 2023, Vol. 43
2023, Vol. 43文章信息
- 刘启龙, 程赛赛, 陈婷, 常亮, 高梅香
- LIU Qilong, CHENG Saisai, CHEN Ting, CHANG Liang, GAO Meixiang
- 不同林型地表甲虫食性及营养级关系: 基于碳氮稳定同位素分析
- The relationship between the feeding habits and trophic levels of surface beetles in different forest types: Based on carbon and nitrogen stable isotope analysis
- 生态学报. 2023, 43(6): 2242-2252
- Acta Ecologica Sinica. 2023, 43(6): 2242-2252
- http://dx.doi.org/10.5846/stxb202102070398
-
文章历史
- 收稿日期: 2021-02-07
- 网络出版日期: 2022-11-03

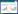




2. 南京农业大学资源与环境科学学院, 土壤生态实验室, 南京 210095;
3. 中国科学院东北地理与农业生态研究所, 长春 130301;
4. 宁波大学地理与空间信息技术系, 宁波 315211;
5. 农产品质量安全危害因子与风险防控国家重点实验室, 宁波 315211
2. Soil Ecology Laboratory, College of Resources and Environmental Sciences, Nanjing Agricultural University, Nanjing 210095, China;
3. Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130301, China;
4. Department of Geography and Spatial Information Techniques, Ningbo University, Ningbo 315211, China;
5. State Key Laboratory for Managing Biotic and Chemical Threats to the Quality and Safety of Agro-Products, Ningbo 315211, China
土壤动物作为土壤生态系统循环的重要组成部分, 因其种类繁多、食性复杂等特点, 对生态系统过程中物质和能量的流动起着十分重要的作用[1—3]。因此通过了解土壤动物的食性、营养关系和它们之间的作用机制对于理解物种间相互作用的机理是十分重要的[4]。传统研究营养关系的方法是通过对消化道内的食物成分进行分析来对食物进行确定, 但因为不同的食物类型被消化吸收的速度不同, 而且对于土壤动物这些体型较小的动物很难对他们进行胃含物的分析[5—8]。因此本文选取稳定同位素分析技术分析地表甲虫的营养级及食性。稳定碳同位素可以用来追踪碳源, 理解生物圈各组成部分之间的运动或重建过去的生态环境条件[9—10], 并通过追踪动物特定食物中稳定的氮同位素丰度能提供有关动物营养水平的信息[1]。
稳定同位素技术已经成为研究生态系统食物网结构和营养关系的重要手段, 近年来已被广泛应用在生态学研究领域[11]。碳、氮稳定同位素通常随着营养等级升高而富集, 并且每个营养级间具有相对恒定(1.5‰—3.0‰)的富集效应, 通过测定生物体内13C值能够准确的分析出不同生物的食物来源、15N值能够分析出营养级位置[12]。国内外对土壤动物的研究包括物种的空间格局[13]、群落结构[14]及驱动因素[15]等方面。目前对生物营养级的研究主要集中在潮间带、洞穴等的水生生物和陆生动物上[16—17], 对土壤动物的研究为大型土壤动物在人工防护林中的营养级划分及土壤群落的营养结构方面[18], 稳定同位素分析方法在了解各种生态系统土壤动物群落的营养结构和营养联系方面取得了重大进展[19]。Anton等利用稳定同位素分析方法研究了土壤群落的营养结构, 阐明了土壤食物网的基础食物资源, 揭示了地上和地下生态系统间的联系[20]。而对土壤动物中地表甲虫的研究多集中在单科甲虫的幼虫摄食习性等方面[21], 对不同林型地表甲虫的食性和营养级研究较少。由于土壤动物的食性特征和营养等级是表征生态系统食物链的重要依据, 因此本文利用稳定同位素技术分析小兴安岭凉水自然保护区地表甲虫不同林型不同物种的13C、15N同位素、营养级和食性特征, 为森林生态系统食物链研究提供基础。
1 材料与方法 1.1 研究区概况研究区位于黑龙江省伊春市带岭区凉水自然保护区内(图 1), 坐标为47°10′50″N, 128 °53′20″E。该区地处欧亚大陆东缘, 小兴安岭山脉南坡达里带岭支脉的东坡, 海拔高度在280—707m之间。为典型的低山丘陵地貌。属温带大陆性夏雨季风气候, 夏季短, 多雨水, 降雨占全年60%以上。冬季长且寒冷干燥。年平均降水量680 mm左右, 年平均气温-0.3 ℃。地带性土壤为暗棕壤, 非地带性土壤为草甸土、沼泽土和泥炭土。境内森林覆盖率达96%, 具有古老的区系和群落发生的历史, 保存了较完整而典型的地带性顶极群落类型及自然景观。区内地带性植被是以红松(Pinus korarensis)为主的温带针阔混交林, 伴生有冷杉( Abies nephrolepis)、红皮云杉(Picea koraiensis)、鱼鳞云杉(Picea jezoensis)等针叶树种。本文选取2种原始林: 原始阔叶红松林(KY)和谷地云冷杉林(YL)、2种次生林: 阔叶红松择伐林(ZF)和次生白桦林(BH)、2种人工林: 落叶松人工林(RL) 和红松人工林(RHS), 共6种林型作为实验样地。

|
| 图 1 研究区概况图 Fig. 1 Overview of the study area |
于2017年6月和7月在6种不同林型内分别设置3个20m×30m的重复样方, 3个样方随机分布且彼此间隔50m以上。采用陷阱法采集地表甲虫, 在每个样方的4个顶点中随机选择3个顶点(以白色PVC管作为地标), 在以地标为中心的30cm半径圆内选取一个采样点。在每个采样点用土钻(内径7cm)挖取一个深15cm的柱状土坑, 将诱捕杯(内径7cm, 高度14cm)置于土坑内使杯口与地面齐平, 内置约占诱捕杯容积2/3的饱和NaCl溶液, 杯口上方约10cm处支起一个一次性餐盘防止凋落物和雨水等杂物进入, 陷阱放置野外7天7夜后取回。手捡法分拣地表鞘翅目成虫, 并置于95%医用酒精中保存, 甲虫成虫在体视显微镜(MoticSMZ168)下观察, 参照《原色中国东北土壤甲虫图鉴—步行虫类》[22]、《原色中国东北土壤甲虫图鉴—隐翅虫类拟步甲类》[23]、《中国东北的葬甲科研究》[24]、《中国土壤动物检索图鉴》[25]将其鉴定到种。
1.3 碳氮稳定同位素的测定将6月和7月采集的不同林型的部分物种样本(根据实际采集情况, 每个物种采集1—5只不等, 取其均值)的稳定同位素含量由MAT253气体稳定同位素质谱仪(美国T核膜Fisher公司)测定。碳、氮稳定同位素比值用国际通用的δ值表示[20], 计算公式如下:
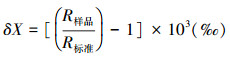
|
(1) |
式中, X代表13C或15N, R为重同位素和轻同位素的比值(13C/12C、15N/14N)。其中碳稳定同位素的标准物质为VPDB(Vienna Pee Dee Belemnite), 氮稳定同位素的标准物质为空气中的N2。δ13C值分析精度 < 0.2‰, δ15N值分析精度 < 0.4‰。
1.4 营养级计算公式营养等级的计算公式为:
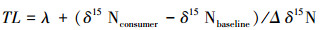
|
(2) |
式中, δ15Nbaseline为生态系统食物网的初级生产者或初级消费者的氮稳定同位素比率, λ=1时, baseline为初级生产者, λ=2时, baseline为初级消费者, Δδ15N为营养等级富集度[26]。
基线生物的正确选取是保证生物营养级计算准确的关键因素[27]。在不同林型中, 地表甲虫δ15N最低值分别为原始阔叶红松林中的Curculionidae sp., 阔叶红松择伐林中的Philonthus japonicas sp., 次生白桦林中的Philonthus parcus sp., 落叶松人工林中的Caficus nudus sp., 红松人工林中的Dicranoncus femoralis sp.和谷地云冷杉林中的ocypus lewisius sp.。因此选取以上地表甲虫作为基准生物[12]。参考国外学者对δ15N在相邻营养级的富集量的研究, 选择3.4‰作为富集量进行分析[28]。
1.5 数据处理运用SPSS22.0中的Shapiro-Wilk检验方法对6种不同林型内地表甲虫碳氮稳定同位素值进行正态分布检验, 数据符合正态分布。运用单因素ANOVA分别对6月(样本量为138只)、7月(样本量为169只)不同林型碳氮稳定同位素值进行分析, 并对6种林型间营养级差异进行分析(样本量为307只), 均通过最小显著性差异(LSD) 法对其进行多重比较。并对不同科地表甲虫进行聚类分析, 分析食性特征。
2 结果 2.1 碳氮稳定同位素值 2.1.1 不同林型间地表甲虫碳氮稳定同位素差异分析于2017年6月和7月在凉水自然保护区样地内共捕获10科31种地表甲虫成虫(群落结构特征详见参考文献[29], 6、7月份不同林型地表甲虫的δ13C值差异显著(P < 0.05) (图 2), 其中原始阔叶红松林和次生白桦林显著高于阔叶红松择伐林, 原始阔叶红松林、次生白桦林、红松人工林和谷地云冷杉林显著高于落叶松人工林。不同林型地表甲虫的δ15N值差异显著(P < 0.05), 其中阔叶红松择伐林显著高于其他5种林型。

|
| 图 2 不同林型地表甲虫δ13C、δ15N值 Fig. 2 Values of δ13C、δ15N of different forest type surface beetles KY, BH, ZF, RHS, YL, RL分别为原始阔叶红松林, 次生白桦林, 阔叶红松择伐林, 红松人工林, 谷地云冷杉林, 落叶松人工林; 图中小写字母顺序表示该组数据平均数由大到小的排列, 不同小写字母指不同林型之间地表甲虫δ13C、δ15N值差异显著(P < 0.05) |
保护区内6月份地表甲虫的δ13C和δ15N值范围分别为-30.45‰至-22.57‰和-1.43‰至5.73‰, 平均值分别为-27.47‰和1.64‰。δ13C值最大的为隐翅虫科, 最小的为步甲科。δ15N值最大值和最小值均为隐翅虫科。6月份不同林型地表甲虫的δ13C值差异显著(P < 0.05)(图 3), 其中原始阔叶红松林和次生白桦林显著高于阔叶红松择伐林, 原始阔叶红松林、次生白桦林和红松人工林显著高于落叶松人工林; 不同林型地表甲虫的δ15N值差异显著(P < 0.05), 其中阔叶红松择伐林显著高于原始阔叶红松林、次生白桦林、红松人工林和落叶松人工林。7月份地表甲虫δ13C值范围为-34.73‰至-23.73‰, 最大的为隐翅虫科, 最小的为叶甲科, 均值为-27.88‰。δ15N值变化范围为-4.1‰至8.38‰, 最大值为葬甲科, 最小值为叶甲科, 均值为2.07‰。7月份不同林型地表甲虫的δ13C值差异显著(P < 0.05)(图 3), 其中原始阔叶红松林和阔叶红松择伐林显著高于红松人工林和谷地云冷杉林; 不同林型地表甲虫的δ15N值差异显著(P < 0.05), 其中次生白桦林和谷地云冷杉林显著高于原始阔叶红松林和落叶松人工林。

|
| 图 3 不同林型内地表甲虫6月和7月δ13C、δ15N值 Fig. 3 Values of δ13C、δ15N in June and July of different forest types KY, BH, ZF, RHS, YL, RL分别为原始阔叶红松林, 次生白桦林, 阔叶红松择伐林, 红松人工林, 谷地云冷杉林, 落叶松人工林; 图中小写字母顺序表示该组数据平均数由大到小的排列, 不同小写字母指6月和7月不同林型之间地表甲虫δ13C、δ15N值差异显著(P < 0.05) |
6种林型内7月份较6月份物种多, 其中葬甲科、步甲科和隐翅虫科较其它科物种多(图 4)。6、7月份均出现的物种为步甲科的Pterostichus maoershanensis和Pterostichus adstrictus, 以上两种物种的δ13C值在阔叶红松择伐林中和δ15N值在次生白桦林中相差较大。原始阔叶红松林中阎甲科的Histeridae sp.、阔叶红松择伐林中叶甲科的Chrysomelidae sp. δ13C、δ15N值均最大。次生白桦林中步甲科的Carabus billbergi和Aulonocarabus canaliculatus 6月份和7月份δ13C值相差较大。

|
| 图 4 林型内不同物种地表甲虫δ13C和δ15N值 Fig. 4 δ13C and δ15N values of surface beetles of different species in forest type KY, BH, ZF, RHS, YL, RL分别为原始阔叶红松林, 次生白桦林, 阔叶红松择伐林, 红松人工林, 谷地云冷杉林, 落叶松人工林; Patr, N.ten等为地表甲虫物种代码 |
利用地表甲虫δ15N值对保护区内夏季6种不同林型地表甲虫的营养级进行划分(图 5), 结果显示保护区样地内地表甲虫营养级差异较大, 处于2—4.5营养级之间, 其中阔叶红松择伐林跨度最大(2—4.27), 其次为谷地云冷杉林(2—4.22), 跨度最小的为次生白桦林(2—3.31)。

|
| 图 5 不同林型地表甲虫营养级划分 Fig. 5 Dividing the trophic levels of surface beetles in different forest types |
不同林型地表甲虫的营养级差异显著(P < 0.05)(图 6), 其中次生白桦林分别和阔叶红松择伐林、红松人工林、落叶松人工林、谷地云冷杉林差异显著, 落叶松人工林分别和原始阔叶红松林、阔叶红松择伐林、红松人工林差异显著。原始阔叶红松林、次生白桦林、落叶松人工林和红松人工林营养层次的长度为2级, 阔叶红松择伐林和谷地云冷杉林营养层次的长度为2.5级。

|
| 图 6 不同林型地表甲虫营养级差异性比较 Fig. 6 Comparison of the difference of trophic levels of surface beetles in different forest types KY, BH, ZF, RHS, YL, RL分别为原始阔叶红松林, 次生白桦林, 阔叶红松择伐林, 红松人工林, 谷地云冷杉林, 落叶松人工林; 图中小写字母顺序表示该组数据平均数由大到小的排列, 不同小写字母指不同林型之间地表甲虫营养级差异显著(P < 0.05) |
原始阔叶红松林和次生白桦林中阎甲科的Histeridae sp.营养级最高(图 7)。阔叶红松择伐林和落叶松人工林中葬甲科的Nicrophorus vespilloides营养级最高。落叶松人工林和红松人工林中隐翅虫科的Philonthus wuesthoffi和Philonthus japonicus营养级最高。6种林型中营养级最高的物种均出现在7月份。不同林型中6月份和7月份营养级差异较大的为红松人工林中隐翅虫科的Philonthus japonicas, 相差1.28个营养级, 其它林型中物种营养级在两个月份间相差较小。

|
| 图 7 林型内不同物种地表甲虫营养级 Fig. 7 Vegetative grade of surface beetles of different species in forest type KY, BH, ZF, RHS, YL, RL分别为原始阔叶红松林, 次生白桦林, 阔叶红松择伐林, 红松人工林, 谷地云冷杉林, 落叶松人工林 |
保护区内地表甲虫碳氮同位素比值差异显著(P < 0.05)(图 8), 表明甲虫食源组成复杂, 碳氮比值范围跨度较大。不同科地表甲虫营养级差异显著(P < 0.05)(图 9), 其中葬甲科营养级长度最长, 为2.5, 其次为隐翅虫科和步甲科, 营养级长度分别为2.46和1.98。聚类分析结果表明保护区样地内地表甲虫可分为两个营养群组(图 10): 第一群组为杂食偏肉食性甲虫(隐翅虫科、葬甲科、步甲科和长朽木甲科)和植食性甲虫(叶甲科和阎甲科), 第二组由植食性(象甲科)、肉食性(拟虎甲科)和杂食偏植食性(叩甲科和金龟科)组成。

|
| 图 8 地表甲虫碳氮同位素比值 Fig. 8 Carbon and nitrogen isotope ratios of surface beetles KY, BH, ZF, RHS, YL, RL分别为原始阔叶红松林, 次生白桦林, 阔叶红松择伐林, 红松人工林, 谷地云冷杉林, 落叶松人工林; 图中小写字母顺序表示该组数据平均数由大到小的排列, 不同小写字母指不同林型之间地表甲虫碳氮同位素比值差异显著(P < 0.05) |

|
| 图 9 不同科地表甲虫营养级划分 Fig. 9 Classification of trophic levels of surface beetles in different families |

|
| 图 10 地表甲虫聚类分析 Fig. 10 Surface beetle cluster analysis |
大多数高级动物类群的营养生态位至少跨越两个营养级, 这意味着资源的使用范围很广[20], 这与本文的研究结果吻合, 凉水自然保护区地表甲虫生物营养级范围较广, 处于2—4.5营养级之间, 表现出从植食性到肉食性的广泛食性特征。由于阔叶红松择伐林是在原始红松林样地内经过择伐后形成的林型, 择伐这种干扰增加了植物多样性和土壤动物多性, 进而增加了土壤动物之间关系的复杂性, 导致其营养级跨度比原始阔叶红松林要高。葬甲科的Nicrophorus tenuipes在原始阔叶红松林营养级为2.5, 在谷地云冷杉林为4, 在这两种林型中营养级变化较大, 表明林型对藏甲的营养级有影响。步甲科的Dicranoncus femoralis在红松人工林的营养级为0.7, 与其它林型的物种营养级相差较大, 说明取食特性发生了很大的变化。次生白桦林营养级跨度最小, 说明次生林和人工林内甲虫的营养级数低于原始林。隐翅虫科内不同林型内部营养级跨度较高, 这可能与隐翅虫科物种较丰富有关, 还有可能是受甲虫的自身饮食习性有关。
通过分析生物体的同位素特征可以了解食物网中的营养关系[30]。尽管甲虫具有普遍的形态相似性, 但它们表现出广泛的不同的营养模式: 植食、腐食、捕食、尸食和粪食等, 食物来源广泛[31]。地表甲虫具有杂食食性的种类较多, 杂食性是指生物体利用不同的捕食对象或食物类型的能力, 包括营养水平的杂食性, 即取食于一个以上的营养水平。动物稳定的同位素组成一般反映了它们的食物来源, 有研究表明叩甲科表现出从植食到捕食的广泛的营养模式[30], 毛翅目幼虫在发育过程中由食草性向食肉性转变[32]。国外学者利用稳定碳、氮同位素特征研究了叶甲科幼虫(Diabrotica virgifera)的摄食习性特征, 表明生长在不同的植物上的幼虫, 表现出不同的13C值[17]。利用稳定碳氮同位素研究了两种步甲科幼虫(A.congrua, A.chalcites)采用不同饲养方式的食性, 结果表明甲虫的存活率存在差异[33—34]。本文不同林型地表甲虫物种之间生态位重叠面积较大, 表明地表甲虫食源复杂。保护区内δ13C值和δ15N值跨度7月份均大于6月份, 原因是7月份植物和动物物种都较6月份多。葬甲科是食尸性土壤动物, 6月和7月δ13C值在阔叶红松择伐林最大, 其它林型δ13C值相似; 6月份δ15N值没有明显变化, 7月份6种林型δ15N值均有显著提升。说明δ13C值受原始阔叶红松林择伐后影响较大。由于步甲主要是捕食者, 不同林型土壤动物组成不一样, 也就是步甲捕食的动物(食物来源)不一样, 这样可能会导致这种结果。说明在不同林型中, 由于潜在的食物资源不一样, 步甲捕食的动物不一样, 就会导致δ15N的这种显著差异。因此在调查土壤动物食性关系时, 需要考虑林型之间的差异, 不同林型和生境土壤动物的δ15N会有很大的差别。葬甲科部分物种在不同林型间营养级跨度较大, 而藏甲科主要是食尸性土壤动物, 因此很可能是所食肉食的差异。隐翅虫科在不同林型内部跨度较大, 而隐翅虫科主要是杂食性土壤动物, 表现出所食食物的复杂性。
4 结论(1) 6、7月份不同林型地表甲虫的δ13C、δ15N值差异显著(P < 0.05), 总体来看阔叶红松林和次生白桦林显著高于其它林型, 其中δ13C值受原始阔叶红松林择伐后影响较大。6、7月份均出现的步甲科的Pterostichus maoershanensis和Pterostichus adstrictus δ13C值在阔叶红松择伐林中相差较大、δ15N值在次生白桦林中相差较大。
(2) 6种不同林型地表甲虫营养级处于2—4.5营养级之间, 其中阔叶红松择伐林跨度最大(2—4.27), 其次为谷地云冷杉林(2—4.22), 跨度最小的为次生白桦林(2—3.31)。6种林型中营养级范围较小的象甲科和步甲科营养位置较低, 营养级范围较大的隐翅虫科和藏甲科营养位置较高。6种林型中营养级最高的物种均出现在7月份。保护区内地表甲虫食源组成复杂, 可分为两个营养群组: 第一群组由杂食偏肉食性甲虫和植食性甲虫组成, 第二组由植食性、肉食性和杂食偏植食性组成。
| [1] |
Post D M. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology, 2002, 83(3): 703-718. DOI:10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2 |
| [2] |
Tillberg C V, McCarthy D P, Dolezal A G, Suarez A V. Measuring the trophic ecology of ants using stable isotopes. Insectes Sociaux, 2006, 53(1): 65-69. DOI:10.1007/s00040-005-0836-7 |
| [3] |
Zalewski M, Dudek D, Godeau J F, Maruszkiewicz M. Stable isotopic research on ground beetles. Review of methods. Baltic Journal of Coleopterology, 2012, 12(1): 91-98. |
| [4] |
Juen A, Traugott M. Revealing species-specific trophic links in soil food webs: Molecular identification of scarab predators. Molecular Ecology, 2007, 16(7): 1545-1557. DOI:10.1111/j.1365-294X.2007.03238.x |
| [5] |
杨纪明. 渤海无脊椎动物的食性和营养级研究. 渔业信息与战略, 2001, 16(9): 8-16. DOI:10.3969/j.issn.1004-8340.2001.09.002 |
| [6] |
颜云榕, 卢伙胜, 金显仕. 海洋鱼类摄食生态与食物网研究进展. 水产学报, 2011, 35(1): 145-153. |
| [7] |
窦永静, 常亮, 吴东辉. 土壤动物食物网研究方法. 生态学杂志, 2015, 34(1): 247-255. DOI:10.13292/j.1000-4890.2015.0035 |
| [8] |
李斌, 陈发军, 陶敏, 岳兴建, 王志坚, 张耀光. 夏季若尔盖高寒湿地水生生物群落食物网结构特征. 水生生物学报, 2016, 40(2): 313-320. |
| [9] |
Staddon P L. Carbon isotopes in functional soil ecology. Trends in Ecology & Evolution, 2004, 19(3): 148-154. |
| [10] |
West J B, Bowen G J, Cerling T E, Ehleringer J R. Stable isotopes as one of nature's ecological recorders. Trends in Ecology & Evolution, 2006, 21(7): 408-414. |
| [11] |
Ribera I, Foster G N, Downie I S, McCracken D I, Abernethy V J. A comparative study of the morphology and life traits of Scottish ground beetles (Coleoptera, Carabidae). Annales Zoologici Fennici, 1999, 36(1): 21-37. |
| [12] |
Potapov A M, Semenina E E, Kurakov A V, Tiunov A V. Large 13C/12C and small 15N/14N isotope fractionation in an experimental detrital foodweb (litter-fungi-collembolans). Ecological Research, 2013, 28(6): 1069-1079. DOI:10.1007/s11284-013-1088-z |
| [13] |
Gao M X, Cheng S S, Ni J P, Lin L, Lu T Y, Wu D H. Negative spatial and coexistence patterns and species associations are uncommon for carrion beetles (Coleoptera: Silphidae) at a small scale. European Journal of Soil Biology, 2017, 83: 52-57. DOI:10.1016/j.ejsobi.2017.10.003 |
| [14] |
Chang L, Song X Y, Wang B F, Wu D H, Reddy G V P. Effect of Bt Corn (Bt 38) Cultivation on Community Structure of Collembola. Annals of the Entomological Society of America, 2020, 113(1): 1-5. DOI:10.1093/aesa/saz038 |
| [15] |
Guo Y X, Gao M X, Liu J, Zaitsev A S, Wu D H. Disentangling the drivers of ground-dwelling macro-arthropod metacommunity structure at two different spatial scales. Soil Biology and Biochemistry, 2019, 130: 55-62. DOI:10.1016/j.soilbio.2018.12.002 |
| [16] |
宋博, 陈琳琳, 闫朗, 姜少玉, 刘春云, 李秉钧, 李宝泉. 山东东营和烟台潮间带海草床食物网结构特征. 生物多样性, 2019, 27(9): 984-992. |
| [17] |
王庆, 卢志兴, 赵婧文, 陈又清. 互利关系和植物多样性对节肢动物群落不同营养级的影响路径和强度. 生态学报, 2020, 40(1): 51-59. |
| [18] |
张淑花, 张雪萍. 基于δ15N稳定同位素分析的人工防护林大型土壤动物营养级研究. 生态学报, 2014, 34(11): 2892-2899. |
| [19] |
Minagawa M, Wada E. Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochimica et Cosmochimica Acta, 1984, 48(5): 1135-1140. DOI:10.1016/0016-7037(84)90204-7 |
| [20] |
Potapov A M, Tiunov A V, Scheu S. Uncovering trophic positions and food resources of soil animals using bulk natural stable isotope composition. Biological Reviews, 2019, 94(1): 37-59. DOI:10.1111/brv.12434 |
| [21] |
Ivan H, Adamczyk J J Jr, Higdon M L, Clark T L, Ellersieck M R, Hibbard B E. Carbon isotope ratios document that the elytra of western corn rootworm (coleoptera: chrysomelidae) reflects adult versus larval feeding and later instar larvae prefer Bt corn to alternate hosts. Environmental Entomology, 2014, 43(3): 840-848. DOI:10.1603/EN13248 |
| [22] |
李井科, 林琳, 张雪萍. 原色中国东北土壤甲虫图鉴—步行虫类. 哈尔滨: 哈尔滨地图出版社, 2015.
|
| [23] |
李井科, 张利敏, 张雪萍. 原色中国东北土壤甲虫图鉴—隐翅虫类, 拟步甲类. 哈尔滨: 哈尔滨地图出版社, 2015.
|
| [24] |
李景科, 张雪萍, 林琳, Flutsch G, 于强. 中国东北的葬甲科研究. 哈尔滨: 哈尔滨地图出版社, 2011.
|
| [25] |
尹文英, 胡圣豪, 沈韫芬, 宁应之, 孙希达, 吴纪华, 诸葛燕, 张云美, 王敏, 陈建英, 徐成刚, 梁彦龄, 王洪铸, 杨潼, 陈德牛, 张国庆, 宋大祥, 陈军, 梁来荣, 胡成业, 王慧芙, 张崇洲, 匡溥人, 陈国孝, 赵立军, 谢荣栋, 张骏, 刘宪伟, 韩美贞, 毕道英, 肖宁年, 杨大荣. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [26] |
张丹, 闵庆文, 成升魁, 王玉玉, 杨海龙, 何露. 应用碳、氮稳定同位素研究稻田多个物种共存的食物网结构和营养级关系. 生态学报, 2010, 30(24): 6734-6740. |
| [27] |
Hoekman D, Bartrons M, Gratton C. Ecosystem linkages revealed by experimental lake-derived isotope signal in heathland food webs. Oecologia, 2012, 170(3): 735-743. DOI:10.1007/s00442-012-2329-5 |
| [28] |
Vander Zanden M J, Rasmussen J B. Variation in 15N and 13C trophic fractionation: implications for aquatic food web studies. Limnology and Oceanography, 2001, 46(8): 2061-2066. DOI:10.4319/lo.2001.46.8.2061 |
| [29] |
程赛赛, 倪娟平, 高梅香, 李景科. 小兴安岭森林地表鞘翅目成虫对不同干扰梯度的响应. 生态学报, 2018, 38(9): 3097-3109. |
| [30] |
Ouyang F, Cao J, Liu X H, Men X Y, Zhang Y S, Zhao Z H, Ge F. Tracing dietary origins of aphids and the predatory beetle Propylea japonica in agricultural systems using stable isotope analyses. Entomologia Experimentalis et Applicata, 2015, 155(2): 87-94. |
| [31] |
Samoylova E S, Tiunov A V. Flexible trophic position of polyphagous wireworms (Coleoptera, Elateridae): a stable isotope study in the steppe belt of Russia. Applied Soil Ecology, 2017, 121: 74-81. |
| [32] |
Céréghino R. Shift from a herbivorous to a carnivorous diet during the larval development of some Rhyacophila species (Trichoptera). Aquatic Insects, 2002, 24(2): 129-135. |
| [33] |
Sasakawa K. Laboratory studies on larval food habits of two syntopic, related, granivorous ground beetles Amara chalcites and A. congrua (Coleoptera: Carabidae): a comparison with stable isotope analysis. Applied Entomology and Zoology, 2011, 46(4): 511-518. |
| [34] |
Tsurikov S M, Goncharov A A, Tiunov A V. Intra-body variation and ontogenetic changes in the isotopic composition (13C/12C and 15N/14N) of beetles (Coleoptera). Entomological Review, 2015, 95(3): 326-333. |