 2023, Vol. 43
2023, Vol. 43文章信息
- 周怡, 张荷悦, 孙涛, 曹浩冰, 周曾
- ZHOU Yi, ZHANG Heyue, SUN Tao, CAO Haobing, ZHOU Zeng
- 底栖生物影响下的潮滩微地貌演化数值模拟
- Numerical simulation of micro-geomorphic evolution under the influence of benthic organisms in a tidal flat
- 生态学报. 2023, 43(5): 2024-2033
- Acta Ecologica Sinica. 2023, 43(5): 2024-2033
- http://dx.doi.org/10.5846/stxb202202180384
-
文章历史
- 收稿日期: 2022-02-18
- 网络出版日期: 2022-10-21

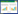




2. 河海大学江苏省海岸海洋资源开发与环境安全重点实验室, 南京 210098;
3. 北京师范大学环境学院, 北京 100875
2. Jiangsu Key Laboratory of Coast Ocean Resources Development and Environment Security, Hohai University, Nanjing 210098, China;
3. College of Environment, Beijing Normal University, Beijing 100875, China
潮滩是陆海交接的过渡地带, 受海陆交互作用, 潮滩上营养物质丰富, 孕育有大量的底栖微藻, 初级生产力非常活跃, 为各类微生物、底栖动物提供了良好的食物基础。潮滩内丰富的生物资源又可通过食物网, 影响整个海岸带生态系统能量流动[1—2]。此外, 一定宽度和坡度的潮滩在周期性水位波动和水沙过程作用下, 通过影响盐沼湿地的淹水条件, 改变盐度在整个潮间带上自然分布梯度, 影响着盐沼植被分布格局和演替进程[3—5]。
潮滩湿地是脆弱的生态敏感区, 也是我国湿地保护的薄弱环节。近些年来, 岸线侵蚀, 环境污染多重扰动下, 潮滩面临的威胁并非局限于单一要素的退化, 生物与物理环境的交互作用使得潮滩出现系统退化问题, 剧烈的泥沙侵蚀同时伴随生物量的丧失[6—7]。但是我国现有潮滩湿地中考虑生物和非生物方法系统融合的生态修复工程较少, 对潮滩湿地生态系统的自然构建和运行的基本规律研究不充分, 限制了系统修复工程的发展, 亟需对底栖生物影响下的潮滩微地貌系统演化机制进行探究。
海岸系统中, 水动力、泥沙运动和地貌演化过程交互作用下可形成丰富迷人的地貌形态, 这种经典的物理地貌动力学方法通过动量守恒、能量守恒以及质量守恒方程等表达形式, 已成功用于模拟和预测河口三角洲, 潮滩等地貌形态[8]。随着研究者对海岸动力地貌过程认识的逐渐深入, 越来越多的研究发现海岸生态系统中生物的动态变化并非只是物理地貌演化的附生现象。生物或生态过程与物理地貌过程相互作用, 直接影响、改变和决定着潮滩景观[9]。因此, 耦合生物与物理动力学过程已逐步成为准确理解和模拟潮滩系统演化的关键。
D′Alpaos等充分考虑盐沼与潮滩水位差, 探究其对滩涂上沉积的泥沙的起动切应力分布的影响, 结合水动力过程模型与地貌形态动力学模型, 在小尺度上模拟了滩涂的发展过程[9]。生物与物理过程间的反馈作用一方面能增加潮滩的稳定性, 但也可能使得潮滩系统发生突变响应, 系统会从一种状态跳跃到另一种状态[10—12]。Marani等通过结合植被生长方程, 同时考虑了高程和底栖微藻对泥沙侵蚀率的影响, 预测了滩涂系统状态转变, 发现滩涂系统在特定海平面高度范围内出现双稳态特性[13]。
生物与物理环境的相互作用会通过空间尺度上的交流影响着系统状态, 从而呈现斑图分布状态, 即自组织微地貌斑图。海岸生态系统中, 退潮状态下的潮滩上出现被底栖微藻覆盖的高丘和积水洼地交替出现的微地貌斑图。微地貌斑图的形成为底栖动物提供了丰富多样的栖息环境, 斑图中底栖微藻生物量和淹水状况具有明显的空间差异性, 其中底栖微藻是底栖动物的重要食物来源, 且食物量的大小和淹水状况均可影响滩涂上底栖腹足类动物的移动行为[14—15]。Weerman等通过野外实验与模型方程结合的方式证明潮滩上的微地貌斑图是底栖微藻-泥沙-水动力自组织行为的结果[16]。然而, 底栖微藻与物理环境自组织形成的异质性栖息环境如可影响底栖动物的种群动态过程, 耦合底栖动物与底栖微藻的生长、死亡反馈过程以及底栖微藻与物理环境自组织过程后, 滩涂上的微地貌斑图系统弹性适应能力将如何变化?这些问题仍不清楚。
微地貌斑图微地貌斑图对黄河三角洲潮滩微地貌斑图的野外调查发现, 微地貌斑图呈现季节性变化, 4月至6月份, 伴随着滩涂上泥螺生物量的增加, 微地貌斑图逐渐减弱, 猜测除受温度等条件的影响, 底栖微藻斑图的变化也受到泥螺活动的影响。本文以黄河三角洲湿地潮滩为研究区, 针对4至6月份滩涂上微地貌斑图变化以及系统对扰动的响应阈值问题, 通过构建二维空间动力模型, 探究底栖生物对微地貌格局演化的作用机制, 分析底栖生物对微地貌系统稳定性的影响, 结果有助于加深对自组织微地貌斑图和地貌演化的机制认识。
1 研究区及方法 1.1 研究区概况黄河三角洲湿地(36°55′—38°12′N, 118°7′—119°10′E)位于山东省东营市东北部(图 1)[17], 与渤海、莱州湾相邻, 处在大陆性季风气候区。由于自然因素和用水增加, 黄河入海口泥沙量持续减少, 影响三角洲潮滩地貌演变。黄河口潮滩总面积达12万hm2, 底质肥沃, 光照充分, 底栖微藻大量繁殖(以硅藻为主), 为底栖动物提供了良好的食物基础[18]。黄河三角洲滩涂区约有大型底栖动物30种, 其中, 软体动物门中的泥螺, 适应能力较强, 作为外来物种在河口滩涂存活后生长繁殖, 已成为黄河口滩涂区域的主要优势物种[19]。
对黄河三角洲潮滩进行野外调查发现, 黄河口滩涂上具有微地貌斑图景观。微地貌斑图呈现沙堆高丘和积水洼地交替出现的空间格局, 内部有大量的底栖生物资源, 其中高丘上覆盖着底栖微藻。黄河口潮滩微地貌斑图呈现季节性变化:春季4月份温度较低时底栖微藻生长迅速, 斑图格局分布明显。6月份温度升高, 底栖微藻生长受到抑制, 规则状斑图减弱, 且在斑图中发现大量的泥螺等底栖动物分布, 次年春季斑图又会重新形成(图 1)。

|
| 图 1 研究区位置及不同季节微地貌斑图 Fig. 1 The location of study area and micro-geomorphic patterns photoed in different seasons |
Weerman等的研究发现, 潮滩微地貌斑图是底栖微藻-泥沙-水动力在不同空间尺度上相互作用形成的结果:底栖微藻分泌胶态碳水化合物, 形成生物膜并附着在滩涂表面, 增加了泥沙粘聚力和起动切应力, 抑制泥沙的侵蚀, 利于泥沙的沉积, 同时为底栖微藻提供了生长发育的环境, 底栖微藻和泥沙交互聚集, 形成沙丘[20—22];同时, 退潮后潮水在高丘旁边的洼地形成积水, 胶状化合物难以形成生物膜, 因此洼地侵蚀严重[16]。这种沙丘上泥沙不断聚集和底栖微藻生长以及洼地中泥沙和微藻被侵蚀使得潮滩上出现微地貌斑图, 高低交替, 模型示意图如图 2。本研究在Weerman等的一维底栖微藻斑图自组织模型基础上, 考虑泥沙和水流在二维空间的扩散, 构建二维底栖微藻自组织模型, 同时采用非线性偏微分方程的形式描述生态物理过程[23]。

|
| 图 2 潮滩微地貌斑图自组织过程及模型结构示意图 Fig. 2 Diagrammatic sketch of self-organization process and model structure of the micro-geomorphic pattern in intertidal mudflats D: 底栖微藻生物量; S: 泥沙高度; W: 水位; t: 时间; Du: 扩散系数; ∇: 梯度算子 |
本研究主要考虑三个关键要素, 包括底栖微藻、泥沙和水位, 如下式(1)、(2)、(3)中分别对其进行描述。采用非线性偏微分方程, 各个方程式包括反应项和扩散项。
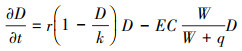
|
(1) |
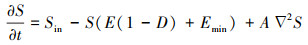
|
(2) |
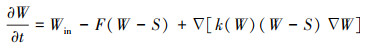
|
(3) |
式(1)描述了微藻的生长和死亡, D (g/m2)是底栖微藻生物量, 第一项描述了底栖微藻的生长过程, r (潮-1)为底栖微藻生长率, k (g/m2)代表底栖微藻环境容纳量;第二项描述底栖微藻死亡, E (cm/潮)为泥沙最大侵蚀率。C (潮-1)是将泥沙侵蚀转化为底栖微藻死亡的常数, W (cm)是水位, q (cm)代表底栖微藻死亡一半时水位高度。
式(2)为泥沙沉积、侵蚀和扩散的过程, S (cm)为泥沙高度, Sin(cm/潮)为泥沙的沉积速率, 中间一项描述泥沙侵蚀, Emin (潮-1)代表泥沙最小侵蚀率。最后一项为重力作用下泥沙从高处扩散到洼地的过程, A (cm2/潮)是泥沙扩散系数, ∇2S描述泥沙扩散。
式(3)中W (cm)代表水位高度, Win (cm/潮)为退潮后的潮滩余水, 中间一项描述排水过程, F (潮-1)代表排水率, 最后一项描述水位坡降引起的水流过程, ∇W为水位梯度引起的水的流动, K(W)代表水深较浅处由渗透引起的水位减少, 如式(4):
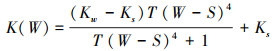
|
(4) |
其中, Kw (cm/潮)代表积水处水渗透率, Ks (cm/潮)代表无积水处渗透率, T为将水深转化为渗透率的无量纲常数。底栖微藻-泥沙-水动力模型所使用参数的释义及取值见表 1。
| 符号 Parameter |
释义 Interpretation |
取值 Value |
单位 Unit |
数据来源 Data source |
| r | 底栖微藻生长率 | 0.8 | 潮-1 | Weerman等[16] |
| k | 底栖微藻环境容纳量 | 1 | g/m2 | Weerman等[16] |
| E | 泥沙侵蚀率 | [0.0012, 0.0032] | cm/潮 | 由1640—2010年间黄河三角洲滩涂高程实测数据估算得出[24] |
| C | 泥沙侵蚀转为底栖微藻死亡的常数 | 500 | 潮-1 | 估测 |
| q | 底栖微藻半饱和常数 | 1 | cm | 估测 |
| Sin | 泥沙沉积率 | 0.02 | cm/潮 | 由二维水动力-泥沙动力过程数值模型计算得到 |
| Emin | 泥沙最小侵蚀率 | 0.0005 | 潮-1 | 估测 |
| A | 重力引起的泥沙扩散速率 | 50 | cm2/潮 | 姚晓等[24] |
| Win | 退潮后的剩余水 | 0.02 | cm/潮 | 估测 |
| F | 排水率 | 0.02 | 潮-1 | 估测 |
| Kw | 水深较高处渗透率 | 10 | cm/潮 | 姚晓等[24] |
| Ks | 水深为零渗透率 | 0.1 | cm/潮 | 姚晓等[24] |
| T | 将水深转化为渗透率常数 | 200 | 姚晓等[24] |
泥螺作为黄河三角洲底栖动物优势种, 通过摄食底栖微藻将会对自组织系统造成较大影响。在潮滩的非均质环境条件下泥螺分布满足关系式(5), 由泥螺生长项与自然死亡项组成。底栖微藻由于受泥螺摄食作用反馈, 增加摄食项如式(6)。
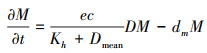
|
(5) |
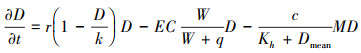
|
(6) |
其中, M代表二维模拟空间中任意位置处泥螺的密度(kg/m2), 泥螺的生长是由对底栖微藻摄食从而转化成的泥螺生物量增加, c代表泥螺对底栖微藻的最大摄食率(kg-1m2潮-1), Kh(g/m2)代表当泥螺对底栖微藻摄食率达到最大值一半时的底栖微藻生物量, Dmean(g/m2)代表模拟范围内底栖微藻的空间平均生物量。式(5)中, 方程式右边第一项代表泥螺生物量的增加, e(kg/g)代表摄食底栖微藻后转化为泥螺生物量的转化效率, 即吸收率;方程式右边第二项代表泥螺的死亡, dm(kgm-2潮)代表泥螺的死亡率, 参数的取值及释义见表 2。
首先对底栖微藻微地貌进行模拟, 初始状态下, 设置泥沙高度为10±2cm的随机分布, 微藻生物量设置为极小值10-7g/m2, 初始水位设置与泥沙高度相同。图 3展示了不同模拟时间T的泥沙高度(S, cm)、水位(W, cm)和底栖微藻生物量(D, g/m2)分布状态。结果显示, 潮滩底栖微藻生长、泥沙与水流在反馈调节作用下形成底栖微藻覆盖的沙丘与积水洼地交替分布的空间规则微地貌景观。当模拟时间为750秒, 微地貌斑图初步形成, 底栖微藻所分泌的胶状物质(EPS)的反馈作用使得泥沙扩散减少, 形成高丘, 同时伴随水位差异增大。1500秒时, 底栖微藻产生的反馈作用持续增强, 致使泥沙高度差不断增加, 底栖微藻生物量增大。4500秒时, 微地貌达到稳定状态。根据泥沙高度、水位和底栖微藻生物量随时间的变化均值, 结果表明稳定状态的泥沙高度差, 水位差, 底栖微藻生物量比初始状态明显增大。

|
| 图 3 滩涂泥沙淤积高度S(cm)、水位W(cm)、底栖微藻生物量D(g/m2)斑图状态随时间变化 Fig. 3 Simulated sediment elevations S (cm), water levels W (cm), and microalgal contents D (g/m2) of the self-organization model under different simulation time |
底栖微藻是潮滩上底栖动物的食物基础, 其生物量的变化会对整个潮滩生态系统的能量流动和稳态响应产生影响[29—30]。为探讨底栖微藻斑图对于滩涂生态系统生物量和泥沙淤积的影响, 将具有完整自组织行为的滩涂系统与不具有完整自组织行为的滩涂系统进行对比。
改变底栖微藻-泥沙-水动力自组织模型中泥沙扩散系数A, 而控制模型中其他关键物理参数保持不变, 最终可获得拥有不同动力学性质的系统:一是具有完整自组织行为系统, 泥沙扩散系数A取值为5 cm2/潮, 此时泥沙扩散能力有限, 水流的扩散能力更强, 泥沙与底栖微藻的正向反馈在潮滩高丘处频繁, 水流与底栖微藻在低洼处发生负反馈调节, 使得系统具有完整自组织行为;二是不具有完整自组织行为系统, 将泥沙扩散系数A设置为50 cm2/潮, 此时泥沙扩散能力较强, 潮滩的微地貌高丘处的泥沙不断向低洼处扩散, 此时各种反馈发生在统一尺度上, 系统不具有完整自组织行为。
模拟结果显示(图 4), 初期(0 < T < 100)底栖微藻生物量迅速增长至0.55 g/m2, 后在水流侵蚀作用下, 底栖微藻减少(100 < T < 400)。此后, 斑图系统中生物自组织行为不断增强, 底栖微藻与泥沙的反馈调节使得生物量不断增加, 同时高丘处泥沙抗侵蚀性能增强, 泥沙不断淤积。而均匀系统中由于泥沙与底栖微藻的正向反馈调节能力有限, 水流侵蚀下微藻生长与泥沙淤积受限。最终系统稳定时, 斑图系统下潮滩生态系统中底栖微藻生物量(0.62 g/m2)和泥沙淤积高度(17.9 cm), 远大于均匀系统下滩涂生态系统中的底栖微藻生物量(0.38 g/m2)和泥沙淤积高度(11.5 cm)。由此证明, 微地貌斑图微地貌的形成有利于泥沙淤积, 同时可增加潮滩生态系统初级生产力。

|
| 图 4 斑图系统和均匀系统底栖微藻生物量D(g/m2)、泥沙淤积高度S(cm)随模拟时间变化 Fig. 4 Changes in microalgae biomass D (g/m2) and sediment elevation S (cm) over simulation time in a patterned system and a uniform system, respectively |
生态系统中生物与环境间的正向反馈关系使得系统对于外界环境扰动呈现非线性复杂的响应。一是存在突变点, 当外界环境扰动使得系统达到突变点时, 系统会从一个状态快速转变为另一个状态;二是系统在某一外界环境扰动范围内存在双稳态, 在这一范围内系统的状态受其历史状态的影响。双稳态的两个端点对应系统的两个突变点, 即系统分叉点:一是在外界胁迫不断增大条件下, 系统崩溃的突变点;二是在外界环境改善条件下, 系统恢复的突变点。双稳态范围的大小可表征系统弹性, 双稳态区域越宽则系统弹性越小, 双稳态区域越窄则系统弹性越大。因此, 探究系统弹性应包括两部分:一是系统在面临外界干扰时的稳定性;二是系统崩溃后, 随外界环境改善, 系统的恢复力。对于生态系统而言, 系统内部动力学行为的变化会改变系统自身弹性和外界扰动下系统发生分岔行为的扰动阈值[31]。对于潮滩微地貌, 环境扰动会使微地貌中斑图不断变化, 并在某一状态发生崩溃。当后续扰动强度恢复至系统崩溃同一水平时, 系统并不会沿着原先演变状态恢复, 若要使系统再次发生突变恢复, 则需要外界扰动强度继续降低。
为明确底栖微藻自组织作用对潮滩微地貌系统弹性的影响, 以泥沙侵蚀率E(cm/潮)作为外界环境干扰因素, 以模型底栖微藻生物量D(g/m2)和泥沙淤积高度S(cm)作为系统状态指标, 模拟潮滩侵蚀率变化下, 微地貌中斑图形态以及底栖微藻生物量与泥沙淤积高度的变化。设置初始生物量为0.3 g/m2泥沙初始高度为10±0.2cm, 侵蚀率从E = 0.0025 cm/潮运行记录模型达到稳定状态后的底栖微藻生物量的空间平均值, 后将侵蚀率逐次增加0.0001 cm/潮, 直至模拟形成的微地貌中不再出现斑图为止。后将初始生物量改为10-7 g/m2, 泥沙高度与水位设置较先前不变, 模型由侵蚀率E=0.0035 cm/潮开始运行, 后逐次减小0.0001 cm/潮, 直至模拟结果中斑图重新出现。
模拟结果显示(图 5), 在底栖微藻-泥沙-水动力自组织行为的作用下, 当侵蚀率在0.0025至0.00338 cm/潮范围内, 底栖微藻生物量随着侵蚀率增大不断减小, 泥沙受到侵蚀淤积高度减小。当E = 0.00338 cm/潮时, 系统达到崩溃突变点, 底栖微藻生物量突变为零, 泥沙淤积高度迅速减小, 斑图消失。若此时减小侵蚀率, 微地貌不会演变至先前状态, 直至侵蚀率减小至E = 0.00318 cm/潮时, 系统达到恢复突变点, 底栖微藻生物量迅速增加且泥沙淤积, 微地貌中斑图出现。由此证明, 潮滩微地貌中存在的自组织行为使潮滩生态系统具有一定抵抗环境因素干扰保持系统稳定和系统失稳后恢复的能力。

|
| 图 5 微地貌系统中底栖微藻生物量D和泥沙淤积高度S对侵蚀率的响应 Fig. 5 Responses of microalgal biomass D and sediment elevation S to erosion rates in a micro-geomorphic system |
泥螺作为养殖物种被人为引入黄河三角洲地区, 由于其适应力和竞争力极强, 且天敌较少, 在逃逸至黄河三角洲潮滩后迅速形成稳定的种群, 后成为黄河三角洲潮滩区域的主要优势种[32—34]。泥螺主要以潮滩底栖硅藻为食, 环境适宜且食物充足使其生长繁殖速度较快, 泥螺种群生物量不容易受到控制。因此, 在黄河三角洲, 相比其他底栖生物, 泥螺对潮滩微地貌中底栖微藻自组织机制将产生更大影响[35]。为明确泥螺入侵后自组织系统对于滩涂生态系统初级生产力与泥沙淤积的影响, 模型的网格大小、模拟范围及其他物理参数均不变, 仅增加泥螺与底栖微藻的摄食反馈作用控制方程。
模型结果显示(图 6), 初期(0 < T < 50)底栖微藻生物量迅速增长至0.51 g/m2, 后在水流侵蚀与泥螺摄食作用下, 底栖微藻减少(50 < T < 400)。此后, 由于泥螺分布在底栖微藻聚集明显的潮滩高丘处, 此处摄食压力更大, 致使高丘处微藻生物量减少, 底栖微藻与泥沙间正向反馈作用减弱, 泥沙受到侵蚀。此时泥螺由于底栖微藻生物量的减少, 其数量也随之减少, 摄食压力降低, 底栖微藻自组织作用使得生物量再次上升, 造成泥沙重新淤积, 在此作用下, 模拟得到的微藻生物量与泥沙淤积高度随时间变化呈现起伏状态。均匀系统中, 由于底栖微藻空间分布对泥螺分布的控制作用, 泥螺也呈均匀分布状态, 系统较快达到平衡状态。

|
| 图 6 考虑泥螺摄食影响条件下, 底栖微藻生物量D(g/m2)、泥沙淤积高度S(cm)随模拟时间变化 Fig. 6 Changes in microalgal biomass D (g/m2) and sediment elevation S (cm) over simulation time considering the consumption of mud snails |
以泥沙侵蚀率E(cm/潮)作为外界环境干扰因素, 以模型底栖微藻生物量D(g/m2)和泥沙淤积高度S(cm)作为系统状态指标, 模拟泥螺入侵的潮滩在侵蚀率变化下, 微地貌中斑图形态以及底栖微藻生物量与泥沙淤积高度的变化。
模拟结果显示(图 7), 当侵蚀率在0.0020至0.00214范围内时, 随着泥沙侵蚀率增加, 模拟得到稳定状态的斑图中底栖微藻斑块面积变小, 同时系统内底栖微藻平均生物量和泥沙淤积高度逐渐减小, 但这一范围内系统出现的变化可逆;当侵蚀率在0.00214至0.00215范围内时, 系统出现双稳态, 存在两个突变点, 一个是当侵蚀率增加达到0.00215时, 底栖微藻生物量和泥沙淤积程度由较强的斑图状态转变为均匀状态, 另一个当侵蚀率减小至0.00214时系统由底栖微藻生物量较低的均匀状态转变为斑图状态, 这一范围内, 系统向哪一种状态发展取决于系统的历史状态, 系统的变化不可逆。

|
| 图 7 考虑泥螺摄食影响条件下, 微地貌系统中底栖微藻生物量D和泥沙淤积高度S对侵蚀率的响应 Fig. 7 Responses of microalgal biomass D and sediment elevation S to erosion rates in a micro-geomorphic system considering the consumption of mud snails |
根据我国近年来的海洋环境质量公报调查结果显示黄河三角洲中潮带, 泥螺在2006—2012年间种群数量及分布范围逐年扩大, 最高生物量为46.4 g/m2 [32]。后经采取多种方法控制泥螺扩散发展, 黄河三角洲一带泥螺生物量于2013年回落至15.84 g/m2, 但仍对黄河三角洲潮滩生物群落造成较大破坏[36]。

|
| 图 8 泥螺不同生物量条件下系统中底栖微藻生物量对侵蚀率变化的响应 Fig. 8 Responses of benthic microalgae concentrations to erosion rates under the influence of different biomass of mud snails |
本研究模拟了泥螺入侵量M0为15 g/m2、20 g/m2、25 g/m2情况下潮滩底栖微藻生物量对侵蚀率变化的响应。模拟结果显示(图 8), 当泥螺生物量为15 g/m2时系统崩溃突变点为0.00215, 且当侵蚀率减少至0.00214时系统恢复;当泥螺生物量为20 g/m2时系统崩溃突变点为0.0019, 且当侵蚀率减少至0.00188时系统恢复;当泥螺生物量为25 g/m2时系统崩溃突变点为0.00165, 且当侵蚀率减少至0.00162时系统恢复。综上所示, 泥螺入侵量的增加减小了潮滩系统的稳定性, 且系统失稳后再恢复的难度增加。
3 结语本文以黄河三角洲潮滩为研究区, 运用模型模拟、稳态分析等方法, 揭示了底栖生物对潮滩微地貌演化的作用机制, 分析得到了在外部干扰下潮滩微地貌斑图动态变化及系统稳定性, 结论如下:
(1) 黄河口潮滩底栖微藻呈不规则斑图分布, 高丘与低洼交替出现。高丘处底栖微藻生物量与泥沙淤积高度较高, 主要由于底栖微藻分泌胶态碳水化合物, 形成生物膜并附着在滩涂表面, 增加了泥沙粘聚力和起动切应力, 抑制泥沙的侵蚀且利于泥沙的沉积, 形成沙丘。同时, 退潮后潮水在洼地处形成积水, 胶状化合物难以形成生物膜, 使得洼地侵蚀加重。底栖微藻与泥沙沉积间存在重要的正向反馈。
(2) 通过对比完整和非完整自组织系统下的底栖微藻生物量和泥沙淤积状况, 结果表明斑图系统下潮滩生态系统中底栖微藻生物量和泥沙淤积高度, 都远大于均匀系统的情况, 微地貌斑图微地貌的形成有利于潮滩生态系统初级生产力增加和泥沙淤积。
(3) 根据分岔分析结果, 当潮滩泥沙侵蚀率发生变化时, 底栖微藻生物量和泥沙淤积高度系统呈非线性关系, 当侵蚀率增大至0.00338 cm/潮时斑图消失, 底栖微藻迅速消失, 泥沙高程呈现均匀状态。崩溃后的潮滩系统可在侵蚀率重新减小至0.00318 cm/潮时再次恢复至先前水平。黄河口泥螺入侵使得系统抵抗外界环境扰动能力减小, 且当泥螺入侵量增加时, 系统崩溃后恢复能力降低, 这是由于泥螺对底栖微藻的摄食压力增大, 抑制了底栖微藻与泥沙沉积间的互馈过程。
本文的结果有助于对黄河三角洲潮滩生态系统微地貌格局演化机制的深入认识, 为海岸带保护修复的国家需求提供科学依据。
| [1] |
陈勇. 海岸带滩涂资源遥感应用研究进展. 遥感技术与应用, 2012, 27(2): 296-304. |
| [2] |
李少朋, 邢前国. 潮滩底栖微藻生物量垂直变化对其遥感反演模式的影响. 生态科学, 2014, 33(6): 1155-1159. DOI:10.14108/j.cnki.1008-8873.2014.06.019 |
| [3] |
Lawrenz E, Smith E M, Richardson T L. Spectral irradiance, phytoplankton community composition and primary productivity in a salt marsh estuary, north inlet, South Carolina, USA. Estuaries and Coasts, 2013, 36(2): 347-364. DOI:10.1007/s12237-012-9567-y |
| [4] |
Silliman B R, Van De Koppel J, Bertness M D, Stanton L E, Mendelssohn I A. Drought, snails, and large-scale die-off of southern U.S. salt marshes. Science, 2005, 310(5755): 1803-1806. DOI:10.1126/science.1118229 |
| [5] |
Statham P J. Nutrients in estuaries-An overview and the potential impacts of climate change. Science of the Total Environment, 2012, 434: 213-227. DOI:10.1016/j.scitotenv.2011.09.088 |
| [6] |
陈玮彤, 张东, 崔丹丹, 吕林, 谢伟军, 施顺杰, 侯泽宇. 基于遥感的江苏省大陆岸线岸滩时空演变. 地理学报, 2018, 73(7): 1365-1380. |
| [7] |
毋亭, 侯西勇. 海岸线变化研究综述. 生态学报, 2016, 36(4): 1170-1182. |
| [8] |
Giosan L, Syvitski J, Constantinescu S, Day J. Climate change: protect the world's deltas. Nature, 2014, 516(7529): 31-33. DOI:10.1038/516031a |
| [9] |
D'Alpaos A, Lanzoni S, Marani M, Fagherazzi S, Rinaldo A. Tidal network ontogeny: Channel initiation and early development. Journal of Geophysical Research: Earth Surface, 2005, 110(F2): F02001. |
| [10] |
Rietkerk M, Dekker S C, De Ruiter P C, Van De Koppel J. Self-organized patchiness and catastrophic shifts in ecosystems. Science, 2004, 305(5692): 1926-1929. DOI:10.1126/science.1101867 |
| [11] |
Rietkerk M, Van de Koppel J. Regular pattern formation in real ecosystems. Trends in Ecology & Evolution, 2008, 23(3): 169-175. |
| [12] |
Suding K N, Hobbs R J. Threshold models in restoration and conservation: a developing framework. Trends in Ecology & Evolution, 2009, 24(5): 271-279. |
| [13] |
Marani M, D'Alpaos A, Lanzoni S, Carniello L, Rinaldo A. Biologically-controlled multiple equilibria of tidal landforms and the fate of the Venice lagoon. Geophysical Research Letters, 2007, 34(11): L11402. DOI:10.1029/2007GL030178 |
| [14] |
Xiao R, Zheng Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnology Advances, 2016, 34(7): 1225-1244. DOI:10.1016/j.biotechadv.2016.08.004 |
| [15] |
De Brouwer J F C, Bjelic S, De Deckere E M G T, Stal L J. Interplay between biology and sedimentology in a mudflat (Biezelingse Ham, Westerschelde, The Netherlands). Continental Shelf Research, 2000, 20(10/11): 1159-1177. |
| [16] |
Weerman E J, Van De Koppel J, Eppinga M B, Montserrat F, LIU Q X, Herman P M J. Spatial self-organization on intertidal mudflats through biophysical stress divergence. The American Naturalist, 2010, 176(1): E15-E32. DOI:10.1086/652991 |
| [17] |
陈利顶, 傅伯杰. 黄河三角洲地区人类活动对景观结构的影响分析——以山东省东营市为例. 生态学报, 1996, 16(4): 337-344. |
| [18] |
刘陆. 黄河口潮间带泥滩环境微生物群落特征研究[D]. 青岛: 中国海洋大学, 2014.
|
| [19] |
夏江宝, 李传荣, 许景伟, 郑莉, 刘立杰. 黄河三角洲滩涂区大型底栖动物群落数量特征. 生态环境学报, 2009, 18(4): 1368-1373. DOI:10.3969/j.issn.1674-5906.2009.04.029 |
| [20] |
Callaway R M, Brooker R W, Choler P, Kikvidze Z, Lortie C J, Michalet R, Paolini L, Pugnaire F I, Newingham B, Aschehoug E T, Armas C, Kikodze D, Cook B J. Positive interactions among alpine plants increase with stress. Nature, 2002, 417(6891): 844-848. DOI:10.1038/nature00812 |
| [21] |
Lumborg U, Andersen T J, Pejrup M. The effect of Hydrobia ulvae and microphytobenthos on cohesive sediment dynamics on an intertidal mudflat described by means of numerical modelling. Estuarine, Coastal and Shelf Science, 2006, 68(1/2): 208-220. |
| [22] |
龚政, 葛冉, 冯骞, 魏佳欣, 苏敏, 靳闯, 罗景阳. 泥沙颗粒间黏结力作用及其对泥沙起动影响研究进展. 水科学进展, 2021, 32(5): 801-812. |
| [23] |
Zhang H Y, Sun T, Yang W. Short-term environmental flow assessment of a functional estuarine tidal flat ecosystem: a nonlinear ecological response to flow alteration. Water Resources Research, 2020, 56(12): e2020WR027084. |
| [24] |
姚晓. 黄河三角洲南部潮间带底栖生产力研究[D]. 青岛: 中国海洋大学, 2010.
|
| [25] |
Møhlenberg F, Riisgård H U. Filtration rate, using a new indirect technique, in thirteen species of suspension-feeding bivalves. Marine Biology, 1979, 54(2): 143-147. DOI:10.1007/BF00386593 |
| [26] |
Winter J E. The filtration rate of Mytilus edulis and its dependence on algal concentration, measured by a continuous automatic recording apparatus. Marine Biology, 1973, 22(4): 317-328. DOI:10.1007/BF00391388 |
| [27] |
吴文广. 莱州湾泥螺(Bullacta exarata)生理、生态学特性及生态风险评价[D]. 上海: 上海海洋大学, 2013.
|
| [28] |
尤仲杰, 王一农, 丁伟, 徐海军. 几种环境因子对不同发育阶段的泥螺Bullacta exarata的影响. 浙江水产学院学报, 1994, 13(2): 79-85. |
| [29] |
Underwood G J C, Kromkamp J. Primary production by phytoplankton and microphytobenthos in estuaries. Advances in Ecological Research, 1999, 29: 93-153. |
| [30] |
Kirwan M L, Megonigal J P. Tidal wetland stability in the face of human impacts and sea-level rise. Nature, 2013, 504(7478): 53-60. DOI:10.1038/nature12856 |
| [31] |
赵东升, 张雪梅. 生态系统多稳态研究进展. 生态学报, 2021, 41(16): 6314-6328. |
| [32] |
赵文溪, 宋静静, 于超勇, 刘莹, 刘元文, 官曙光, 于道德. 黄河三角洲区域泥螺入侵与扩散研究进展. 海洋开发与管理, 2017, 34(S2): 142-147. |
| [33] |
胡思敏. 试论我国沿海滩涂资源的自然条件特征及利用途径. 土壤通报, 1983(2): 4-8. |
| [34] |
张晓龙, 李萍, 刘乐军, 李培英, 单凯. 黄河三角洲湿地生物多样性及其保护. 海岸工程, 2009, 28(3): 33-39. |
| [35] |
Zhang H Y, Sun T, Cao H B, Zhang Y, Yang W, Shao D D, Cui B S, Zhou Z. Movement of mud snails affects population dynamics, primary production and landscape heterogeneity in tidal flat ecosystems. Landscape Ecology, 2021, 36(12): 3493-3506. |
| [36] |
蒋万钊. 黄河三角洲泥螺分布、生长规律的演变及控制技术研究. 河北渔业, 2014(4): 11-12. |