 2023, Vol. 43
2023, Vol. 43文章信息
- 霍兵兵, 孙哲明, 欧文慧, 毛鸿志, 胡傲, 杨予静, 李中强
- HUO Bingbing, SUN Zheming, OU Wenhui, MAO Hongzhi, HU Ao, YANG Yujing, LI Zhongqiang
- 环境筛选和扩散限制对长江流域湖北段湿地植物群落构建的共同影响
- Environmental filtering and dispersal limitation jointly affect wetland plant community assembly in Hubei section of the Yangtze River Basin
- 生态学报. 2023, 43(5): 1804-1811
- Acta Ecologica Sinica. 2023, 43(5): 1804-1811
- http://dx.doi.org/10.5846/stxb202201110110
-
文章历史
- 收稿日期: 2022-01-11
- 网络出版日期: 2022-10-21





2. 武汉市桥梁工程有限公司, 武汉 430074;
3. 湖北省林业科学研究院, 武汉 430075
2. Wuhan Bridge Engineering Co., Ltd., Wuhan 430074, China;
3. Hubei Academy of Forestry, Wuhan 430075, China
植物群落构建机制研究对于阐明物种共存和维持物种多样性至关重要, 因此该方面研究一直是群落生态学研究的重要科学问题之一[1—2], 并取得许多重要研究成果[3—4]。在群落构建机制众多假说中, 基于物种的生态位理论和中性理论是目前解释植物群落构建机制的两个主要理论[5—6]。生态位理论认为群落构建的驱动力是环境因子筛选和限制相似性[7—8], 中性理论则认为距离效应所代表的扩散限制决定了群落中物种的组成[9]。近年来, 关于植物群落构建机制的研究更倾向于将生态位理论与中性理论进行整合, 综合多种方法、多影响因素来探讨群落的构建机制[2, 10—12]。越来越多的研究表明环境筛选和扩散限制并非单独影响群落构建, 多数情况下, 两者共同作用于群落构建, 区别仅在于相对贡献的大小[4]。
目前, 关于植物群落构建机制的研究主要集中在草原、森林等陆生植物群落方面[13—15], 且发现不同植物群落类型环境筛选和扩散限制对群落构建的贡献率不同[13—16], 因此需要加强不同生态系统类型中不同群落类型的构建机制研究, 来进一步阐明植物群落的构建机制[10]。相对陆生植物群落而言, 有关湿地植物群落构建机制的研究较少[17], 且相关研究发现由于陆生和湿生环境在气候、光照等环境因子方面的巨大差异, 导致湿生植物群落构建机制不同于陆生植物[18], 因此需加强不同湿地类型中不同湿地植物群落构建机制研究来阐明湿地植物群落的构建机制。
湖北省位于长江流域, 是我国湿地资源最丰富的地区之一, 为全球典型的江湖淤塞淡水湿地区[19], 其湿地类型丰富, 有河流湿地、湖泊湿地、沼泽湿地和库塘湿地等[20]。区域内丰富的湿地资源为湿地植物群落构建机制研究提供了良好条件。β多样性是指生境间的物种多样性, 可以较为直观地描述群落物种组成差异, 是研究群落构建机制的重要指标[21—23]。本研究以湖北省不同类型湿地内主要湿地植物为研究对象, 利用β多样性分析其群落构建以期明确湖北省湿地植物群落构建机制, 为该区域湿地生态系统管理和受损湿地生态恢复与重建提供重要理论依据。
1 材料与方法 1.1 研究区概况长江流域湖北段(29°01′53″—33°6′47″N, 108°21′42″—116°07′50″E) 多属典型的亚热带季风性湿润气候。年降水量约为1000—1500 mm, 光热充足, 降水丰沛, 雨热同期[24]。长江流域湖北段地势略呈由西北向东南倾斜的趋势, 长江通过三峡由西进入湖北, 在黄冈市黄梅县流出;宜昌以西流域属长江上游, 地形多以山地为主;宜昌以东流域属长江中游, 地形以丘陵、平原为主[24—25]。
根据2012年全国第二次湿地资源调查结果, 长江流域湖北段8 hm2以上的湿地总面积为144.50万hm2, 占湖北省国土面积的7.8%[20]。根据《全国湿地资源调查与监测技术规程》分类标准, 湖北省湿地主要分为4大类:河流型湿地、湖泊型湿地、库塘型湿地、沼泽与沼泽化草甸型湿地[20]。
1.2 调查方法本研究于2020年8—10月在长江流域湖北段内选择8个不同类型的代表性湿地进行湿地植物群落调查, 8个湿地分别为神农架大九湖湿地、湖北远安沮河湿地、湖北襄阳汉江湿地、湖北松滋洈水湿地、湖北公安崇湖湿地、洪湖湿地、湖北武汉府河湿地、湖北武汉东湖湿地, 包括河流、湖泊、沼泽和库塘4种湿地类型[20](图 1)。植物群落调查根据实地情况所选湿地面积的大小和生境的复杂程度来确定样点的数量, 共计调查108个样点:其中神农架大九湖湿地设置17个样点、湖北远安沮河湿地设置38个样点、湖北襄阳汉江湿地设置8个样点、湖北松滋洈水湿地设置9个样点、湖北公安崇湖湿地设置10个样点、洪湖湿地设置6个样点、湖北武汉府河湿地设置10个样点、湖北武汉东湖湿地设置10个样点。

|
| 图 1 长江流域湖北段8个采样区位置分布图 Fig. 1 Map showing the 8 sampling areas and its location in Hubei section of the Yangtze River Basin |
本次研究湿地植物包括生活在水中的水生植物及生长在含水量达100%的土壤上的湿生植物。根据植物群落清查的主要内容、方法和技术规范[26]方法对样点湿地植物群落进行调查, 其中沉水植物群落、浮叶植物群落样方大小设置为1 m×1 m, 高大挺水植物群落为2 m×2 m;植被稀疏生境的湿地植物群落为5 m×5 m或10 m×10 m。调查记录植物物种名、多度、盖度等。同时利用手持全球定位系统(GPS)(Garmin e Trex 10)读取样方地理坐标(经纬度与海拔)并进行记录。使用照度计测量水体表面和水下光强, 并记录水深, 根据公式计算其光衰减系数[27]。

|
(1) |
式中, 用Kd表示水体光衰减系数, Z为所测量水体深度, E(Z)为水深Z处的辐照度, E0为水体表面的辐照度。
使用多功能水质检测仪(YSI Pro2030)读取部分水体溶解氧、电导率和温度等理化指标并记录。现场采集水样100 mL进行避光冷冻保存, 调查结束后带回实验室利用紫外分光度计(DR6000)检测氮磷等其它指标。水体中氨氮(NH4+-N)的测量使用纳氏试剂光度法;硝态氮(NO3-N)的含量使用紫外分光光度法测量;总氮(TN)的含量使用碱性过硫酸钾消解-紫外分光光度法测定;总磷(TP)的测定使用碱性过硫酸钾消解-钼酸铵分光光度法[28]。气候数据来源于全球气候数据网站“WorldClim” (Http://www.worldclim.org)。
植物群落命名方法采用优势度原则[29], 优势种的确定采用重要值方法。相同层次的优势种之间用“+”连接, 不同层次的优势种之间用“-”连接。
1.3 数据分析方法植物群落命名采用重要值方法[26]:

|
根据研究区域所调查样方内的物种重要值命名植物群落, 采用聚类分析方法将调查区域内的植物群落总结得出主要湿地植物群丛类型。
采用非对称的距离计算群落间物种β多样性指数即物种相异性指数[30], 其公式如下:
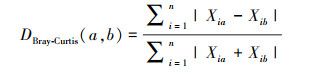
|
(2) |
式中DBray-Curtis(a, b)指第a个和第b个样方间的物种相异性指数, n为物种数, Xia为第a个样方中第i个物种的多度, Xib为第b个样方的第i种的多度。使用多元非参数方差分析(PERMANOVA)检验不同样地湿地植物群落β多样性(Bray-Curtis相异性指数)差异程度和显著性。
地理距离和环境因子距离都基于欧式距离进行计算, 其中地理距离计算公式如下:
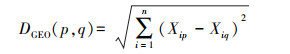
|
(3) |
式中DGEO(p, q)为第p个和第q个样方间地理距离系数。n为地理坐标维度, Xip为第p个样方的第i维地理坐标数据, Xiq为第q个样方的第i维地理坐标数据。
环境因子距离计算公式为:
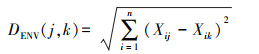
|
(4) |
式中DENV(j, k)为第j个和第k个样方间环境因子欧式距离系数。其中n为环境因子数, Xij为第j个样方的第i维环境因子数据, Xik为第k个样方的第i维环境因子数据。
将采样点海拔、总磷、硝态氮、氨态氮、溶解氧、电导率、流速、光衰减系数、年均温、最暖季均温、最冷季均温、年降雨量共同构成环境因子矩阵。先对环境因子执行Z-Score标准化, 以消除量纲差异带来的影响;用物种相异性指数方法计算β多样性指数。利用物种相异性指数、地理距离和环境因子距离构建物种相异性矩阵-地理距离矩阵及物种相异性矩阵-环境因子距离矩阵。对环境因子变量进行冗余度评估, 手动去除部分相关性高的变量, 保留相关性低的变量, 采用相关性检验(Mantel test)分别分析群落物种相异性矩阵与各环境因子、地理距离矩阵和环境距离矩阵的相关性;通过基于相异度多元回归分析(MRM)研究地理距离和环境距离对群落物种变异性指数的影响的贡献率, 并利用非参数置换检验其显著性。以上操作在R 4.1.1软件中利用vegan、Hmisc、ecodist等程序包中的相关函数实现, 并用ggplog2程序包制作散点图[31]。
2 结果与分析 2.1 研究区内主要湿地植物群丛类型及不同湿地β多样性指数格局在湖北省内8个不同类型的代表性湿地中共调查到湿地植物103科259属338种, 根据中国植被分类法, 结合聚类分析与数量分类法, 分析表明调查区域的植物群落可划分为12个群丛, 分别为菹草群丛(Potamogeton crispus Association)、睡菜群丛(Menyanthes trifoliata Association)、莲群丛(Nelumbo nucifera Association)、芦苇群丛(Phragmites australis Association)、苦草群丛(Vallisneria natans Association)、大茨藻群丛(Najas marina Association)、狐尾藻+黑藻群丛-木贼(Myriophyllum spicatum+Hydrilla verticillate.-Equisetum hyemale Association)、矛叶荩草群丛(Arthraxon lanceolatus Association)、香茶菜群丛(Isodon amethystoides Association)、沼生水马齿群丛(Callitriche palustris Association)、满江红群丛(Azolla pinnata subsp. asiatica Association)、竹叶眼子菜群丛(Potamogeton wrightii Association)。
Bray-Curtis相异性指数分析表明研究区内8个不同类型湿地内部植物群落Bray-Curtis相异性指数的中位数从大到小分别为:湖北远安沮河湿地(0.955)>湖北公安崇湖湿地(0.943)>湖北松滋洈水湿地(0.933)>神农架大九湖湿地(0.908)>湖北武汉府河湿地(0.892)>湖北武汉东湖湿地(0.846)>洪湖湿地(0.783)>湖北襄阳汉江湿地(0.698), 见图 2。PERMANOVA分析表明8个不同类型湿地内植物群落物种β多样性指数存在显著差异(P < 0.001)。

|
| 图 2 长江流域湖北段不同类型湿地植物群落间物种相异性指数差异 Fig. 2 Differences in Bray-Curtis dissimilarity index among different types of wetland plant communities in Hubei section of the Yangtze River Basin 1:湖北公安崇湖湿地;2:湖北武汉东湖湿地;3:神农架大九湖湿地;4:湖北武汉府河湿地;5:洪湖湿地;6:湖北松滋洈水湿地;7:湖北远安沮河湿地;8:湖北襄阳汉江湿地 |
研究区域各环境因子与湿地植物群落β多样性指数相关性分析表明在所检测的环境因子中, 除流速与水体中总磷含量外, 其余各环境因子均与群落中物种的相异性即β多样性指数呈显著正相关关系, 其中海拔、溶解氧、电导率、光衰减系数、年均温、最暖季均温、最冷季均温和年降雨量呈极显著正相关(表 1)。
| 环境因子 Environmental factors |
相关性Correlation | 贡献率Contribution rate | |||
| r | P | r2 | P | ||
| 海拔Altitude | 0.44 | 0.0001 | — | — | |
| 总氮Total Nitrogen | 0.09 | 0.005 | 0.008 | 0.01 | |
| 总磷Total Phosphorus | 0.06 | 0.10 | 0.003 | 0.21 | |
| 硝态氮Potassium Nitrate | 0.08 | 0.03 | 0.004 | 0.10 | |
| 氨态氮Ammonia Nitrogen | 0.06 | 0.07 | 0.003 | 0.16 | |
| 溶解氧Dissolved Oxygen | 0.12 | 0.0001 | 0.014 | 0.0001 | |
| 电导率Conductivity | 0.19 | 0.0001 | 0.030 | 0.0001 | |
| 流速Flow Velocity | -0.07 | 0.96 | 0.006 | 0.71 | |
| 光衰减系数Attenuation Coefficients | 0.14 | 0.0001 | 0.017 | 0.0003 | |
| 年均温Annual Average Temperature | 0.43 | 0.0001 | 0.184 | 0.0001 | |
| 最暖季均温Mean Temperature in the Warmest Season | 0.43 | 0.0001 | — | — | |
| 最冷季均温Mean Temperature in the Coldest Season | 0.42 | 0.0001 | — | — | |
| 年降雨量Annual Precipitation | 0.22 | 0.0001 | 0.043 | 0.0001 | |
| P>0.05不显著;P < 0.05显著;P < 0.001极显著 | |||||
Mantel test结果表明研究区内群落间物种β多样性指数与地理距离和环境距离均呈显著正相关关系(P < 0.01)(图 3、图 4), 但从相关系数来看, 环境距离与β多样性的相关性更高。

|
| 图 3 长江流域湖北段湿地植物群落物种相异性指数与环境距离的相关性 Fig. 3 Correlation between species dissimilarity index of wetland vegetation community and environmental distance in Hubei section of the Yangtze River Basin |

|
| 图 4 长江流域湖北段湿地植物群落物种相异性指数与地理距离的相关性 Fig. 4 Correlation between species dissimilarity index of wetland vegetation community and geographic distance in Hubei section of the Yangtze River Basin |
MRM分析结果显示, 环境距离和地理距离共同解释了研究区内群落物种变异性指数的54.72%。其中, 环境距离独立解释率为22.03%, 地理距离独立解释率为9.98%, 二者联合解释率为22.71%, 未解释率为45.28%(图 5)。共线性分析去除了海拔、最暖季均温和最冷季均温之后, 环境因子中对湿地植物群落构建解释率最高的为年均温, 解释率为18.40%(表 1)。

|
| 图 5 环境距离和与地理距离对长江流域湖北段湿地植物β多样性指数影响的相对贡献率 Fig. 5 The relative contribution rate of environmental distance and geographical distance on the β-diversity index of wetland plants in Hubei section of the Yangtze River Basin |
β多样性表征不同群落之间物种组成上的相异性, 当群落所处的生境异质性越大时, 群落间的结构和组成差异也会越大, 即生境过滤作用[32]。同样, 扩散限制作用会限制种子的扩散距离, 随着两个群落间距离的增加, 群落间的结构和组成差异也会增加, 进而表现出较高的β多样性, 即群落的距离衰减效应[30]。与以往研究结果一致[18], 本研究发现长江流域湖北段不同湿地植物群落间物种β多样性指数与环境距离均呈显著正相关关系, 表明环境因子差异越大, 该区域湿地植物群落物种组成差异越大。环境变化影响着物种在群落中是否经历了环境过滤的筛选过程, 经过环境条件和生物的相互作用, 具有特定性状并符合各环境筛的特定物种共同构建群落[33]。本研究发现与该区域的湿地植物群落物种组成差异最相关的环境因子为年均温等气候因子, 这与长江中下游岸带湿地植物群落构建机制研究结果一致[17], 其可能解释是湿地植物为一年生或多年生草本植物, 受气温影响较大, 大多数湿地植物冬季地上部分会死亡, 通过采用种子和地下繁殖体方式渡过不良生境。虽然本研究表明年均温是所检测的因子中对湿地植物群落构建解释率最高的环境因子, 但解释率也仅为18.40%, 解释率并不高;因此需要进一步研究其他因素如时间尺度、人类干扰等在该区域湿地植物群落构建中所起的作用。此外, 本研究也发现, 即使在同样一个调查区域内的湿地植物群落, 其物种相异性指数也很高, 且物种相异性指数与水中的总氮、电导率、光衰减系数和溶解氧呈显著相关, 这也进一步表明环境筛选过程对湿地植物群落建成影响巨大。其原因是湿地植物为适应水位变化而形成四种生活型, 同一湿地范围内不同湿地植物群落是经历了水位、水体透明度等环境筛选过程而形成的。同样, 与早期关于湿地植物β多样性构建机制研究结果一致[32], 本研究区内湿地植物群落物种组成差异也随地理距离的增加而增大, 许多研究表明扩散限制在物种周转与群落构建中都扮演了重要的角色[11, 34], 其原因是随着样地间距离的增加, 扩散限制对繁殖体的传播束缚能力变大[33];尽管湿生植物繁殖方式多样, 常以种子、断枝、地下根茎、芽等方式进行繁衍和种群扩散[35], 但由于受自身生物学特点(寿命、萌发率等)、捕食、环境因子及江湖阻隔等因素影响, 因而表现为空间距离往往限制着物种的扩散而呈现群落聚集的相似性随距离衰减关系[36]。
近年来关于生态位和中性理论的验证研究已经取得了显著的成果, 但对于局域植物群落构建机制的认识仍存在很大争议[12]。越多的研究表明基于生态位的环境筛选和基于中性理论的扩散限制等多个过程可能共同影响着植物群落的构建, 区别仅在于相对贡献的大小不同[11], 且具有尺度依赖性[12]。与早期研究结果一样[17], 本研究亦证明长江流域湖北段湿地植物群落构建是基于生态位理论的环境筛选和基于中性理论的扩散限制共同作用的结果, 且环境距离对研究区内湿地植物群落构建作用更大, 这主要由于不同水生或湿生植物物种对环境如水位、透明度和基质等要求和适应性不同[37]。同样本研究也发现, 环境距离和地理距离联合解释率对该研究区域湿地植物群落构建解释率较高, 也进一步表明该区域湿地植物群落构建除了受到环境筛选的影响外, 物种的扩散在一定程度上还受限于空间距离[17]。事实上, 扩散限制不仅仅取决于地理距离的隔绝, 还受制于物种本身的性状, 如个体的扩散能力等[38], 同时也可能受到长江流域枯水期、丰水期的水位变化影响[39]。尽管本研究以不同湿地类型为研究对象, 且由于江湖阻隔, 彼此间水系连通较少, 但本研究却表明扩散限制对长江流域湖北段湿地植物群落构建贡献相对较小, 其原因为:(1)许多湿生植物特别是水生植物多为广布种, 具有无性繁殖发达、种子等繁殖体容易扩散和湿地生境的相对同质性[40];(2)与研究尺度相关, 因为尺度依赖性是影响植物群落构建一个重要的影响因子[12]。因此, 后续研究应同时考虑空间和时间尺度下植物群落的构建过程[10], 以便完整地理解该区域湿地植物群落的构建机制。
4 结论本研究表明环境筛选和扩散限制共同影响长江流域湖北段湿地植物群落构建, 且环境筛选作用贡献更大。表明在对该区域受损湿地生态系统恢复与重建过程中, 特别需要重视环境因子的作用, 有针对性地改善湿地植物群落的生境从而促进湿地植物群落的恢复与重建。同时不能忽视扩散限制作用的影响, 如实施适当的人为引种等措施促进群落物种扩散以恢复受损湿地生物多样性。
| [1] |
Liu M X, Ma Y B, Wang X Y, Xu L. Plant community assembly mechanisms of a subalpine meadow community along different successional time. Rangeland Ecology & Management, 2021, 77: 118-125. |
| [2] |
Zhu J T, Zhang Y J, Yang X, Chen N, Li S P, Wang P D, Jiang L. Warming alters plant phylogenetic and functional community structure. Journal of Ecology, 2020, 108(6): 2406-2415. DOI:10.1111/1365-2745.13448 |
| [3] |
Sabatini F M, Jiménez-Alfaro B, Burrascano S, Lora A, Chytry' M. Beta-diversity of central European forests decreases along an elevational gradient due to the variation in local community assembly processes. Ecography, 2018, 41(6): 1038-1048. DOI:10.1111/ecog.02809 |
| [4] |
Gravel D, Canham C D, Beaudet M, Messier C. Reconciling niche and neutrality: the continuum hypothesis. Ecology Letters, 2006, 9(4): 399-409. DOI:10.1111/j.1461-0248.2006.00884.x |
| [5] |
Rosindell J, Hubbell S P, Etienne R S. The unified neutral theory of biodiversity and biogeography at age ten. Trends in Ecology & Evolution, 2011, 26(7): 340-348. |
| [6] |
Vellend M. Conceptual synthesis in community ecology. The Quarterly Review of Biology, 2010, 85(2): 183-206. DOI:10.1086/652373 |
| [7] |
Chase J M. Ecological niche theory. The theory of ecology, 2011, 93-107. |
| [8] |
Webb C O, Ackerly D D, McPeek M A, Donoghue M J. Phylogenies and community ecology. Annual Review of Ecology and Systematics, 2002, 33: 475-505. DOI:10.1146/annurev.ecolsys.33.010802.150448 |
| [9] |
Koo K A, Park S U. A review of ecological niche theory from the early 1900s to the Present1a. Korean Journal of Environment and Ecology, 2021, 35(4): 316-335. DOI:10.13047/KJEE.2021.35.4.316 |
| [10] |
Linares-Palomino R, Kessler M. The role of dispersal ability, climate and spatial separation in shaping biogeographical patterns of phylogenetically distant plant groups in seasonally dry Andean forests of Bolivia. Journal of Biogeography, 2009, 36(2): 280-290. DOI:10.1111/j.1365-2699.2008.02028.x |
| [11] |
王丹, 王孝安, 郭华, 王世雄, 郑维娜, 刘史力. 环境和扩散对草地群落构建的影响. 生态学报, 2013, 33(14): 4409-4415. |
| [12] |
柴永福, 岳明. 植物群落构建机制研究进展. 生态学报, 2016, 36(15): 4557-4572. |
| [13] |
Jones M M, Tuomisto H, Borcard D, Legendre P, Clark D B, Olivas P C. Explaining variation in tropical plant community composition: influence of environmental and spatial data quality. Oecologia, 2008, 155(3): 593-604. DOI:10.1007/s00442-007-0923-8 |
| [14] |
许金石, 柴永福, 刘晓, 岳明, 郭垚鑫, 康慕谊, 刘全儒, 郑成洋, 吉成均, 闫明, 张峰, 高贤明, 王仁卿, 石福臣, 张钦弟, 王茂. 华北区域环境梯度上阔叶林构建模式及分布成因. 植物生态学报, 2019, 43(9): 732-741. |
| [15] |
赵鸣飞, 王国义, 邢开雄, 王宇航, 薛峰, 康慕谊, 罗开. 秦岭西部森林群落相似性递减格局及其影响因素. 生物多样性, 2017, 25(1): 3-10. |
| [16] |
施晶晶, 赵鸣飞, 王宇航, 薛峰, 康慕谊, 江源. 黄土高原腹地人工林下草本层群落构建机制. 植物生态学报, 2019, 43(9): 834-842. |
| [17] |
欧阳键. 湿生植物的群落构建机制研究——以长江中下游岸带和西藏的高原湖滨带为例[D]. 拉萨: 西藏大学, 2020.
|
| [18] |
欧阳键, 刘贵华, 周雯. 长江中下游岸带(宜昌-铜陵段)湿生植物的群落构建机制研究. 植物科学学报, 2020, 38(6): 751-761. |
| [19] |
史玉虎, 吴翠, 赵金萍. 长江中下游湿地开发模式探讨. 湖北林业科技, 2010, 39(5): 43-44, 51. |
| [20] |
石道良, 朱兆泉, 肖宇, 蒲云海. 湖北湿地资源特点及保护管理对策研究. 湖北林业科技, 2004, 33(1): 38-40. |
| [21] |
Mori A S, Isbell F, Seidl R. Β-diversity, community assembly, and ecosystem functioning. Trends in Ecology & Evolution, 2018, 33(7): 549-564. |
| [22] |
Rodriguez-Artigas S M, Ballester R, Corronca J A. Factors that influence the beta-diversity of spider communities in northwestern Argentinean Grasslands. PeerJ, 2016, 4: e1946. |
| [23] |
Marion Z H, Fordyce J A, Fitzpatrick B M. Pairwise beta diversity resolves an underappreciated source of confusion in calculating species turnover. Ecology, 2017, 98(4): 933-939. |
| [24] |
潘磊, 唐万鹏, 史玉虎, 袁传武. 长江中游(湖北段)江岸带立地分类与评价. 湖北林业科技, 2002, 31(3): 12-17. |
| [25] |
蒲云海, 刘胜祥, 朱兆泉, 苗文杰, 郑联合. 湖北省湿地自然保护区监测手册. 武汉: 湖北科学技术出版社, 2020.
|
| [26] |
方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [27] |
张运林, 秦伯强, 陈伟民, 杨顶田. 太湖水体光学衰减系数的特征及参数化. 海洋与湖沼, 2004, 35(3): 209-213. |
| [28] |
张金美. 鄱阳湖区典型真水生植物组织中碳、氮同位素特性研究[D]. 南昌: 南昌大学, 2016.
|
| [29] |
田玉清, 石道良, 张淑倩, 龚旭昇, 汪正祥, 李中强. 河西走廊水生植物多样性格局、群落特征及影响因素. 生态学报, 2020, 40(1): 202-212. |
| [30] |
刘晓红, 李校, 彭志杰. 生物多样性计算方法的探讨. 河北林果研究, 2008, 23(2): 166-168. |
| [31] |
孙哲明. 基于β多样性的长江中下游湿地植物的群落构建机制[D]. 武汉: 湖北大学, 2021.
|
| [32] |
张淼淼, 秦浩, 王烨, 张峰. 汾河中上游湿地植被β多样性. 生态学报, 2016, 36(11): 3292-3299. |
| [33] |
Saito V S, Perkins D M, Kratina P. A metabolic perspective of stochastic community assembly. Trends in Ecology & Evolution, 2021, 36(4): 280-283. |
| [34] |
Gibson N, Prober S, Meissner R, van Leeuwen S. Implications of high species turnover on the south-western Australian sandplains. PLoS One, 2017, 12(2): e0172977. |
| [35] |
何池全, 赵魁义, 余国营. 湿地克隆植物的繁殖对策与生态适应性. 生态学杂志, 1999, 18(6): 38-46. |
| [36] |
Flinn K M, Gouhier T C, Lechowicz M J, Waterway M J. The role of dispersal in shaping plant community composition of wetlands within an old-growth forest. Journal of Ecology, 2010, 98(6): 1292-1299. |
| [37] |
Su H J, Chen J, Wu Y, Chen J F, Guo X C, Yan Z B, Tian D, Fang J Y, Xie P. Morphological traits of submerged macrophytes reveal specific positive feedbacks to water clarity in freshwater ecosystems. The Science of the Total Environment, 2019, 684: 578-586. |
| [38] |
楼倩. 鄱阳湖湿地植被群落β-多样性格局[D]. 南昌: 南昌大学, 2016.
|
| [39] |
Silvertown J. Plant coexistence and the niche. Trends in Ecology & Evolution, 2004, 19(11): 605-611. |
| [40] |
Santamaría L. Why are most aquatic plants widely distributed? Dispersal, clonal growth and small-scale heterogeneity in a stressful environment. Acta Oecologica, 2002, 23(3): 137-154. |