 2023, Vol. 43
2023, Vol. 43文章信息
- 鲍歆歆, 周伟奇, 郑重, 徐林莉
- BAO Xinxin, ZHOU Weiqi, ZHENG Zhong, XU Linli
- 城市植物挥发性有机化合物排放与臭氧相互作用及其机制
- The interactions and mechanisms between biogenic volatile organic compounds emissions and ozone concentrations in urban areas: A review
- 生态学报. 2023, 43(5): 1749-1762
- Acta Ecologica Sinica. 2023, 43(5): 1749-1762
- http://dx.doi.org/10.5846/stxb202202240431
-
文章历史
- 收稿日期: 2022-02-24
- 网络出版日期: 2022-10-21





2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085;
3. 中国科学院大学, 北京 100049;
4. 中国科学院生态环境研究中心北京城市生态系统研究站, 北京 100085
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China;
4. Beijing Urban Ecosystem Research Station, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
我国众多城市存在近地面臭氧(O3)浓度超标问题, O3已成为继PM2.5后影响我国空气质量的另一种重要二次污染物, 在部分城市已经成为首要污染物[1—2]。近地面O3作为一种强氧化剂不仅会影响农业生产, 对人体健康也会产生较大的危害[3], O3浓度增加会导致过早死亡人数增加[4]。O3的生成与其前体物挥发性有机化合物(VOCs)、氮氧化物(NOx)浓度及其比例密切相关[1, 5]。近年来我国NOx排放控制卓有成效, 而VOCs排放量巨大, 其来源构成复杂, 减排难度较大, 使得VOCs与NOx减排比例失衡, 是造成O3污染加剧的重要因素[6—7]。因此, O3污染调控需注重VOCs和NOx的协同控制, 尤其是加大VOCs的减排力度。
VOCs是易挥发碳氢化合物的总称, 主要释放源包括人为源和植物源。植物源挥发性有机化合物(BVOCs)是指植物器官(叶、花、果实等)通过次生代谢途径合成的低沸点、挥发性强的不同种类碳氢化合物总称[8—9]。据统计, 全球植物释放的VOCs种类有30000多种[10], 年排放量约占全球挥发性有机化合物年排放总量的90%, 远超人为源挥发性有机化合物(AVOCs)的年排放量[11]。因此, BVOCs对O3形成的贡献不容忽视[12]。
BVOCs对O3形成的贡献, 在城市地区受到高度关注。虽然在城市地区BVOCs的排放低于AVOCs[13], 但BVOCs的化学活性较高, 可以被羟基氧化并影响大气的氧化能力[14]。受城市复杂环境影响, 异戊二烯在城市中产生O3的效率是在农村地区的两倍, 随着NOx排放的逐年减少, 城市建成区内部绿地排放的异戊二烯对城市地区O3形成的贡献超过来源于非城市地区传输的异戊二烯[7]。此外, 城市中促进植物释放BVOCs的环境条件, 比如城市高温, 也会促进O3的生成[15], 从而加剧O3污染。例如, 受城市热岛效应影响, 市中心较高的空气温度会导致更高水平的BVOCs释放, 进而导致O3浓度升高, 而随着全球气候变暖, 这种效应会进一步增强[16]。热浪期间, O3浓度经常会升高, 达到或超过世卫组织设定的O3暴露阈值[16—19]。近年来, 随着城市居民对人居环境改善需求的增加, 城市增大了生态建设的力度, 城市绿色空间增长显著, 城市绿地的增长速度甚至超过了城市人口的增长速度[20]。随着城市绿地的增加, 城市BVOCs的排放也呈现逐年增加的趋势[21—22], 有可能对城市中O3的生成带来更大的影响。尤其是, 城市绿化过程中不合理的植物选择、配置可能会进一步增加BVOCs的排放[23], 从而加剧O3污染[7]。
BVOCs与O3之间具有复杂的相互作用关系。BVOCs是近地面O3形成的重要前体物, 对O3的生成有较大的贡献。其中占比较高的异戊二烯类与单萜类化合物具有较高的化学反应活性, 能与O3反应, 影响大气的氧化性质。已有研究显示, 植物释放的BVOCs对近地面O3生成的贡献不容忽视。与此同时, 植物本身具有一定的吸收O3的能力, 可以通过气孔和非气孔机制吸收O3以改善环境空气质量。BVOCs通过叶子细胞间隙和植物外部环境空气中的气相化学反应, 在去除O3方面也发挥着关键作用[24]。在O3胁迫下, 不同城市植物的BVOCs排放会如何变化?如何影响O3生成?回答这些问题对于城市树种的优化配置非常重要。围绕上述问题, 本文从植物BVOCs的特性与作用出发, 综述了BVOCs和O3两者之间的相互作用与机制研究的最新进展, 进而分析了相关研究存在的不足和未来亟需加强的研究, 以期为城市建设中的树种选择及减轻城市O3污染提供理论依据和借鉴。

|
| 图 1 BVOCs排放对环境因素的响应及其与O3相互作用 Fig. 1 BVOCs emissions responses to environmental stress and its interaction with O3 BVOCs:植物源挥发性有机化合物Biogenic volatile organic compounds |
根据BVOCs的特性, 将其分为4类[11], 其对植物生理及人体健康均具有重要的作用。此外, BVOCs具有很高的化学反应活性, 对流层大气化学过程起到重要的作用。尤其是占比较高的异戊二烯和单萜烯, 对近地面O3的形成有较高的贡献, 有许多研究对其合成机理及特性进行了探究。
1.1 BVOCs的种类及作用Guenther等[11]学者依据BVOCs的化学结构、活性以及其在大气中的滞留时间等特性将其分为4类:异戊二烯类、单萜类(α-蒎烯、β-蒎烯、柠檬烯等)、其他活性VOC(甲基丁烯醇(MBO)、乙烯醛、有机酸、羰基化合物等)、其他次活性VOC(甲醇、乙醇、丙酮等)。其中异戊二烯类与单萜类化合物占比较高, 且具有很高的化学反应活性, 在对流层大气化学过程中具有重要作用。相较于异戊二烯类和单萜类化合物, 另外两类VOC在大气中的滞留时间更久, 在典型的对流层条件下, 其他活性VOC的寿命小于一天, 而其他次活性VOC的寿命大于一天[11]。BVOCs对植物生理及人体健康均具有重要的作用。
BVOCs对植物的生理生态具有重要的影响。除丙酮、甲醇等作为代谢废物排放到体外, BVOCs是植物体内重要的信号传递物质, 在植物的生长、繁殖、发育、抵御生物胁迫和非生物胁迫等方面具有重要作用[25—26];植物释放的单萜类物质能够防御昆虫及哺乳动物的取食[27], 防御以昆虫为载体的细菌和植物体内潜在的致病菌的侵害[25], 能够引诱害虫天敌和传粉昆虫;异戊二烯能够保护植物免受高温和强光的伤害[28]。
BVOCs对人体健康也具有重要影响。随着对BVOCs生理和生态作用的认识, 人们开始研究BVOCs与人体健康之间的关系[29]。近年来, BVOCs的提取、分离和鉴定等方面取得了突破性进展, BVOCs对人体健康的作用从机理上得到了证实。一方面, BVOCs具有抗菌、抑菌, 消除疲劳、促进安眠, 提神醒脑, 解除紧张状态等作用[30—33]。比如, 蜡菊(Helichrysum araxinum)对葡萄球菌分离株有一定的抑菌活性[34], 而针叶树释放的BVOCs能使人体处于放松状态, 缓解紧张情绪。另一方面, 部分BVOCs对人体有害, 过量吸入可能会引起肺部疾病, 损伤中枢神经系统, 甚至致癌致畸[32]。例如, 实验表明, 珍珠梅鲜花和暴马丁香鲜花释放的BVOCs对小鼠的学习记忆能力、探索性、兴奋性及运动性有抑制作用[33]。
由于BVOCs具有高反应活性, 其在对流层大气化学过程中起到十分重要的作用, 具体包括:(1)增加了近地面O3的浓度[35]。BVOCs是O3形成的前体物之一, 能够与NOx以及O3的光解产物发生光化学反应形成O3, 研究表明, 与AVOCs相比, BVOCs的O3形成潜能约占一半[14], 对近地面O3的形成具有较高的贡献。(2)通过与大气中的O3、羟基自由基等氧化性物质的反应形成次级有机气溶胶(SOA)[36], 同时减小了大气的还原性[28]。SOA的形成影响地球系统的辐射平衡, 且增加大气中云凝结核的浓度[37]。大气还原性的减小, 使甲烷等温室气体滞留时间延长;(3)增加偏远地区的酸沉降[38]。BVOCs在NOx存在下可被氧化形成各种寿命相对较长的有机硝酸盐, 在被分解前可进行远距离运输, 以造成偏远地区的酸沉降及NOx的远距离运输;(4)影响CO形成从而影响全球碳循环[28]。光化学反应是大气中CO的重要来源之一, 而CO是影响羟基自由基浓度的重要微量气体之一, BVOCs参与光化学反应影响CO形成, 从而影响碳循环[38]。
1.2 典型BVOCs的合成机理及特性异戊二烯及单萜烯是主要的两类BVOCs, 其中异戊二烯全球年排放量占全球植物总BVOCs年排放量的近一半, 单萜约占十分之一[11]。除占比较高外, 这两类化合物的反应活性较强, 具有较高的O3生成潜力。有许多研究对两者的合成机理及特性进行探究, 异戊二烯合成主要采用甲羟戊酸(MVA)途径和丙酮酸/磷酸甘油醛(MEP)途径[39—40], 单萜类化合物仅通过甲羟戊酸途径合成[41]。除合成途径不同外, 两者的排放机制也存在差异, 异戊二烯在叶绿体内即时合成即时排放[42], 仅在白天排放, 晚上排放量基本为零, 而部分单萜烯合成后可存储在特殊器官中。因此异戊二烯的释放速率与光合有效辐射(PAR)关系密切, 光照影响腺嘌呤核苷三磷酸(ATP)/二甲基烯丙基二磷酸酯(DMAPP)的合成, 间接控制异戊二烯合成酶的活性, 随着光照的增强, 异戊二烯的释放速率迅速增加[43]。而叶片内单萜合成酶的活性受光强的影响较小, 多数试验表明, 晚上单萜烯的排放量变化不明显[44], 但罗勒烯的释放速率受PAR影响强烈[42]。近期有研究表明部分植物合成单萜类也受光照影响[45]。
温度是影响植物释放异戊二烯与单萜烯的另一重要因素, 温度通过增强合成酶的活性, 升高BVOCs蒸气压, 以及降低扩散路径的阻力, 提高释放速率[41], 当温度升高至一定程度后, 其速率会下降[46—48]。相比于异戊二烯, 温度对单萜烯的影响更大, 两者之间呈较好的指数关系[49]。
异戊二烯和单萜烯是最常见的植物源挥发性有机化合物, 在生产甲基乙烯基酮(MVK)、甲基丙烯醛(MACR)、羟基氢过氧化物(ISOPOOH)、甲醛(HCHO)等主要的光化学氧化剂的过程中起着重要的作用[50]。O3的形成潜力一定程度上取决于VOCs的反应活性, 不同化合物的反应活性不同。异戊二烯是66种VOC中羟基自由基·OH反应活性最高的[51], 其反应活性比是AVOCs苯的22倍[51—52]。当气温超过30℃时, 异戊二烯的·OH反应活性是苯的443倍[51]。在夏季与秋季的白天, 植物源异戊二烯浓度高于人为源异戊二烯, 同时白天残留的异戊二烯为夜间贡献了很大一部分, 残留的大部分是生物源的异戊二烯[53]。在城市环境中, 植物源异戊二烯与人为源异戊二烯协同作用下, 对近地面O3的形成有更高的贡献[14], 因此需要进一步关注BVOCs的释放。
2 影响植物释放BVOCs的主要因素不同植物在不同环境中释放BVOCs的速率与组分不同, 与环境的差异及植物的遗传变异有关[54]。影响植物释放BVOCs的内部因素主要包括:树种、树龄、叶龄、芽期、花期等;环境因素主要包括光照、温度、湿度、氮素、二氧化碳、生物胁迫、O3等。随着O3污染的加剧, 其对植物释放BVOCs的影响受到广泛的关注。
2.1 影响植物释放BVOCs的内部因素不同植物释放BVOCs的速率、组分差别很大。排放BVOCs的树种主要是落叶树种和针叶树种如栎属、杨属、柳属、桉属、云杉属等[55—58]。其中柳属、杨属、栎属的大部分种类异戊二烯的排放高, 而桦属、松属等多为萜烯类化合物释放者[59—60]。部分属不同种植物BVOCs释放速率也存在差异, 通过测量柳属4个不同品种柳树的BVOCs排放, 发现4个品种柳树的异戊二烯、单萜烯、倍半萜的释放速率均存在差异[54]。
单个植物的不同发育阶段也会对BVOCs的释放有一定影响, 相较于成熟阶段, 小麦在衰老阶段的BVOCs总排放量增加了两倍[61]。树龄、叶龄、是否处于花期等因素也影响植物BVOCs的释放。幼苗比成熟植株更脆弱, 因此会释放更多BVOCs以提高存活率, 未成熟叶片的总挥发物浓度高于成熟叶片[62], 且植物幼叶相较成熟叶有更高的BVOCs排放速率, 由于新枝叶较薄的表皮厚度会导致较高的排放速率[63—66]。成年树相较幼树有更高的排放速率, 而排放的BVOCs组分是相似的[54, 65]。是否处于开花期同样会影响植物的BVOCs排放速率, 美国皂荚(Gleditsia triacanthos)、海棠(Malus spectabilis)等植物开花期的总单萜排放速率高于开花后, 通过自然气体和气溶胶排放模型(MEGAN)模拟发现, 春季开花期间花卉的BVOCs排放量约占该研究区总单萜的11%[67]。
2.2 影响植物释放BVOCs的环境因素植物释放BVOCs受到许多环境因素的影响。光照和温度影响大部分植物BVOCs的释放速率[68], 主要通过影响合成酶的活性影响BVOCs的排放, 与植物节律也有一定的关系[69]。增温一般对植物BVOCs的释放具有促进作用, 由于瞬时温度效应, 随着气温升高, 能显著增加植物释放的萜烯类化合物浓度[66], 但如果植物长期受高温胁迫, 可能会影响植物的防御能力[70], 从而降低植物的萜烯类化合物释放。例如, 生长季增温增加了挪威云杉针叶中单萜烯的浓度, 但随着云杉的生长, 单萜烯排放速率下降[71]。光照则主要影响异戊二烯合成酶的活性, 随着光照的增强, 异戊二烯的排放速率显著升高, 除罗勒烯外, 大部分单萜烯的排放受光照的影响较小。
食草[72]、真菌或细菌感染[73—74]等生物胁迫及干旱[75]、重金属[76]等非生物胁迫均会诱导植物释放绿叶挥发物(GLVs), GLVs是构成BVOCs中的重要组成部分, 植物在受到机械损伤[77]和生物胁迫时几乎立即释放GLVs以防御草食动物的啃食和病原体的侵入, 并对细菌和真菌产生直接的毒性作用[78]。减少食草动物的啃食显著降低植物BVOCs的排放量[70]。
此外, 湿度、氮素、二氧化碳等因素均对植物释放BVOCs有一定的影响。不同植物受湿度的影响不同, 部分树种释放速率随着环境湿度增加而逐渐增加[79—80], 但有些树种对环境湿度的变化不敏感, 甚至随着湿度的增加而降低;Caser等[81]研究发现干旱胁迫会调节丹参体内与BVOCs有关基因的表达, 特别是引起倍半萜产量的增加。施氮对植物BVOCs释放的影响结果存在较大不确定性[82], 叶面施氮与土壤施氮对植物BVOCs释放的影响不同[83], 如对于马尾松(Pinus massoniana Lamb.), 土壤施氮能显著增加总BVOCs排放量, 而叶面施氮无显著影响。目前研究表明, 施氮对植物BVOCs的影响具有化合物特异性[49], 已有研究显示叶片氮素浓度与植物叶片异戊二烯释放呈显著正相关[84], 但对于萜烯类物质的影响较为复杂, 大部分研究结果显示氮添加显著降低植物萜烯类物质的释放[85—87]。然而, Carriero等[88]关于苏格兰松(Pinus sylvestris)的研究表明, 氮添加对于萜烯类物质的作用并不一致[89], 氮添加对蒎烯有显著抑制作用, 但能刺激己醛、DMNT((E) -4, 8-二甲基-1, 3, 7-壬三烯)等BVOCs的释放。二氧化碳对BVOCs释放的影响, 目前不同研究结果不同, 存在一些争议, 有部分研究显示高浓度二氧化碳可以促进部分植物BVOCs的排放, 但一些研究表明高浓度二氧化碳熏蒸可以抑制BVOCs的排放, 也有研究显示BVOCs对二氧化碳浓度变化不敏感[50, 90—92]。环境因子除影响叶片BVOCs的释放外, 部分环境因子对根际BVOCs的释放也有影响[71], 例如氮添加、变暖会减少了苏格兰松(Pinus sylvestris)根际BVOCs的排放总量[93];单次暴露的动物啃食会增加根际BVOCs的排放, 但长期胁迫下, 相较往年会减少根际BVOCs的排放[94]。
近年来, 随着近地面O3浓度的增加, 越来越多的研究关注O3浓度增加对植物、环境、人体健康等的影响, O3的峰值浓度不仅会导致叶片损伤和抑制植物生长, 还会影响包括BVOCs在内的次生代谢产物的合成和释放[8]。由于BVOCs对大气光化学过程的重要影响以及O3与BVOCs之间复杂的相互作用关系, O3胁迫对植物BVOCs释放的研究受到日益广泛的关注。
2.3 O3对植物BVOCs排放的影响通过对已有文献进行检索, 获取O3单因子及复合因子胁迫相关研究, 通过式(1)将对照组与实验组的速率进行比较分析, 得到BVOCs速率增减的比例(图 2)。结果显示, O3胁迫产生的抑制作用在异戊二烯释放上更明显, 对于单萜烯的释放更多为促进作用。
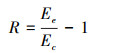
|
(1) |

|
| 图 2 O3对不同植物BVOCs排放速率的影响 Fig. 2 The effects of ozone on BVOCs emission rates among species |
式中, R表示BVOCs速率增减的比例, Ee表示实验组速率, Ec表示对照组速率。
图中数字为BVOCs速率增减的比例, 数字-1表示对照组检测到排放, 实验组未检测到排放;数字1表示对照组未检测到排放, 实验组检测到排放; 角标:r-O3耐受种, s-O3敏感种;字母区分多篇文章中的同一物种;数字区分O3浓度, 数字越大浓度越大。涉及的植物物种学名、拉丁名及文献来源见表 1; DMNT:(E)-4, 8-二甲基-1, 3, 7-壬三烯;OVOCs:其他挥发性有机化合物高浓度的O3暴露通常会抑制异戊二烯, 但不同树种的抑制不同。例如, 在高浓度O3刺激下, 番茄(Solanum lycopersicum L. var. Tiny Tim.)BVOCs释放量显著增加[95], 高浓度O3刺激并不影响白松(Pinus halepensis L.)释放BVOCs[96]。而我国长三角地区落叶阔叶林的异戊二烯排放量由于O3暴露受到抑制[5]。
| 物种 Species |
重复次数 Repetitions |
文献来源 References |
物种 Species |
重复次数 Repetitions |
文献来源 References |
|
| 银杏Ginkgo biloba L. | 3, 4 | [97, 109] | 黑芥Brassica nigra | 5, 4 | [100—101] | |
| 油松Pinus tabuliformis Carr. | 3 | [97] | 长豆角Ceratonia siliqua | 3 | [100] | |
| 多花巴豆Croton floribundus | 3 | [110] | 油橄榄Olea europaea | 3 | [100] | |
| 野甘蓝Brassica oleracea subsp. Capitata | 5(8) | [111] | 冬青栎Quercus ilex | 3, 6, 4 | [100, 112—113] | |
| 蒙古栎Quercus mongolica var. crispula | 4 | [114] | Quercus ilex rotundifolia | 3 | [100, 115] | |
| 枹栎Quercus serrata | 4 | [114] | 杂交杨Populus tremula× tremuloides(Clone55, Clone110) | 4 | [108, 115—116] | |
| 柔毛栎Quercus pubescens | 4, 5 | [99, 117] | 白杨树Populus tremuloides(271;42E) | 3, 30* | [108, 116] | |
| 白杨Populus alba | 4 | [118] | 杂交落叶松Larix gmelinii var. japonica× Larix kaempferi | 3 | [119] | |
| 黑杨Populus nigra | 4 | [102] | 垂枝桦Betula pendula Roth(Clone4;Clone80) | 4 | [120] | |
| 美洲黑杨Populus deltoides(clone 546) | 3, 3 | [98, 121] | 青杨Cathay poplar | 3 | [122] | |
| 白芥Sinapis alba | 5 | [101] | 欧洲云杉Picea abies | 4(24) | [123] | |
| 野芥Sinapis arvensis | 5 | [101] | 欧洲山杨Populus tremula(GT2.2;GT5.2) | 4 | [124] | |
| 欧洲白菜Brassica napus | 5, 3(4) | [101, 125] | 苏格兰松Pinus sylvestris L. | 4(18), 4(18) | [49, 87] | |
| 括号内表示每次重复的植物棵数, 若无标注则表示每次重复为一株植物, *表示该实验中测量了30片叶子的排放速率 | ||||||
植物对O3不同的响应主要取决于植物释放的BVOCs类型[97], O3暴露浓度、时间[98—99], 植物类型[100—101], 以及植物叶片的不同生长阶段[102]等因素。短期O3暴露与长期O3暴露对植物BVOCs释放的影响不同。玫瑰花卉的倍半萜烯释放速率在急性O3暴露下增加, 而长期暴露使其单萜烯与倍半萜烯释放速率均下降, 受合成酶底物的限制[103], 且在长期暴露下植物将BVOC排放改为含氧和反应性较低的化合物。某些植物在长期O3熏蒸条件下会促进异戊二烯的排放, 高浓度O3刺激其防御机制。单萜排放对长期暴露于O3的敏感性较低, 对急性暴露应激反应较大[104], 由于其合成后存储在植物中。春季, O3浓度升高降低了挪威云杉(Picea abies)和苏格兰松(Pinus sylvestris)地上部分BVOCs的排放量, 因为植物的休眠恢复受到抑制[105];夏季的结果与之相反, 两个物种地上部分的BVOCs的排放都随着O3的增加而增加, 此时, O3诱导的单萜烯释放是主要的挥发性防御反应。此外, O3还会增强变暖对植物BVOCs排放的刺激作用, 植物通过加快发育增加了合成萜烯类化合物的能力, 以保护植物免受高浓度O3胁迫的影响[66]。
已有部分研究对O3影响异戊二烯排放的生理生化机制进行了探究。O3会破坏/抑制BVOCs合成酶活性从而影响BVOCs的释放, 通过影响叶片表皮蜡的生物合成, 从而影响BVOCs的释放, 但O3还可能影响植物的信号转导途径, 从而诱导BVOCs的产生[106]。在低每小时臭氧浓度超过40nmol/mol部分的积累值(AOT40)下, 随着O3暴露的增加, 异戊二烯的排放量有一定的增长, 但随着O3暴露的增加, 异戊二烯的排放量总体呈线性下降, 这可能是由氧化防御机制引起的[5, 107]。O3暴露可能导致胞间二氧化碳的增加, 并增加产生异戊二烯所需的磷酸烯醇丙酮酸盐的消耗, 从而降低异戊二烯的排放速率[98]。异戊二烯排放的减少也可能是由于抑制了蛋白质合成的表达和活性[108]。
3 植物BVOCs排放对O3的影响植物BVOCs是O3形成的前体物之一, 对近地面O3的生成及其远距离运输具有一定的作用, 且其可作为植物抵抗O3胁迫的保护剂, 并帮助清除大气中的O3。目前, 部分城市绿地的平均BVOCs排放强度高于郊区森林[126], 部分地区地表O3浓度随归一化植被指数(NDVI)增加呈逻辑斯蒂增长, BVOCs是植被良好地区O3浓度高的原因[127]。
BVOCs能够通过与NOx反应以及与O3的光解产物反应等途径直接和间接增加近地面O3浓度。当大气中NOx浓度较高时不利于O3的形成, 而植物释放的BVOCs增加了大气中VOC的浓度, 使得VOC/NOx趋向最佳生产区, 增加近地面O3浓度。BVOCs还能与O3光解产物反应, 使O3光解产生的羟基自由基快速氧化为过氧自由基, 一氧化氮转化为二氧化氮, 从而增强O3的形成[128], 影响大气的还原性。模型模拟显示夏季BVOCs对O3形成的贡献高达16%—20%[22], 在热浪期间甚至高达60%[17]。与AVOCs相比, 异戊二烯的排放量占O3形成潜能(OFP)的一半(49.5%)[14]。
异戊二烯的氧化产物在O3远距离生成中起到一定作用, 其能与NOx反应产生有机硝酸盐, 其中一种为过氧乙酰硝酸酯(PAN)[53], 是光化学烟雾的主要成分, 在低温下比较稳定, 但在较高温度下又能分解形成NOx。因此在温度较低的高海拔地区产生的PAN能在下降到温度较高的低海拔地区之前被远距离运输, 之后又被分解, 使得NOx被远距离运输, 导致远距离O3的产生[129—130]。
此外, 植物存在气孔及非气孔的O3沉积[131], 当城市树木暴露在由土壤不透水性和根系器官的约束引起的干旱胁迫下, 气孔活性损伤时, 树冠内的气孔活动减弱, 会发生额外的非气孔O3沉积过程[131]。夏季柑橘树可以利用气孔和非气孔机制吸收O3来改善果园的空气质量[24]。吸收的O3可在细胞间隙累积, 会导致壁细胞氧化, 对光合器官造成损伤, 使植物的光合作用和生理代谢技能受到限制, 对生长速率、生物量的积累有不利影响, 加速植物的衰老[132]。而异戊二烯可以作为羟基自由基的清除剂以及作为光合作用膜和大分子免受氧化损伤的保护剂[133—134], 单萜可以代替光呼吸, 作为高温下产生的活性氧的清除剂, 以防止高温下的光损伤[135]。在同时具有高温和高O3浓度的条件下, BVOCs可作为抗氧化剂, 通过抑制O3或活性氧, 稳定对高温和高O3浓度敏感的膜[66, 134], 在压力条件下维持性能, 并帮助清除大气中的O3[118]。
4 研究展望植物具有滞尘[136]、净化空气、涵养水源、固碳释氧, 消除疲劳、减轻压力等作用。城市绿地在缓解城市环境问题、改善人居环境等方面发挥着不可替代的作用。随着城市居民对人居环境质量要求的不断提高, 增加城市绿色空间已成为城市生态建设的一个重要组成部分。但植物释放的BVOCs也有一些不容忽视的负面影响[137], 不合理的树种选择和配置导致植物释放的BVOCs增加, 进而加剧雾霾和O3污染。
BVOCs对植物、人体及环境的作用已取得较大的进展。随着分离、提取技术的进步, 其对植物本身及人体健康的作用进一步得到验证。在城市居民对人居环境质量要求提高的背景下, BVOCs与环境之间的相互作用备受关注。BVOCs对近地面O3、次级有机气溶胶等二次污染物形成具有重要贡献, 不断有学者提出新的模型方法以更精确估算BVOCs排放量, 但不同植物对污染的抗性不同, 其BVOCs的释放会受到环境因子的影响。因此, 需要加强排放估算模型的本地化, 深入探究环境胁迫对BVOCs释放影响的内部机理机制, 充分考虑植物BVOCs排放水平及其抗性之间的平衡。具体来说, 包括以下几个方面:
(1) 加强本地树种BVOCs排放因子的实测及精细化尺度BVOCs排放量估算。我国植被覆盖面积广且物种丰富, 部分树种的排放速率会随着经纬度等的变化发生改变[138—140], 因此建立本地物种的排放速率数据库尤为重要。早期测量多限于异戊二烯和单萜烯, 且多采用静态封闭式采样法, 会导致较大的误差。随着技术的发展, 目前多采用动态封闭式采样法, 可以进一步提升精度[141]。而目前关于BVOCs排放量估算的研究中, 由于树种排放速率的缺失, 部分研究可能会选择同科属植物替代, 或基于粗糙的植被空间分布仅考虑优势树种, 会导致BVOCs的估算结果产生较大的误差。
(2) 进一步探究城市中常见树种BVOCs释放对复合胁迫的响应。已有研究多基于单一环境条件和单一种属水平的比较, 而在城市环境中的植物BVOCs释放受到复合因素的影响。关于复合作用的研究较少, 且各研究结果存在差异[116, 142]。O3浓度升高和增温对欧洲山杨异戊二烯排放的影响依据基因型的不同而异, 但在田间暴露环境下发现O3浓度升高和增温对挪威云杉的异戊二烯排放没有显著影响, 对单萜烯和倍半萜烯排放的影响更显著[123]。在O3和二氧化碳对异戊二烯排放的复合影响研究, 也得到了相似的结果[109]。此外, 随着全球气候变化, 极端天气的增加, 是否会影响BVOCs的释放也亟待研究。综上, 需进一步探究复合胁迫对植物BVOCs排放的影响及其内在机理。
(3) 平衡树种的生态系统服务与其BVOCs排放对O3生成的贡献。城市景观建设中, 对树种选择与配置时, 除了满足生态、景观等功能外, 还需考虑植物对环境质量和人体身心健康的潜在的负面影响。植物BVOCs会一定程度上加剧O3形成, 但BVOCs也是植物抵御环境胁迫的保护剂, 高排放树种相较低排放树种能更好抵御O3等环境胁迫, 更好的适应城市环境, 并提供更多的生态系统服务。如何选择合适的树种, 优化其配置, 从而减少其负面影响, 提升其生态系统服务, 亟需开展更多的研究。
| [1] |
刘峰, 朱永官, 王效科. 我国地面臭氧污染及其生态环境效应. 生态环境, 2008(4): 1674-1679. DOI:10.3969/j.issn.1674-5906.2008.04.069 |
| [2] |
Liu Y M, Wang T. Worsening urban ozone pollution in China from 2013 to 2017-Part 1: the complex and varying roles of meteorology. Atmospheric Chemistry and Physics, 2020, 20(11): 6305-6321. DOI:10.5194/acp-20-6305-2020 |
| [3] |
Feng Z Z, Calatayud V, Zhu J G, Kobayashi K. Ozone exposure- and flux-based response relationships with photosynthesis of winter wheat under fully open air condition. Science of the Total Environment, 2018, 619/620: 1538-1544. DOI:10.1016/j.scitotenv.2017.10.089 |
| [4] |
WHO air quality guidelines for particulate matter, ozone, nitrogen dioxide and sulfur dioxide: global update 2005. Summary of risk assessment, 2006.
|
| [5] |
Wang Y T, Zhao Y, Zhang L, Zhang J, Liu Y. Modified regional biogenic VOC emissions with actual ozone stress and integrated land cover information: a case study in Yangtze River Delta, China. Science of the Total Environment, 2020, 727: 138703. DOI:10.1016/j.scitotenv.2020.138703 |
| [6] |
严刚, 薛文博, 雷宇, 宁淼, 武卫玲, 刘伟. 我国臭氧污染形势分析及防控对策建议. 环境保护, 2020, 48(15): 15-19. DOI:10.14026/j.cnki.0253-9705.2020.15.003 |
| [7] |
Gao Y, Ma M C, Yan F F, Su H, Wang S X, Liao H, Zhao B, Wang X M, Sun Y L, Hopkins J R, Chen Q, Fu P Q, Lewis A C, Qiu Q H, Yao X H, Gao H W. Impacts of biogenic emissions from urban landscapes on summer ozone and secondary organic aerosol formation in megacities. Science of the Total Environment, 2022, 814: 152654. DOI:10.1016/j.scitotenv.2021.152654 |
| [8] |
冯兆忠, 袁相洋. 臭氧浓度升高对植物源挥发性有机化合物(BVOCs)影响的研究进展. 环境科学, 2018, 39(11): 5257-5265. DOI:10.13227/j.hjkx.201802075 |
| [9] |
胡春芳, 袁相洋, 田媛, 冯兆忠. 常见花卉植物释放挥发性有机化合物的研究进展. 生态学杂志, 2018, 37(2): 588-595. DOI:10.13292/j.1000-4890.201802.037 |
| [10] |
Peñuelas J, Llusià J. Plant VOC emissions: making use of the unavoidable. Trends in Ecology & Evolution, 2004, 19(8): 402-404. |
| [11] |
Guenther A, Hewitt C N, Erickson D, Fall R, Geron C, Graedel T, Harley P, Klinger L, Lerdau M, McKay W A, Pierce T, Scholes B, Steinbrecher R, Tallamraju R, Taylor J, Zimmerman P. A global model of natural volatile organic compound emissions. Journal of Geophysical Research, 1995, 100(D5): 8873. DOI:10.1029/94JD02950 |
| [12] |
Mo Z W, Shao M, Wang W J, Liu Y, Wang M, Lu S H. Evaluation of biogenic isoprene emissions and their contribution to ozone formation by ground-based measurements in Beijing, China. The Science of the Total Environment, 2018, 627: 1485-1494. DOI:10.1016/j.scitotenv.2018.01.336 |
| [13] |
高超, 张学磊, 修艾军, 黄凌, 赵红梅, 王堃, 童清清. 中国生物源挥发性有机物(BVOCs)时空排放特征研究. 环境科学学报, 2019, 39(12): 4140-4151. |
| [14] |
赵芸程, 李杰, 杜惠云, 王辉, 王自发, 杨文夷, 吴其重. 北京夏季近地面臭氧及其来源的数值模拟研究. 环境科学学报, 2019, 39(7): 2315-2328. DOI:10.13671/j.hjkxxb.2019.0078 |
| [15] |
Li H D, Meier F, Lee X H, Chakraborty T, Liu J F, Schaap M, Sodoudi S. Interaction between urban heat island and urban pollution island during summer in Berlin. The Science of the Total Environment, 2018, 636: 818-828. DOI:10.1016/j.scitotenv.2018.04.254 |
| [16] |
Li D, Sun T, Liu M F, Yang L, Wang L L, Gao Z Q. Contrasting responses of urban and rural surface energy budgets to heat waves explain synergies between urban heat Islands and heat waves. Environmental Research Letters, 2015, 10(5): 054009. DOI:10.1088/1748-9326/10/5/054009 |
| [17] |
Churkina G, Kuik F, Bonn B, Lauer A, Grote R, Tomiak K, Butler T M. Effect of VOC emissions from vegetation on air quality in berlin during a heatwave. Environmental Science & Technology, 2017, 51(11): 6120-6130. |
| [18] |
Wang H L, Wu K, Liu Y M, Sheng B S, Lu X, He Y P, Xie J L, Wang H C, Fan S J. Role of heat wave-induced biogenic VOC enhancements in persistent ozone episodes formation in Pearl River Delta. Journal of Geophysical Research: Atmospheres, 2021, 126(12). |
| [19] |
Ramacher M O P, Kari M, Bieser J, Feldner, J. The impact of BVOC emissions from urban trees on O3 production in urban areas under heat-period conditions. Air Pollution Modeling and Its Application XXVⅡ, 2021, 241-247. |
| [20] |
Seto K C, Güneralp B, Hutyra L R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(40): 16083-16088. DOI:10.1073/pnas.1211658109 |
| [21] |
Li L Y, Yang W Z, Xie S D, Wu Y. Estimations and uncertainty of biogenic volatile organic compound emission inventory in China for 2008-2018. The Science of the Total Environment, 2020, 733: 139301. DOI:10.1016/j.scitotenv.2020.139301 |
| [22] |
Cao J, Situ S P, Hao Y F, Xie S D, Li L Y. Enhanced summertime ozone and SOA from biogenic volatile organic compound (BVOC) emissions due to vegetation biomass variability during 1981-2018 in China. Atmospheric Chemistry and Physics, 2022, 22(4): 2351-2364. DOI:10.5194/acp-22-2351-2022 |
| [23] |
Wu J, Long J Y, Liu H X, Sun G P, Li J, Xu L J, Xu C Y. Biogenic volatile organic compounds from 14 landscape woody species: tree species selection in the construction of urban greenspace with forest healthcare effects. Journal of Environmental Management, 2021, 300: 113761. DOI:10.1016/j.jenvman.2021.113761 |
| [24] |
Fares S, Park J H, Ormeno E, Gentner D R, McKay M, Loreto F, Karlik J, Goldstein A H. Ozone uptake by Citrus trees exposed to a range of ozone concentrations. Atmospheric Environment, 2010, 44(28): 3404-3412. DOI:10.1016/j.atmosenv.2010.06.010 |
| [25] |
孔垂华, 徐涛, 胡飞, 黄寿山. 环境胁迫下植物的化感作用及其诱导机制. 生态学报, 2000, 20(5): 849-854. DOI:10.3321/j.issn:1000-0933.2000.05.022 |
| [26] |
Dudareva N, Pichersky E, Gershenzon J. Biochemistry of plant volatiles. Plant Physiology, 2004, 135(4): 1893-1902. DOI:10.1104/pp.104.049981 |
| [27] |
Peñuelas J, Staudt M. BVOCs and global change. Trends in Plant Science, 2010, 15(3): 133-144. DOI:10.1016/j.tplants.2009.12.005 |
| [28] |
Peñuelas J. BVOCs: plant defense against climate warming?. Trends in Plant Science, 2003, 8(3): 105-109. DOI:10.1016/S1360-1385(03)00008-6 |
| [29] |
Heuberger E, Hongratanaworakit T, Böhm C, Weber R, Buchbauer G. Effects of chiral fragrances on human autonomic nervous system parameters and self-evaluation. Chemical Senses, 2001, 26(3): 281-292. DOI:10.1093/chemse/26.3.281 |
| [30] |
方治国, 欧阳志云, 胡利锋, 王效科, 林学强. 北京市夏季空气微生物群落结构和生态分布. 生态学报, 2005, 25(1): 83-88. |
| [31] |
Ilmberger J, Heuberger E, Mahrhofer C, Dessovic H, Kowarik D, Buchbauer G. The influence of essential oils on human attention. I: alertness. Chemical Senses, 2001, 26(3): 239-245. DOI:10.1093/chemse/26.3.239 |
| [32] |
Lun X X, Lin Y, Chai F H, Fan C, Li H, Liu J F. Reviews of emission of biogenic volatile organic compounds (BVOCs) in Asia. Journal of Environmental Sciences: China, 2020, 95: 266-277. DOI:10.1016/j.jes.2020.04.043 |
| [33] |
高岩. 北京市绿化树木挥发性有机物释放动态及其对人体健康的影响[D]. 北京: 北京林业大学, 2005.
|
| [34] |
Najar B, Nardi V, Cervelli C, Mancianti F, Nardoni S, Ebani V V, Pistelli L. Helichrysum araxinum Takht. ex Kirp. grown in Italy: volatiloma composition and in vitro antimicrobial activity. Zeitschrift Fur Naturforschung C, Journal of Biosciences, 2020, 75(7/8): 265-270. |
| [35] |
Houweling S, Dentener F, Lelieveld J. The impact of nonmethane hydrocarbon compounds on tropospheric photochemistry. Journal of Geophysical Research: Atmospheres, 1998, 103(D9): 10673-10696. DOI:10.1029/97JD03582 |
| [36] |
Wang Q G, Han Z W, Wang T J, Zhang R J. Impacts of biogenic emissions of VOC and NOx on tropospheric ozone during summertime in Eastern China. The Science of the Total Environment, 2008, 395(1): 41-49. DOI:10.1016/j.scitotenv.2008.01.059 |
| [37] |
Han D M, Gao S, Fu Q Y, Cheng J P, Chen X J, Xu H, Liang S, Zhou Y, Ma Y N. Do volatile organic compounds (VOCs) emitted from petrochemical industries affect regional PM2.5?. Atmospheric Research, 2018, 209: 123-130. DOI:10.1016/j.atmosres.2018.04.002 |
| [38] |
Fehsenfeld F, Calvert J, Fall R, Goldan P, Guenther A B, Hewitt C N, Lamb B, Liu S, Trainer M, Westberg H, Zimmerman P. Emissions of volatile organic compounds from vegetation and the implications for atmospheric chemistry. Global Biogeochemical Cycles, 1992, 6(4): 389-430. DOI:10.1029/92GB02125 |
| [39] |
Rohmer M, Knani M, Simonin P, Sutter B, Sahm H. Isoprenoid biosynthesis in bacteria: a novel pathway for the early steps leading to isopentenyl diphosphate. The Biochemical Journal, 1993, 295(Pt 2): 517-524. |
| [40] |
Lichtenthaler H K, Schwender J, Disch A, Rohmer M. Biosynthesis of isoprenoids in higher plant chloroplasts proceeds via a mevalonate-independent pathway. FEBS Letters, 1997, 400(3): 271-274. DOI:10.1016/S0014-5793(96)01404-4 |
| [41] |
Tingey D T, Turner D P, Weber J A. Factors controlling the emissions of monoterpenes and other volatile organic compounds. Trace Gas Emissions by Plants, 1991, 93-119. |
| [42] |
Sharkey T D, Loreto F. Water stress, temperature, and light effects on the capacity for isoprene emission and photosynthesis of kudzu leaves. Oecologia, 1993, 95(3): 328-333. DOI:10.1007/BF00320984 |
| [43] |
Guenther A B, Zimmerman P R, Harley P C, Monson R K, Fall R. Isoprene and monoterpene emission rate variability: model evaluations and sensitivity analyses. Journal of Geophysical Research, 1993, 98(D7): 12609. DOI:10.1029/93JD00527 |
| [44] |
Leung D Y C, Wong P, Cheung B K H, Guenther A. Improved land cover and emission factors for modeling biogenic volatile organic compounds emissions from Hong Kong. Atmospheric Environment, 2010, 44(11): 1456-1468. DOI:10.1016/j.atmosenv.2010.01.012 |
| [45] |
Owen S M, Harley P, Guenther A, Hewitt C N. Light dependency of VOC emissions from selected Mediterranean plant species. Atmospheric Environment, 2002, 36(19): 3147-3159. DOI:10.1016/S1352-2310(02)00235-2 |
| [46] |
任琴, 谢明惠, 张青文, 齐钢, 刘小侠. 不同温度、光照对虫害紫茎泽兰挥发物释放的影响. 生态学报, 2010, 30(11): 3080-3086. |
| [47] |
Guidolotti G, Pallozzi E, Gavrichkova O, Scartazza A, Mattioni M, Loreto F, Calfapietra C. Emission of constitutive isoprene, induced monoterpenes, and other volatiles under high temperatures in Eucalyptus camaldulensis: a 13 C labelling study. Plant, Cell & Environment, 2019, 42(6): 1929-1938. |
| [48] |
Bamberger I, Ruehr N K, Schmitt M, Gast A, Wohlfahrt G, Arneth A. Isoprene emission and photosynthesis during heatwaves and drought in black locust. Biogeosciences, 2017, 14(15): 3649-3667. DOI:10.5194/bg-14-3649-2017 |
| [49] |
Kivimäenpää M, Ghimire R P, Sutinen S, Häikiö E, Kasurinen A, Holopainen T, Holopainen J K. Increases in volatile organic compound emissions of Scots pine in response to elevated ozone and warming are modified by herbivory and soil nitrogen availability. European Journal of Forest Research, 2016, 135(2): 343-360. DOI:10.1007/s10342-016-0939-x |
| [50] |
Constable J V H, Litvak M E, Greenberg J P, Monson R K. Monoterpene emission from coniferous trees in response to elevated CO2 concentration and climate warming. Global Change Biology, 1999, 5(3): 252-267. DOI:10.1046/j.1365-2486.1999.00212.x |
| [51] |
Wagner P, Kuttler W. Biogenic and anthropogenic isoprene in the near-surface urban atmosphere—a case study in Essen, Germany. The Science of the Total Environment, 2014, 475: 104-115. DOI:10.1016/j.scitotenv.2013.12.026 |
| [52] |
Carter W P L. Development of ozone reactivity scales for volatile organic compounds. Air & Waste, 1994, 44(7): 881-899. |
| [53] |
Wang J L, Chew C, Chang C Y, Liao W C, Lung S C C, Chen W N, Lee P J, Lin P H, Chang C C. Biogenic isoprene in subtropical urban settings and implications for air quality. Atmospheric Environment, 2013, 79: 369-379. DOI:10.1016/j.atmosenv.2013.06.055 |
| [54] |
Karlsson T, Rinnan R, Holst T. Variability of BVOC emissions from commercially used willow (Salix spp.) varieties. Atmosphere, 2020, 11(4): 356. DOI:10.3390/atmos11040356 |
| [55] |
Benjamin M T, Sudol M, Bloch L, Winer A M. Low-emitting urban forests: a taxonomic methodology for assigning isoprene and monoterpene emission rates. Atmospheric Environment, 1996, 30(9): 1437-1452. DOI:10.1016/1352-2310(95)00439-4 |
| [56] |
Loreto F, Sharkey T D. A gas-exchange study of photosynthesis and isoprene emission inQuercus rubra L. Planta, 1990, 182(4): 523-531. DOI:10.1007/BF02341027 |
| [57] |
Staudt M, Seufert G. Light-dependent emission of monoterpenes by holm oak (Quercus ilex L.). Naturwissenschaften, 1995, 82(2): 89-92. DOI:10.1007/BF01140148 |
| [58] |
Martin D M, Gershenzon J, Bohlmann J. Induction of volatile terpene biosynthesis and diurnal emission by methyl jasmonate in foliage of Norway spruce. Plant Physiology, 2003, 132(3): 1586-1599. DOI:10.1104/pp.103.021196 |
| [59] |
Yang W Z, Cao J, Wu Y, Kong F L, Li L Y. Review on plant terpenoid emissions worldwide and in China. The Science of the Total Environment, 2021, 787: 147454. DOI:10.1016/j.scitotenv.2021.147454 |
| [60] |
Loreto F, Schnitzler J P. Abiotic stresses and induced BVOCs. Trends in Plant Science, 2010, 15(3): 154-166. DOI:10.1016/j.tplants.2009.12.006 |
| [61] |
Gomez L G, Loubet B, Lafouge F, Ciuraru R, Bsaibes S, Kammer J, Buysse P, Durand B, Gueudet J C, Fanucci O, Zurfluh O, Decuq C, Truong F, Gros V, Boissard C. Effect of senescence on biogenic volatile organic compound fluxes in wheat plants. Atmospheric Environment, 2021, 266: 118665. DOI:10.1016/j.atmosenv.2021.118665 |
| [62] |
Estell R E, James D K, Fredrickson E L, Anderson D M. Within-plant distribution of volatile compounds on the leaf surface of Flourensia cernua. Biochemical Systematics and Ecology, 2013, 48: 144-150. DOI:10.1016/j.bse.2012.11.020 |
| [63] |
Hartikainen K, Kivimäenpää M, Nerg A M, Mäenpää M, Oksanen E, Rousi M, Holopainen T. Elevated temperature and ozone modify structural characteristics of silver birch (Betula pendula) leaves. Tree Physiology, 2020, 40(4): 467-483. DOI:10.1093/treephys/tpz127 |
| [64] |
van Meeningen Y, Wang M, Karlsson T, Seifert A, Schurgers G, Rinnan R, Holst T. Isoprenoid emission variation of Norway spruce across a European latitudinal transect. Atmospheric Environment, 2017, 170: 45-57. DOI:10.1016/j.atmosenv.2017.09.045 |
| [65] |
Kim J C, Kim K J, Kim D S, Han J S. Seasonal variations of monoterpene emissions from coniferous trees of different ages in Korea. Chemosphere, 2005, 59(11): 1685-1696. DOI:10.1016/j.chemosphere.2004.10.048 |
| [66] |
Minna K, Johanna R, Hanna V, Elina H, Holopainen Jarmo K, Toini H. Effects of elevated ozone and warming on terpenoid emissions and concentrations of Norway spruce depend on needle phenology and age. Tree Physiology, 2022, 2022F. |
| [67] |
Baghi R, Helmig D, Guenther A, Duhl T, Daly R. Contribution of flowering trees to urban atmospheric biogenic volatile organic compound emissions. Biogeosciences, 2012, 9(10): 3777-3785. DOI:10.5194/bg-9-3777-2012 |
| [68] |
Ndah F, Valolahti H, Schollert M, Michelsen A, Rinnan R, Kivimäenpää M. Influence of increased nutrient availability on biogenic volatile organic compound (BVOC) emissions and leaf anatomy of subarctic dwarf shrubs under climate warming and increased cloudiness. Annals of Botany, 2022, 129(4): 443-455. DOI:10.1093/aob/mcac004 |
| [69] |
Baggesen N, Li T, Seco R, Holst T, Michelsen A, Rinnan R. Phenological stage of tundra vegetation controls bidirectional exchange of BVOCs in a climate change experiment on a subarctic heath. Global Change Biology, 2021, 27(12): 2928-2944. DOI:10.1111/gcb.15596 |
| [70] |
Ghimire R P, Silfver T, Myller K, Oksanen E, Holopainen J K, Mikola J. BVOC emissions from a subarctic ecosystem, as controlled by insect herbivore pressure and temperature. Ecosystems, 2021, 1-20. |
| [71] |
Rasheed M U, Kivimäenpää M, Kasurinen A. Emissions of biogenic volatile organic compounds (BVOCs) from the rhizosphere of Scots pine (Pinus sylvestris) seedlings exposed to warming, moderate N addition and bark herbivory by large pine weevil (Hylobius abietis). Plant and Soil, 2021, 463(1/2): 379-394. |
| [72] |
Allmann S, Späthe A, Bisch-Knaden S, Kallenbach M, Reinecke A, Sachse S, Baldwin I T, Hansson B S. Feeding-induced rearrangement of green leaf volatiles reduces moth oviposition. eLife, 2013, 2: e00421. DOI:10.7554/eLife.00421 |
| [73] |
Piesik D, Pańka D, Delaney K J, Skoczek A, Lamparski R, Weaver D K. Cereal crop volatile organic compound induction after mechanical injury, beetle herbivory (Oulema spp.), or fungal infection (Fusarium spp.). Journal of Plant Physiology, 2011, 168(9): 878-886. DOI:10.1016/j.jplph.2010.11.010 |
| [74] |
Ponzio C, Gols R, Pieterse C M J, Dicke M. Ecological and phytohormonal aspects of plant volatile emission in response to single and dual infestations with herbivores and phytopathogens. Functional Ecology, 2013, 27(3): 587-598. DOI:10.1111/1365-2435.12035 |
| [75] |
Anna W-P. Volatile Organic Compound Emissions by Winter Wheat Plants (Triticum aestivum L.) under Fusarium spp. Infestation and Various Abiotic Conditions. Polish Journal of Environmental Studies, 2011, 20(5): 1335-1342. |
| [76] |
Obara N, Hasegawa M, Kodama O. Induced volatiles in elicitor-treated and rice blast fungus-inoculated rice leaves. Bioscience, Biotechnology, and Biochemistry, 2002, 66(12): 2549-2559. DOI:10.1271/bbb.66.2549 |
| [77] |
Halitschke R, Ziegler J, Keinänen M, Baldwin I T. Silencing of hydroperoxide lyase and allene oxide synthase reveals substrate and defense signaling crosstalk in Nicotiana attenuata. The Plant Journal: for Cell and Molecular Biology, 2004, 40(1): 35-46. DOI:10.1111/j.1365-313X.2004.02185.x |
| [78] |
Ameye M, Allmann S, Verwaeren J, Smagghe G, Haesaert G, Schuurink R C, Audenaert K. Green leaf volatile production by plants: a meta-analysis. The New Phytologist, 2018, 220(3): 666-683. DOI:10.1111/nph.14671 |
| [79] |
Janson R W. Monoterpene emissions from Scots pine and Norwegian spruce. Journal of Geophysical Research: Atmospheres, 1993, 98(D2): 2839-2850. DOI:10.1029/92JD02394 |
| [80] |
Lüpke M, Leuchner M, Steinbrecher R, Menzel A. Impact of summer drought on isoprenoid emissions and carbon sink of three Scots pine provenances. Tree Physiology, 2016, 36(11): 1382-1399. |
| [81] |
Caser M, Chitarra W, D'Angiolillo F, Perrone I, Demasi S, Lovisolo C, Pistelli L, Pistelli L, Scariot V. Drought stress adaptation modulates plant secondary metabolite production in Salvia dolomitica Codd. Industrial Crops and Products, 2019, 129: 85-96. DOI:10.1016/j.indcrop.2018.11.068 |
| [82] |
Hu B, Jarosch A M, Gauder M, Graeff-Hönninger S, Schnitzler J P, Grote R, Rennenberg H, Kreuzwieser J. VOC emissions and carbon balance of two bioenergy plantations in response to nitrogen fertilization: a comparison of Miscanthus and Salix. Environmental Pollution: Barking, Essex: 1987, 2018, 237: 205-217. |
| [83] |
Huang X R, Lai J M, Liu Y F, Zheng L L, Fang X, Song W, Yi Z. Biogenic volatile organic compound emissions from Pinus massoniana and Schima superba seedlings: their responses to foliar and soil application of nitrogen. Science of The Total Environment, 2020, 705: 135761. DOI:10.1016/j.scitotenv.2019.135761 |
| [84] |
Loreto F, Fares S. Biogenic volatile organic compounds and their impacts on biosphere-atmosphere interactions. Developments in Environmental Science, 2013, 13: 57-75. |
| [85] |
Llusia J, Bermejo-Bermejo V, Calvete-Sogo H, Peñuelas J. Decreased rates of terpene emissions in Ornithopus compressus L. and Trifolium striatum L. by ozone exposure and nitrogen fertilization. Environmental Pollution: Barking, Essex: 1987, 2014, 194: 69-77.
|
| [86] |
Blanch J S, Peñuelas J, Llusià J. Sensitivity of terpene emissions to drought and fertilization in terpene-storing Pinus halepensis and non-storing Quercus ilex. Physiologia Plantarum, 2007, 131(2): 211-225. |
| [87] |
Ghimire R P, Kivimäenpää M, Kasurinen A, Häikiö E, Holopainen T, Holopainen J K. Herbivore-induced BVOC emissions of Scots pine under warming, elevated ozone and increased nitrogen availability in an open-field exposure. Agricultural and Forest Meteorology, 2017, 242: 21-32. DOI:10.1016/j.agrformet.2017.04.008 |
| [88] |
Carriero G, Brunetti C, Fares S, Hayes F, Hoshika Y, Mills G, Tattini M, Paoletti E. BVOC responses to realistic nitrogen fertilization and ozone exposure in silver birch. Environmental Pollution: Barking, Essex: 1987, 2016, 213: 988-995. |
| [89] |
Tiiva P, Häikiö E, Kasurinen A. Impact of warming, moderate nitrogen addition and bark herbivory on BVOC emissions and growth of Scots pine (Pinus sylvestris L.) seedlings. Tree Physiology, 2018, 38(10): 1461-1475. DOI:10.1093/treephys/tpy029 |
| [90] |
Jasoni R, Kane C, Green C, Peffley E, Tissue D, Thompson L, Payton P, Paré P W. Altered leaf and root emissions from onion (Allium cepa L.) grown under elevated CO2 conditions. Environmental and Experimental Botany, 2004, 51(3): 273-280. DOI:10.1016/j.envexpbot.2003.11.006 |
| [91] |
Rapparini F, Baraldi R, Miglietta F, Loreto F. Isoprenoid emission in trees of Quercus pubescens and Quercus ilex with lifetime exposure to naturally high CO2 environment+. Plant, Cell and Environment, 2004, 27(4): 381-391. DOI:10.1111/j.1365-3040.2003.01151.x |
| [92] |
Possell M, Nicholas Hewitt C, Beerling D J. The effects of glacial atmospheric CO2 concentrations and climate on isoprene emissions by vascular plants. Global Change Biology, 2005, 11(1): 60-69. DOI:10.1111/j.1365-2486.2004.00889.x |
| [93] |
Tiiva P, Julkunen-Tiitto R, Häikiö E, Kasurinen A. Belowground responses of Scots pine (Pinus sylvestris) seedlings to experimental warming, moderate nitrogen addition, and bark herbivory. Canadian Journal of Forest Research, 2019, 49(6): 647-660. DOI:10.1139/cjfr-2018-0099 |
| [94] |
Yu H, Blande J D. Diurnal variation in BVOC emission and CO2 gas exchange from above- and belowground parts of two coniferous species and their responses to elevated O 3. Environmental Pollution: Barking, Essex: 1987, 2021, 278: 116830. |
| [95] |
Heiden A C, Hoffmann T, Kahl J, Kley D, Klockow D, Langebartels C, Mehlhorn H, Sandermann H Jr, Schraudner M, Schuh G, Wildt J. Emission of volatile organic compounds from ozone-exposed plants. Ecological Applications, 1999, 9(4): 1160-1167. DOI:10.1890/1051-0761(1999)009[1160:EOVOCF]2.0.CO;2 |
| [96] |
Peñuelas J, Llusià J, Gimeno B S. Effects of ozone concentrations on biogenic volatile organic compounds emission in the Mediterranean region. Environmental Pollution, 1999, 105(1): 17-23. DOI:10.1016/S0269-7491(98)00214-0 |
| [97] |
李德文, 史奕, 何兴元. O3浓度升高对银杏及油松BVOCs排放的影响. 环境科学, 2008, 29(2): 2300-2304. |
| [98] |
Yuan X Y, Feng Z Z, Liu S, Shang B, Li P, Xu Y S, Paoletti E. Concentration- and flux-based dose-responses of isoprene emission from poplar leaves and plants exposed to an ozone concentration gradient. Plant, Cell & Environment, 2017, 40(9): 1960-1971. |
| [99] |
Pinelli P, Tricoli D. A new approach to ozone plant fumigation: the Web-O3- Fumigation. Isoprene response to a gradient of ozone stress in leaves of Quercus pubescens. IForest-Biogeosciences and Forestry, 2008, 1(1): 22-26. DOI:10.3832/ifor0240-0010022 |
| [100] |
Llusià J, Peñuelas J, Gimeno B S. Seasonal and species-specific response of VOC emissions by Mediterranean woody plant to elevated ozone concentrations. Atmospheric Environment, 2002, 36(24): 3931-3938. DOI:10.1016/S1352-2310(02)00321-7 |
| [101] |
Saunier A, Blande J D. The effect of elevated ozone on floral chemistry of Brassicaceae species. Environmental Pollution, 2019, 255: 113257. DOI:10.1016/j.envpol.2019.113257 |
| [102] |
Fares S, Oksanen E, Lännenpää M, Julkunen-Tiitto R, Loreto F. Volatile emissions and phenolic compound concentrations along a vertical profile of Populus nigra leaves exposed to realistic ozone concentrations. Photosynthesis Research, 2010, 104(1): 61-74. DOI:10.1007/s11120-010-9549-5 |
| [103] |
Yuan X Y, Feng Z Z, Hu C F, Zhang K, Qu L Y, Paoletti E. Effects of elevated ozone on the emission of volatile isoprenoids from flowers and leaves of rose (Rosa sp.) varieties. Environmental Pollution: Barking, Essex: 1987, 2021, 291: 118141.
|
| [104] |
Acton W J F, Jud W, Ghirardo A, Wohlfahrt G, Hewitt C N, Taylor J E, Hansel A. The effect of ozone fumigation on the biogenic volatile organic compounds (BVOCs) emitted from Brassica napus above- and below-ground. PLoS One, 2018, 13(12): e0208825. DOI:10.1371/journal.pone.0208825 |
| [105] |
Aalto J, Kolari P, Hari P, Kerminen V M, Schiestl-Aalto P, Aaltonen H, Levula J, Siivola E, Kulmala M, Bäck J. New foliage growth is a significant, unaccounted source for volatiles in boreal evergreen forests. Biogeosciences, 2014, 11(5): 1331-1344. DOI:10.5194/bg-11-1331-2014 |
| [106] |
Laothawornkitkul J, Taylor J E, Paul N D, Hewitt C N. Biogenic volatile organic compounds in the Earth system. New Phytologist, 2009, 183(1): 27-51. DOI:10.1111/j.1469-8137.2009.02859.x |
| [107] |
Kleist E., Heiden A. C., Langenbartels C., et al. Emission of oxygenated volatile organic compounds (VOC) from plants as a response to biotic and abiotic stress, 2002.
|
| [108] |
Calfapietra C, Wiberley A E, Falbel T G, Linskey A R, Mugnozza G S, Karnosky D F, Loreto F, Sharkey T D. Isoprene synthase expression and protein levels are reduced under elevated O3 but not under elevated CO2 (FACE) in field-grown aspen trees. Plant, Cell & Environment, 2007, 30(5): 654-661. |
| [109] |
Li D W, Chen Y, Shi Y, He X Y, Chen X. Impact of elevated CO2 and O3 concentrations on biogenic volatile organic compounds emissions from Ginkgo biloba. Bulletin of Environmental Contamination and Toxicology, 2009, 82(4): 473-477. DOI:10.1007/s00128-008-9590-7 |
| [110] |
da Silva Pedrosa G, de Oliveira D P, Bison J V S, Bugarelli R M, Cruz L S, de Souza S R. Biogenic volatile organic compounds emission of Brazilian Atlantic tree grown under elevated ozone in ambient controlled and field conditions. Bulletin of Environmental Contamination and Toxicology, 2020, 105(6): 958-966. DOI:10.1007/s00128-020-03056-9 |
| [111] |
Pinto D M, Tiiva P, Miettinen P, Joutsensaari J, Kokkola H, Nerg A M, Laaksonen A, Holopainen J K. The effects of increasing atmospheric ozone on biogenic monoterpene profiles and the formation of secondary aerosols. Atmospheric Environment, 2007, 41(23): 4877-4887. DOI:10.1016/j.atmosenv.2007.02.006 |
| [112] |
Vitale M, Salvatori E, Loreto F, Fares S, Manes F. Physiological responses of Quercus ilex leaves to water stress and acute ozone exposure under controlled conditions. Water, Air, and Soil Pollution, 2008, 189(1/2/3/4): 113-125. |
| [113] |
Loreto F, Pinelli P, Manes F, Kollist H. Impact of ozone on monoterpene emissions and evidence for an isoprene-like antioxidant action of monoterpenes emitted by Quercus ilex leaves. Tree Physiology, 2004, 24(4): 361-367. DOI:10.1093/treephys/24.4.361 |
| [114] |
Tani A, Ohno T, Saito T, Ito S, Yonekura T, Miwa M. Effects of ozone on isoprene emission from two major Quercus species native to East Asia. Journal of Agricultural Meteorology, 2017, 73(4): 195-202. DOI:10.2480/agrmet.D-17-00022 |
| [115] |
Blande J D, Tiiva P, Oksanen E, Holopainen J K. Emission of herbivore-induced volatile terpenoids from two hybrid aspen (Populus tremula×tremuloides) clones under ambient and elevated ozone concentrations in the field. Global Change Biology, 2007, 13(12): 2538-2550. DOI:10.1111/j.1365-2486.2007.01453.x |
| [116] |
Calfapietra C, Scarascia Mugnozza G, Karnosky D F, Loreto F, Sharkey T D. Isoprene emission rates under elevated CO2 and O3 in two field-grown aspen clones differing in their sensitivity to O3. The New Phytologist, 2008, 179(1): 55-61. DOI:10.1111/j.1469-8137.2008.02493.x |
| [117] |
Velikova V, Tsonev T, Pinelli P, Alessio G A, Loreto F. Localized ozone fumigation system for studying ozone effects on photosynthesis, respiration, electron transport rate and isoprene emission in field-grown Mediterranean oak species. Tree Physiology, 2005, 25(12): 1523-1532. DOI:10.1093/treephys/25.12.1523 |
| [118] |
Fares S, Barta C, Brilli F, Centritto M, Ederli L, Ferranti F, Pasqualini S, Reale L, Tricoli D, Loreto F. Impact of high ozone on isoprene emission, photosynthesis and histology of developing Populus alba leaves directly or indirectly exposed to the pollutant. Physiologia Plantarum, 2006, 128(3): 456-465. DOI:10.1111/j.1399-3054.2006.00750.x |
| [119] |
Mochizuki T, Watanabe M, Koike T, Tani A. Monoterpene emissions from needles of hybrid larch F1 (Larix gmelinii var. japonica×Larix kaempferi) grown under elevated carbon dioxide and ozone. Atmospheric Environment, 2017, 148: 197-202. DOI:10.1016/j.atmosenv.2016.10.041 |
| [120] |
Vuorinen T, Nerg A M, Vapaavuori E, Holopainen J K. Emission of volatile organic compounds from two silver birch (Betula pendula Roth) clones grown under ambient and elevated CO2 and different O3 concentrations. Atmospheric Environment, 2005, 39(7): 1185-1197. DOI:10.1016/j.atmosenv.2004.09.077 |
| [121] |
Yuan X Y, Calatayud V, Gao F, Fares S, Paoletti E, Tian Y, Feng Z Z. Interaction of drought and ozone exposure on isoprene emission from extensively cultivated poplar. Plant, Cell & Environment, 2016, 39(10): 2276-2287. |
| [122] |
Yuan X Y, Shang B, Xu Y S, Xin Y, Tian Y, Feng Z Z, Paoletti E. No significant interactions between nitrogen stimulation and ozone inhibition of isoprene emission in Cathay poplar. Science of the Total Environment, 2017, 601/602: 222-229. DOI:10.1016/j.scitotenv.2017.05.138 |
| [123] |
Kivimäenpää M, Riikonen J, Ahonen V, Tervahauta A, Holopainen T. Sensitivity of Norway spruce physiology and terpenoid emission dynamics to elevated ozone and elevated temperature under open-field exposure. Environmental and Experimental Botany, 2013, 90: 32-42. DOI:10.1016/j.envexpbot.2012.11.004 |
| [124] |
Hartikainen K, Nerg A M, Kivimäenpää M, Kontunen-Soppela S, Mäenpää M, Oksanen E, Rousi M, Holopainen T. Emissions of volatile organic compounds and leaf structural characteristics of European aspen (Populus tremula) grown under elevated ozone and temperature. Tree Physiology, 2009, 29(9): 1163-1173. DOI:10.1093/treephys/tpp033 |
| [125] |
Himanen S J, Nerg A M, Nissinen A, Pinto D M, Stewart C N Jr, Poppy G M, Holopainen J K. Effects of elevated carbon dioxide and ozone on volatile terpenoid emissions and multitrophic communication of transgenic insecticidal oilseed rape (Brassica napus). The New Phytologist, 2009, 181(1): 174-186. DOI:10.1111/j.1469-8137.2008.02646.x |
| [126] |
Chang J, Ren Y, Shi Y, Zhu Y M, Ge Y, Hong S M, Jiao L, Lin F M, Peng C H, Mochizuki T, Tani A, Mu Y, Fu C X. An inventory of biogenic volatile organic compounds for a subtropical urban-rural complex. Atmospheric Environment, 2012, 56: 115-123. DOI:10.1016/j.atmosenv.2012.03.053 |
| [127] |
张红星, 韩立建, 任玉芬, 姚余辉, 孙旭, 王效科, 周伟奇, 郑华. 北京城市与西北远郊地表臭氧浓度梯度移动监测研究. 生态学报, 2019, 39(18): 6803-6815. |
| [128] |
Sharkey T D, Wiberley A E, Donohue A R. Isoprene emission from plants: why and how. Annals of Botany, 2008, 101(1): 5-18. |
| [129] |
Tuazon E C, Atkinson R. A product study of the gas-phase reaction of Isoprene with the OH radical in the presence of NOx. International Journal of Chemical Kinetics, 1990, 22(12): 1221-1236. DOI:10.1002/kin.550221202 |
| [130] |
Altimir N, Kolari P, Tuovinen J P, Vesala T, Bäck J, Suni T, Kulmala M, Hari P. Foliage surface ozone deposition: a role for surface moisture?. Biogeosciences, 2006, 3(2): 209-228. DOI:10.5194/bg-3-209-2006 |
| [131] |
Calfapietra C, Fares S, Manes F, Morani A, Sgrigna G, Loreto F. Role of Biogenic Volatile Organic Compounds (BVOC) emitted by urban trees on ozone concentration in cities: a review. Environmental Pollution: Barking, Essex: 1987, 2013, 183: 71-80. |
| [132] |
高峰, 李品, 冯兆忠. 臭氧与干旱对植物复合影响的研究进展. 植物生态学报, 2017, 41(2): 252-268. |
| [133] |
Zeidler J G, Lichtenthaler H K, May H U, Lichtenthaler F W. Is isoprene emitted by plants synthesized via the novel isopentenyl pyrophosphate pathway?. Zeitschrift Für Naturforschung C, 1997, 52(1/2): 15-23. |
| [134] |
Monson R K, Weraduwage S M, Rosenkranz M, Schnitzler J P, Sharkey T D. Leaf isoprene emission as a trait that mediates the growth-defense tradeoff in the face of climate stress. Oecologia, 2021, 197(4): 885-902. DOI:10.1007/s00442-020-04813-7 |
| [135] |
Penuelas J, Llusia J. Linking photorespiration, monoterpenes and thermotolerance in Quercus. New Phytologist, 2002, 155(2): 227-237. DOI:10.1046/j.1469-8137.2002.00457.x |
| [136] |
Barwise Y, Kumar P. Designing vegetation barriers for urban air pollution abatement: a practical review for appropriate plant species selection. Npj Climate and Atmospheric Science, 2020, 3: 12. DOI:10.1038/s41612-020-0115-3 |
| [137] |
Chen Y, Ge Y, Yang G F, Wu Z P, Du Y Y, Mao F, Liu S, Xu R H, Qu Z L, Xu B, Chang J. Inequalities of urban green space area and ecosystem services along urban center-edge gradients. Landscape and Urban Planning, 2022, 217: 104266. DOI:10.1016/j.landurbplan.2021.104266 |
| [138] |
Xiao W. Study on emission of isoprene from major plants living in Taihu Basin. Chinese Bulletin of Botany, 2002(2): 224-230+223. DOI:10.3969/j.issn.1674-3466.2002.02.014 |
| [139] |
王志辉, 张树宇, 陆思华, 白郁华. 北京地区植物VOCs排放速率的测定. 环境科学, 2003, 24(2): 7-12. DOI:10.13227/j.hjkx.2003.02.002 |
| [140] |
Xu Y, Li S J, Yuan X Y, Feng Z Z. Emission characteristics of biogenic volatile compounds (BVOCs) from common greening tree species in Northern China and their correlations with photosynthetic parameters. Huan Jing Ke Xue= Huanjing Kexue, 2020, 41(8): 3518-3526. |
| [141] |
Huang A, Li N, Guenther A, Greenberg J, Baker B, Graessli M, Bai J. Investigation on emission properties of biogenic VOCs of landscape plants in Shenzhen. Environmental Science, 2011, 32(12): 3555-3559. |
| [142] |
Calfapietra C, Angelis P D, Gielen B, Lukac M, Moscatelli M C, Avino G, Lagomarsino A, Polle A, Ceulemans R, Mugnozza G S, Hoosbeek M R, Cotrufo M F. Increased nitrogen-use efficiency of a short-rotation poplar plantation in elevated CO2 concentration. Tree Physiology, 2007, 27(8): 1153-1163. DOI:10.1093/treephys/27.8.1153 |