何报寅, 徐贵来, 江明喜, 薛怀平, 肖飞. 神农架大九湖亚高山湿地环境背景与生态恢复. 长江流域资源与环境, 2008, 17(6): 915-919. |
| [22] |
刘林峰, 周先华, 高健, 刘梅群, 潘晓斌, 贺磊, 蔡世耀, 赵以军, 潘超, 王卉君, 张佳敏, 叶有旺, 刘伟. 神农架大九湖湿地浮游植物群落结构特征及营养状态评价. 湖泊科学, 2018, 30(2): 417-430. |
| [23] |
陈旭, 卜兆君, 王升忠, 李鸿凯, 姜丽红. 长白山哈泥泥炭地7种苔藓分布与环境关系研究. 湿地科学, 2008, 6(2): 310-315. |
| [24] |
Li T T, Wang Z X, Bu G J, Lin L Q, Lei Y, Liu C Y, Yang L F, Zheng C L. Effects of microtopography and water table on Sphagnum palustre L. in subtropical high mountains and implications for peatland restoration. Journal of Bryology, 2019, 41(2): 121-134. |
| [25] |
燕红. 泥炭沼泽湿地植被演替规律及植物多样性研究[D]. 长春: 东北师范大学, 2015.
|
| [26] |
van Breemen N. How Sphagnum bogs down other plants. Trends in Ecology & Evolution, 1995, 10(7): 270-275. |
| [27] |
Thompson D K, Waddington J M. Sphagnum under pressure: towards an ecohydrological approach to examining Sphagnum productivity. Ecohydrology, 2008, 1(4): 299-308. |
| [28] |
Kim S, Kim Y, Kim Y, Kim K, Wang S, Kang H, Yoo B. Effects of planting method and nitrogen addition on Sphagnum growth in microcosm wetlands. Paddy and Water Environment, 2014, 12(1): 185-192. |
| [29] |
Grosvernier P, Matthey Y, Buttler A. Growth potential of three Sphagnum species in relation to water table level and peat properties with implications for their restoration in cut-over bogs. Journal of Applied Ecology, 1997, 34(2): 471-483. |
| [30] |
Schouwenaars J M. The impact of water management upon groundwater fluctuations in a disturbed bog relict. Agricultural Water Management, 1988, 14(1/4): 439-449. |
| [31] |
Tuittila E S, Vasander H, Laine J. Sensitivity of C sequestration in reintroduced Sphagnum to water-level variation in a cutaway peatland. Restoration Ecology, 2004, 12(4): 483-493. |
| [32] |
Smolders A J P, Tomassen H B M, Pijnappel H W, Lamers L P M, Roelofs J G M. Substrate-derived CO2 is important in the development of Sphagnum spp. New Phytologist, 2001, 152(2): 325-332. |
| [33] |
Verhoeven J T A, Liefveld W M. The ecological significance of organochemical compounds in Sphagnum. Acta Botanica Neerlandica, 1997, 46(2): 117-130. |
| [34] |
Pouliot R, Rochefort L, Karofeld E, Mercier C. Initiation of Sphagnum moss hummocks in bogs and the presence of vascular plants: is there a link?. Acta Oecologica, 2011, 37(4): 346-354. |
| [35] |
Heijmans M M P D, Klees H, Berendse F. Competition between Sphagnum magellanicum and Eriophorum angustifolium as affected by raised CO2 and increased N deposition. Oikos, 2002, 97(3): 415-425. |