 2023, Vol. 43
2023, Vol. 43文章信息
- 李斌, 郑宇辰, 徐丹丹, 陶敏, 李红
- LI Bin, ZHENG Yuchen, XU Dandan, TAO Min, LI Hong
- 长江上游弥陀漫滩水体鱼类食物网动态的季节性变化
- Seasonal variation in the consumption of food by fish in the Mituo floodplain waters of the upper Yangtze River and implications for food web dynamics
- 生态学报. 2023, 43(4): 1664-1675
- Acta Ecologica Sinica. 2023, 43(4): 1664-1675
- http://dx.doi.org/10.5846/stxb202203240734
-
文章历史
- 收稿日期: 2022-03-24





2. 农业农村部长江下游渔业资源环境科学观测实验站, 中国水产科学研究院淡水渔业研究中心, 无锡 214081;
3. 长江上游鱼类资源保护与利用四川省重点实验室, 内江 641100
2. Scientific Observing and Experimental Station of Fishery Resources and Environment in the Lower Reaches of the Changjiang River, Ministry of Agriculture and Rural Affairs, Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi 214081, China;
3. Key Laboratory of Sichuan Province for Fishes Conservation and Utilization in the Upper Reaches of the Yangtze River, Neijiang 641100, China
长江上游是我国淡水鱼类生物多样性最为丰富的地区之一, 分布着286种鱼类, 其中珍稀、特有鱼类127种, 是我国最重要的鱼类种质资源库[1]。河流漫滩生态系统具有高的生产力和微生境异质性, 对维持区域鱼类生物多样性具有重要作用[2]。水位的季节性(洪枯)变化被认为是维持漫滩水体鱼类物种组成和分布的主要驱动力[3—4]。河流泛洪期漫滩与河道干流连通呈一体, 外源性营养物质和生物的输入拓展了漫滩水体消费者食源的多样性和营养生态位宽度, 改变了消费者与资源的耦合模式[5]。自然河流动力学特征能较好地解释泛洪漫滩水体生态系统功能和完整性的变化, 河流流量的改变与漫滩水体的连通性密切相关, 对关键生态过程产生级联效应[6]。漫滩作为河流生态系统的重要组成部分, 为鱼类提供了多样化的栖息生境, 鱼类在长期进化过程中也形成了与水位变化相适应的生物节律[7—8]。了解季节性变化对河流漫滩水体鱼类食物网结构和营养生态位的影响规律是管理和制定区域生物多样性保护措施的重要前提。然而针对河流漫滩水体鱼类营养与水位变化关联的研究仍十分匮乏且主要集中在长江中下游区域[9—10], 上游河段很少被关注。
弥陀河段位于长江上游的泸州市弥陀镇, 该河段独特的河床地质结构(边滩和辫状河道堆积)在河道右岸形成了漫滩水体生态系统[11](以下简称为弥陀漫滩水体)。该区域丰水期与长江干流完全连通(上、下端均连通), 枯水期与干流断开呈半封闭或封闭的静水状态, 生境异质性高, 适合鱼类的生存繁衍。开展弥陀漫滩水体水生食物网结构季节性动态变化的特征研究对于保护长江上游生物多样性具有重要的指导意义, 而目前尚无该水域鱼类营养结构特征等相关报道。近年来, 稳定碳、氮同位素(δ13C、δ15N)技术作为生态学领域研究的一种重要手段, 已被广泛用于探究不同水域食物网结构特征[12—13]。该方法主要基于δ13C、δ15N在食物链传递过程中的分馏规律(即消费者δ13C、δ15N值与其食物相比富集度分别约为0—1‰和3.4‰)来确定基础碳源对食物网的贡献水平[12]及消费者营养级位置[14—15]。本研究以长江上游弥陀漫滩水体为对象, 应用δ13C、δ15N技术和Bayesian混合模型, 并结合理化指标探究该区域内鱼类营养生态位、食物网结构、营养级长度及基础碳源对食物网贡献的季节性变化规律, 以期为长江上游漫滩水体生态系统鱼类生物多样性保护提供理论基础。
1 材料与方法 1.1 研究区域概况弥陀漫滩水域位于长江上游泸州河段弥陀镇。弥陀漫滩水体长约3.5km, 其主要地貌特征为巨砾、沙洲和沼泽, 该区域属亚热带湿润气候, 雨量充足, 年均降雨量达1200 mm主要集中在6—7月份, 常年平均气温18.2℃;在研究区域的右侧种植有大面积季节性蔬菜, 沿岸优势陆生和水生植物主要是狗尾草Setaria faberii、狗牙根Cynodon dactylon、芒类植物Miscanthus sp.和水蓼Polygonum hydropiper。通常, 丰水期(夏季5—8月)该水域与长江干流连通呈一整体(上、下两端均连通), 枯水期间(秋季9—11月)与干流断开形成一个相对封闭的静水体。依据研究水域生境特征, 设置了3个采样点(图 1), 分别于丰水期(2019年8月, 连通状态, 水域面积约3.15km2, 最大水深15.5m)和枯水期(2019年11月, 完全断开后约2个月, 水域面积约0.5 km2, 最大水深约6.0m, 河面宽度100—180m)对该区域进行了样品采集工作。

|
| 图 1 样点示意图 Fig. 1 Distribution of sampling sites |
使用5L辛德勒采水器采集表层水样, 密封冷藏运回实验室。水体透明度(SD)用塞氏盘现场度量。使用便携式水质分析仪器(YSI-550A等)原位pH和溶解氧(DO)。其余水化学指标, 包括总氮(TN)、总磷(TP)以及化学需氧量(CODMn)的测定参照国家环境保护总局[16]标准执行。叶绿素a(Chla)测定参照金相灿等[17]和章宗涉等[18]方法, 计算公式为: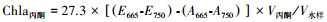
本研究中稳定同位素样品采集参考徐军[19]和曾庆飞等[20]方法, 具体如下:基础碳源采集:徒手采集弥陀漫滩水体周边和干流上游沿岸优势水生、陆生植物, 主要包括水蓼、狗尾草、狗牙根和芒科植物等。颗粒有机物采集(POM):分别在设置的三个样点采集POM, 取表层水, 先经浮游动物网(112μm)过滤, 将滤液再用玻璃纤维滤膜(GF/C Whatman)过滤, 最后把含有颗粒有机物样品的滤膜用锡箔纸包裹, 存放于封口袋中。固着藻类采集:用毛刷从沿岸的石头上刮取, 挑出易区分的杂质, 过滤到滤膜上(GF/C whatman), 处理方法同POM。底栖动物样品采集:利用手抄网(100目)采集水生昆虫(优势类群为扁蜉类);徒手采集背角无齿蚌Anodonta woodiana和铜锈环棱螺Bellamya aeruginosa等软体动物, 分别放置于充分曝气的蒸馏水中过夜, 使其肠含物排空, 然后去除外壳获取软组织。鱼类样品采集:鱼类资源调查参考《河流水生生物调查指南》[21]进行, 在丰水期和枯水期聘请同一位渔民, 使用相同捕捞工具(刺网:规格, 25m×12m、网目>2.0cm和地笼网:规格, 0.5m×0.5m、网目>2.0cm)连续捕捞作业3天, 在鲜活状态下测量鱼类基本形态学参数并取鱼类背侧肌肉存放(样品重量能满足同位素测试分析即可), 见表 1。
| 物种 Species |
缩写 Code |
丰水期Wet season | 枯水期Dry season | |||||
| 体长/cm Body length |
体重/g Body weight |
n | 体长/cm Body length |
体重/g Body weight |
n | |||
| 草鱼Ctenopharyngodon idellus | Ctid | 35.93±3.41 | 903.28±355.1 | 3 | 29.98±4.22 | 391.35±147.6 | 4 | |
| 鲫Carassius auratus | Caau | 12.50±4.09 | 34.80±23.91 | 3 | 14.25±1.15 | 66.99±16.86 | 4 | |
| 岩原鲤Procypris rabaudi | Prra | 11.50 | 30.91 | 1 | ||||
| 鲤Cyprinus (Cyprinus) carpio | Cyca | 16 | 89.45 | 1 | ||||
| 黑尾餐 Hemniculter tchangi | Hetc | 15.80±3.99 | 34.00±15.61 | 4 | 10.80±0.6 | 9.55±1.98 | 3 | |
| 高体近红鲌Ancherythroculter kurematsui | Anku | 14.30 | 33.01 | 1 | ||||
| 翘嘴鲌Erythroculter ilishaeformis | Erha | 21.83±8.95 | 135.96±86.79 | 3 | 18.80 ±2.12 | 45.28±15.03 | 2 | |
| 蒙古鲌Culter mongolicus | Cumo | 30.00 | 382.49 | 1 | ||||
| 四川华鳊Sinibrama taeniatus | Sita | 9.00 | 13.10 | 1 | ||||
| 似鳊Pseudobrama simoni | Pssi | 12.86±0.21 | 35.60±4.01 | 3 | ||||
| 黄尾鲴Xenocypris davidi | Xeda | 18.20 | 76.60 | 1 | ||||
| 圆吻鲴Distoechodon tumirostris | Ditu | 18.3 | 74.41 | 1 | ||||
花 |
Hemi | 9.90±0.10 | 13.00±0.00 | 3 | 10.25±0.95 | 12.47±1.29 | 3 | |
| 蛇鮈Saurogobio dabryi | Sada | 12.23±0.80 | 18.85±4.61 | 3 | 13.20±1.48 | 17.78±6.37 | 4 | |
| 吻鮈Rhinogobio typus | Rhty | 23.5±4.85 | 206.74±25.08 | 2 | ||||
| 长鳍吻鮈 Rhinogobio ventralis | Rhve | 21.45±0.64 | 112.13±0.18 | 2 | ||||
| 银鮈Squalidus argentatus | Sqar | 9.06±0.41 | 11.03±1.51 | 3 | ||||
| 中华鳑鲏Rhodeus sinensis | Rhsi | 6.70±0.44 | 5.55±1.11 | 3 | ||||
| 南方鲇Silurus meridionalis | Sime | 10.60 | 10.93 | 1 | ||||
| 鲇Pelteobagrus vachelle | Sias | 20.5 | 51.32 | 1 | ||||
| 光泽黄颡 Pelteobagrus nitilus | Peni | 11.50 | 17.12 | 1 | 11.20±0.46 | 11.90±1.35 | 3 | |
| 瓦氏黄颡鱼Pelteobagrus vachelle | Peva | 14.68±5.29 | 44.77±45.91 | 4 | 11.33±1.81 | 15.28±6.32 | 4 | |
| 黄颡鱼Pelteobagrus fulvidraco | Pefu | 10.50±0.47 | 18.16±3.29 | 4 | ||||
| 切尾拟鲿Pseudobagrus truncatus | Pstr | 14.00 | 24.02 | 1 | ||||
| 大鳍鱯Hemibagrus macropterus | Hema | 11.35±0.92 | 13.33±3.56 | 2 | ||||
| 长吻鮠Leiocassis longirostris | Lelo | 15.40±1.19 | 36.38±0.18 | 2 | ||||
| 大眼鳜Siniperca kneri | Sikn | 8.60±0.28 | 10.65±2.80 | 2 | ||||
| 子陵栉鰕虎鱼Rhinogobius giurinus | Rhgi | 7.03±0.15 | 8.22±1.82 | 3 | ||||
上述所有样品均密封冷藏运回实验室进行预处理。用于碳稳定性同位素分析的固着藻类滤膜先用1 mol/L的盐酸处理, 再用蒸馏水冲洗干净。最后将所有样品在60℃烘干至恒重, 用研钵研磨成均一粉末, 放入干燥器中保存待测。
1.4 稳定性同位素测试实验样品的稳定C、N同位素比值在北京林业科学院同位素实验室用稳定同位素质谱仪测定, 该仪器由元素分析仪(菲尼根Flash EA112 HT)与同位素比率质谱仪(菲尼根DELTA V Advantage)相连而成。
稳定C、N同位素的自然丰度表示为:

|
式中:X为13C或15N, R为13C/12C或15N/14N。δ13C值是相对于国际PDB标准, δ15N值是相对于空气氮气的丰度。分析精度δ13C < 0.10‰, δ15N < 0.20‰。
1.5 统计与分析营养级水平(TP)计算公式[22]:
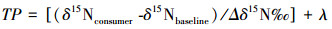
|
式中, Δδ15N=3.4‰;当baseline初级消费者时, λ=2。
Bayesian混合模型分析[23]:参考McCutchan等[24]和Mao等[25]方法, 应用R软件分析弥陀漫滩水体基础碳源对消费者的贡献比例。根据基础碳源来源并结合其δ13C、δ15N值重新进行了分类。主要包括以下五类:(1)POM、(2)固着藻类、(3)C3植物:芒科类、(4)C4植物:狗尾草和狗牙根、(5)水生维管束植物:水蓼。枯水期间未能采集到C3植物、C4植物和水生维管束植物, 但已有研究表明季节变化对陆生植物和水生维管束植物δ13C、δ15N值影响不大[26], 因此, 枯水期碳源贡献分析引用了丰水期的部分数据。
鱼类群落营养结构[27—28]:氮值范围NR(δ15N rang)代表了食物网的垂直结构, 是对营养长度的衡量;碳值范围CR(δ13C rang)代表了食物网底部的生态位多样化;平均离心距离(CD)是每个样本到双坐标图中心点的平均欧氏距离, 反映了营养多样性的平均程度;平均最邻近距离(MNND)是与每个样本的近邻样本的欧氏距离的平均值, 提供了关于物种总体密度信息;最邻近距离标准差(SDNND)衡量物种分布的均匀性;总面积(TA)表示消费者生态位空间面积。基于Bayesian方法计算的标准椭圆面积(SEAC)是消费者在食物网中的同位素生态位面积[28], 比Layman等[29]指标中TA更准确, 且对样品数量的变化不敏感。本研究分别计算了丰水期和枯水期弥陀漫滩水体鱼类群落营养结构的相关参数, 并比较了消费者(底栖动物和鱼类)在不同季节同位素生态位空间面积的大小。
本研究中鱼类群落营养结构参数的计算均在R(4.1.2)软件(SIBER程序包、SIAR程序包)中完成;基本数据的统计分析在STATISTICA.7.0和SPSS 22.0软件中完成。
2 结果与分析 2.1 理化参数表 2显示了弥陀漫滩水体在丰水期和枯水期主要理化参数。丰水期研究区域(水文连通, 水位上涨)除了透明度(SD)和叶绿素a(chla)两项指标低与枯水期(水文断开, 水位降低)外, 其余指标均高于枯水期, 其中溶解氧(DO)和总氮(TN)指标差异较为明显。
| 理化指标Physical and chemical indicators | 枯水期Dry season | 丰水期Wet season |
| 溶解氧Dissolved oxygen/(mg/L) | 7.85±0.71 | 9.58±0.27 |
| pH | 7.65±0.24 | 7.91±0.32 |
| 透明度Secchi depth/ cm | 120.00±35.6 | 15.80±3.61 |
| 总磷Total phosphorus/(mg/L) | 0.62±0.31 | 0.77±0.21 |
| 总氮Total nitrogen/(mg/L) | 0.78±0.21 | 1.12±0.43 |
| 化学需氧量Chemical oxygen demand/(mg/L) | 1.08±0.32 | 1.42±0.27 |
| 叶绿素a Chlorophyll a /(μg/L) | 45.85±6.55 | 12.02±2.92 |
调查期间共收集基础碳源样品26个, 分为内源性碳源和外源性碳源两大类。外源性碳源包括C4植物(狗尾草和狗牙根)和C3植物(芒类植物);内源性碳源由水中颗粒有机物(POM主要成分为浮游藻类)、固着藻类和水生维管束植物(水蓼)组成。就δ13C值而言(图 2和图 3):丰水期(完全连通)基础碳源的δ13C平均值-23.02‰, 变幅为-31.01‰—-11.2‰;枯水期(与干流断开)基础碳源(POM和固着藻类)δ13C平均值-21.93‰, 变幅为-26.31‰—-15.36‰;丰水期POM和固着藻类的δ13C组成均显著低于枯水期间的POM和固着藻类(P < 0.05)。在δ15N组成方面, 丰水期基础碳源的平均δ15N值为2.58‰, 范围为-0.51‰—6.84‰;枯水期POM和固着藻类平均δ15N值为7.22‰, 范围为4.89‰—8.81‰。POM和固着藻类的δ15N值与δ13C值均呈现出丰水期显著低于枯水期的趋势(P < 0.05)。

|
| 图 2 丰水期不同生物类群δ13C-δ15N双坐标图 Fig. 2 Biplot of δ13C and δ15N of sampled basal resources of sampled basal resources, Macrobenthoss and fish collected in the wet season 水生维管束植物(Macrophyte)、颗粒有机物(Particulate organic matter, POM)和固着藻类(Periphyton);扁蜉Heptageniidae (Heni)、背角无齿蚌Anodonta woodiana (Anwo)、铜锈环棱螺Bellamya aeruginosa (Beae)、秀丽白虾Leander modestus (Lemo)和日本沼虾Macrobrachium mipponensis (Mami) |

|
| 图 3 枯水期不同生物类群δ13C-δ15N双坐标图 Fig. 3 Biplot of δ13C and δ15N of sampled basal resources of sampled basal resources, Macrobenthoss and fish collected in the dry season |
消费者样品共计117个, 其中底栖动物样品29个, 鱼类样品88个。丰水期, 底栖动物δ13C平均值为-22.59‰, 范围为-25.63‰—-17.55‰, 其δ15N平均值为10.36‰, 范围为8.69‰—11.58‰(图 2);枯水期底栖动物δ13C、δ15N平均值分别为-25.33‰和9.77‰, 范围分别为-31.65‰—-21.36‰(δ13C)和7.84‰—11.63‰(δ15N)(图 3)。总体而言, 丰水期底栖动物δ13C值显著高于枯水期(P < 0.05), δ15N值略高于枯水期(P=0.23)。
丰水期共采集到鱼类23种, 其δ13C平均值为-22.32‰, 范围为-29.85‰—-18.13‰;其δ15N平均值为10.45‰, 范围为5.61‰—14.36‰。枯水期采集到13种鱼类, 其δ13C、δ15N平均值分别为-21.99‰、10.03‰, 变化范围分别为-24.01‰—-18.52‰、5.72‰—13.39‰。总的来说丰水期鱼类δ13C(P=0.44)、δ15N(P=0.26)值组成与枯水期相比差异均不显著。
2.3 营养级长度依据Post[22]的建议, 本研究选择两个季节均采集到的底栖动物蜉蝣目扁蜉Heptageniidae作为基线生物计算了其它消费者的营养级水平。丰水期食物网营养级长度为3.6个营养级, 多数鱼类的营养级位于第二至第三级之间(杂食性鱼类居多);其中, 以岩原鲤Procypris rabaudi 营养级水平为最高, 位于食物网顶端(图 2), 而草鱼Ctenopharyngodon idellus(1.31)的营养级水平最低, 位于食物网的底层。枯水期调查区域食物网的长度不足3级(2.69), 多数鱼类级处于第二营养级水平, 以翘嘴鲌Erythroculter ilishaeformis营养级水平为最高(2.7), 位于食物网顶端, 草鱼最低位于食物网底层(图 3)。
2.4 Bayesian混合模型Bayesian混合模型分析结果显示:在枯水期底栖动物的基础碳源主要来自内源性营养物质固着藻类和外源性营养物质C3植物(图 4), 固着藻类平均贡献率为35.33%(Mode=36.66%, 置信区间(Confidence interval, CI)= 7%—62%), C3植物平均贡献率为30.71%(Mode=29.62%, CI=5%—53%);鱼类基础碳源主要来自内源性营养物质POM和固着藻类(图 5), 其中POM的平均贡献率为40.70%(Mode=43.45%, CI=18%—43%), 固着藻类的平均贡献率为30.24%(Mode=30.46%, CI =1%—56%)。

|
| 图 4 枯水期不同基础碳源对大型底栖动物的贡献比例 Fig. 4 Contribution ratio of different basal carbon sources to macrobenthos in the dry season |

|
| 图 5 枯水期不同基础碳源对鱼类的贡献比例 Fig. 5 Contribution ratio of different basal carbon sources to fish in the dry season |
丰水期, 底栖动物的基础碳源主要由外源性C4植物和C3植物供给(图 6), 其中C3植物的平均贡献率为29.99%(Mode=28.53%, CI=3%—55%), C4植物的平均贡献率为29.62%(Mode=31.00%, CI=17%—42%);鱼类的主要营养来源和底栖动一致都是以外源性营养物质C3植物和C4植物为主(图 7), 其中C3植物的平均贡献率为57.38%(Mode=58.64%, CI=48%—64%), C4植物的平均贡献率为35.55%(Mode=35.39%, CI=31%—39%)。

|
| 图 6 洪水期不同基础碳源对大型底栖动物的贡献比例 Fig. 6 Contribution ratio of different basal carbon sources to macrobenthos in the wet season |

|
| 图 7 洪水期不同基础碳源对鱼类的贡献比例 Fig. 7 Contribution ratio of different basal carbon sources to fish in the wet season |
应用SIBER分析了丰水期与枯水期弥陀漫滩水体底栖动物和鱼类同位素生态位空间面积和群落结构指标(表 3)。枯水期, 底栖动物的SEAB, SEAC和Layman指标(NR、CR、CD、TA)均高于丰水期, 表明枯时期间底栖动物拥有更大的生态位空间(图 8), 但丰水期间MNND和SDNND指标略小于枯水期。鱼类丰水期SEAB, SEAC和Layman指标(NR、CR、MNND、SDNND、CD、TA)均高于枯水期间, 说明丰水期间鱼类占据了更大的生态位空间(图 9)。
| NR | CR | CD | MNND | SDNND | TA | SEAB | SEAC | n | |
| 底栖动物Macrobenthos | |||||||||
| 洪水期Wet season | 2.89 | 8.08 | 2.24 | 0.99 | 0.73 | 9.03 | 6.07 | 6.93 | 10 |
| 枯水期Dry season | 3.79 | 10.76 | 3.54 | 0.76 | 0.51 | 19.14 | 8.29 | 8.77 | 19 |
| 鱼类Fish | |||||||||
| 洪水期Wet season | 9.75 | 11.72 | 2.41 | 0.59 | 0.72 | 60.67 | 12.28 | 12.53 | 51 |
| 枯水期Dry season | 7.67 | 8.92 | 1.98 | 0.57 | 0.58 | 38.19 | 8.31 | 8.55 | 37 |
| NR:氮值范围δ15N rang; CR:碳值范围δ13C rang; CD:平均离心距离Mean distance to centroid; MNND:平均最邻近距离Mean nearest neighbor distance; SDNND:最邻近距离标准差Standard deviation of nearest neighbor distance; TA:总面积Total area; SEAB:基于贝叶斯椭圆面积Bayesian elliptic area;SEAc:标准椭圆面积Standard elliptic area | |||||||||

|
| 图 8 研究区域不同季节底栖动物碳和氮同位素空间面积 Fig. 8 Carbon and nitrogen isotopic spaces occupied of macrobenthos communities in different seasons in the study area |

|
| 图 9 研究区域不同季节鱼类碳和氮同位素空间面积 Fig. 9 Carbon and nitrogen isotopic spaces occupied of fish communities in different seasons in the study area |
河流碳源依据其来源途径可分为内源性和外源性两种类型[30], 其中内源性碳源是指河流中藻类与高等水生植物等营养物质, 外源性碳源是指来源于河流以外的营养物质包括植物根、茎、叶和果实等[31—32]。
本研究结果显示丰水期研究区域鱼类食物网能量主要依赖于外源性营养物质, 枯水期则是以内源性营养物质为主要贡献者。不同水文连通情况下, 水体的理化参数存在差异(表 1), 相关参数的变化对整个生态系统营养动力学特征产生影响[33]。研究发现丰水期研究区域鱼类同位素生态位空间面积、鱼类群落营养指标和鱼类的营养多样性等参数明显增加, 可能与该区域和长江干流完全连通后(大量外源性营养物质的输入), 鱼类利用了不同来源的碳源或多样化栖息生境有关[29, 34—35]。在丰水期, 陆生C3植物和C4植物对鱼类食物网能量的贡献比例较高(分别为57.38%和35.55%), 促进了鱼类群落结构的营养多样化;河流脉冲作用改变了研究区域水体环境(例如增加水体浊度、降低透明度、改变营养物的量等), 相对较低的Chla含量反映了藻类作为潜在碳源(内源性)的重要性降低, 而丰富的外源性营养物质的输入为消费者提供了更多的选择空间[36]。例如肖协文[37]在饶河流域发现, 随着流域面积的增加, 下游工、农业区带入了大量的有机物质进入饶河食物网内, 导致外源性营养物质在河流食物网能量中的贡献比例明显增大。同时, 研究发现水文的连通为消费者提供了重要的能量路径, 能够提升该区域物种多样性和系统整体生产力水平;消费者对外源性营养物质利用的大幅增加突显了水文连通对支撑该区鱼类生物多样性的重要性[38—39]。
与丰水期相比, 枯水期间研究区域鱼类生态位空间面积和鱼类群落指标均有所下降。相对较低的鱼类群落营养多样性(CR)(表 3)暗示了鱼类可利用食物资源的有限性或栖息生境的简单化[29, 35, 40]。Bayesian混合模结果显示了POM和固着藻类是鱼类食物网能量的主要贡献者(分别为40.70%和30.24%)。枯水期(调查期间弥陀漫滩水体与长江干流断开时间超过两个月), 该区域与干流河道的连接完全断开, 水体透明度显著增加, 此时相对较高的Chla含量表明藻类生物量较大为消费者提供了潜在内源性营养物质[41], 这一结论与Xu等[10]在长江中下游漫滩水域中发现的枯水期消费者能量主要依赖于内源性营养物质的研究结果是一致的。对于该区域而言, 季节性变化导致的水文连通性改变影响了消费者与食物资源之间的耦合模式, 增加了弥陀漫滩水体生态系统在枯水期间的脆弱性[42]。
3.2 季节脉冲对鱼类生态位的影响已有研究表明, 泛洪漫滩水体鱼类营养多样性、生态位空间大小和资源的可利用性受特定区域环境变化的影响, 也与该区域季节性脉冲有关[43]。
丰水期河流水体/陆地群落交错区的扩张, 是决定河流食物网的重要因素, 河流两岸的“脉冲”被认为是控制泛洪漫滩水体生产力和动植物种群的主要因素[38]。在研究区域, 水文连通时段主要集中出现在6—8月, 此时土壤水分饱和, 地表径流和河流流量可能达到峰值, 水域面积(生态系统)也达到了最大延伸。在河流脉冲的作用下, 干流上游和周边的潜在碳源(包括外源性营养)、鱼类等其他水生生物的输入增加了基础碳源的多样性(CR)和栖息生境的多样化, 进而支撑了该区域鱼类物种的多样性(23种);研究区域鱼类群落同位素生态位空间(TA、SEAB和SEAC)面积的增大和鱼类食物网生态位冗余现象的出现提升了丰水期间生态系统结构的稳定性。已有研究显示[31, 44], 河流脉冲作用能够促进漫滩水体微生物循环过程, 激活水体中的异养活性(反硝化作用), 增加潜在食源的δ15N值, 这可能是导致丰水期鱼类群落NR值升高的重要因素。枯水期, 研究区域与干流断开切断了干流与该区域营养物质横向交换的途径, 水量补充主要依赖于主河道渗漏和周边集水区域径流;研究区域外来营养物质输入的减少和水位的回落, 导致种间竞争作用增强及对空间资源利用率下降, 进而引发了鱼类物种多样性的降低(13种)和鱼类群落生态位空间面积的减小[45, 5]。
3.3 食物网营养级长度营养级指示了物质与能量从食物网底端的初级生产者到顶端捕食者的流动过程, 能够表征食物网结构的线状特征[46]。影响食物网营养级长度的因素很多, 包括群落结构、资源可获得性和物种相互作用关系等因素[47]。丰水期研究区域水域面积(生态系统)达到最大, 外源性营养物质输入和栖息生境的多样化驱动了食物网营养多样性与空间变异过程[40, 48], 延伸了食物网营养级长度[49], 而在枯水期水域面积、营养物质组成和生境类型等方面与上述情况正好相反。例如刘军等[50]在对长江上游主要河流鱼类多样性与流域特征关系研究中发现, 物种的绝对数量与流域面积和流量呈显著正相关;Vander Zanden和Fetzer[51]在全球范围内收集并评估了水生态系统食物网, 发现生态系统大小与湖泊营养级长度之间存在相关关系。因此, 生态系统大小理论解释了该区域不同季节鱼类食物网营养多样性和食物链长度的变化过程。
与自然来源相比, 人类产生的氮富含15N, 例如城市废水、粪肥或农业肥料的δ15N值高达15‰[52], 这种富集的δ15N信号可以通过食物链传递进入食物网[12]。本次调查并未发现明显的δ15N值富集信号, 可能与近年来长江上游水域生态环境持续改善有关。为了更加准确的了解长江上游水生生态环境保护措施的有效性, 后续仍需继续开展相关监测研究工作。
4 结论(1) 本研究中丰水期鱼类食物网能量主要依赖于外源性营养物质, 包括C4植物和C3植物;枯水期则以POM、固着藻类和水生维管束植物等内源性营养物质为主要贡献者。
(2) 水文连通性的季节性变化改变了消费者与食物资源之间的耦合模式。研究区域丰水期水文的连通为消费者提供了重要的能量路径, 提升物种多样性和系统整体生产力水平;水体的断开则降低了生态系统在枯水期间的稳定性。
(3) 丰水期基础碳源的增加和栖息生境的多样化拓宽了鱼类群落生态位空间, 鱼类食物网生态位冗余的出现提升了生态系统结构的稳定性;枯水期外来营养物质输入的减少和水位的回落则导致了鱼类物种多样性下降和鱼类生态位空间面积的减小。
(4) 丰水期研究水域鱼类食物网营养多样性和食物链长度均大于枯水期, 由两个季节水域面积(生态系统大小)差异所决定, 这一结果支持了生态系统大小理论。
综述所述, 保证漫滩水域季节性的连通性和减少农业径流污染对维持该区域生物多样性及生态系统稳定性方面具有重要的意义。
| [1] |
杨志, 唐会元, 龚云, 董纯, 陈小娟, 万成炎, 常剑波. 正常运行条件下三峡库区干流长江上游特有鱼类时空分布特征研究. 三峡生态环境监测, 2017, 2(1): 1-10. DOI:10.19478/j.cnki.2096-2347.2017.01.01 |
| [2] |
Amoros C, Bornette G. Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshwater Biology, 2002, 47(4): 761-776. DOI:10.1046/j.1365-2427.2002.00905.x |
| [3] |
Junk W J. Flood pulsing and the linkages between terrestrial, aquatic, and wetland systems. SIL Proceedings, 1922-2010, 2005, 29(1): 11-38. DOI:10.1080/03680770.2005.11901972 |
| [4] |
Rood S B, Samuelson G M, Braatne J H, Gourley C H, Hughes F M R, Mahoney J M. Managing river flows to restore floodplain forests. Frontiers in Ecology and the Environment, 2005, 3(4): 193-201. DOI:10.1890/1540-9295(2005)003[0193:MRFTRF]2.0.CO;2 |
| [5] |
Winemiller K O, Montaña C G, Roelke D L, James B, Cotner J B, Montoya J V, Sanchez L, Castillo M M, Layman C A. Pulsing hydrology determines top-down control of basal resources in a tropical river-floodplain ecosystem. Ecological Monographs, 2014, 84(4): 621-635. DOI:10.1890/13-1822.1 |
| [6] |
Thoms M C. Floodplain-river ecosystems: lateral connections and the implications of human interference. Geomorphology, 2003, 56(3/4): 335-349. |
| [7] |
Welcomme R L. Fisheries Ecology of Floodplain Rivers. New York: Longman, 1979.
|
| [8] |
Winemiller K O. Development of dermal lip protuberances for aquatic surface respiration in South American characid fishes. Copeia, 1989(2): 382-390. |
| [9] |
王朝, 周立志, 戴秉国, 古辰, 蒋忠冠. 水位洪枯变化对菜子湖江湖过渡带鱼类物种和功能多样性的影响. 湖泊科学, 2019, 31(5): 1403-1414. |
| [10] |
Xu J, Wen Z R, Gong Z J, Zhang M, Xie P, Hansson L A. Seasonal trophic niche shift and cascading effect of a generalist predator fish. PLoS One, 2012, 7(12): e49691. DOI:10.1371/journal.pone.0049691 |
| [11] |
王洪铸, 刘学勤, 王海军. 长江河流-泛滥平原生态系统面临的威胁与整体保护对策. 水生生物学报, 2019, 43(S1): 157-182. DOI:10.7541/2019.178 |
| [12] |
Fry B. Food web structure on Georges Bank from stable C, N, and S isotopic compositions. Limnology & Oceanography, 1988, 33(5): 1182-1190. |
| [13] |
Mateo M A, Serrano O, Serrano L, Michener R H. Effects of sample preparation on stable isotope ratios of carbon and nitrogen in marine invertebrates: implications for food web studies using stable isotopes. Oecologia, 2008, 157(1): 105-115. DOI:10.1007/s00442-008-1052-8 |
| [14] |
Minagawa M, Wada E. Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochimica et Cosmochimica Acta, 1984, 48(5): 1135-1140. DOI:10.1016/0016-7037(84)90204-7 |
| [15] |
Hill J M, Jones R W, Hill M P, Weyl O L F. Comparisons of isotopic niche widths of some invasive and indigenous fauna in a South African river. Freshwater Biology, 2015, 60(5): 893-902. DOI:10.1111/fwb.12542 |
| [16] |
国家环境保护总局, 水和废水监测分析方法编委会. 水和废水监测分析方法 (第四版). 北京: 中国环境科学出版社, 2002: 88-284.
|
| [17] |
金相灿, 屠清瑛. 湖泊富营养化调查规范 (第二版). 北京: 中国环境科学出版社, 1990: 268-270.
|
| [18] |
章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社, 1991: 1-414.
|
| [19] |
徐军. 应用碳、氮稳定性同位素探讨淡水湖泊的食物网结构和营养级关系[D]. 武汉: 中国科学院研究生院(水生生物研究所), 2005.
|
| [20] |
曾庆飞, 孔繁翔, 张恩楼, 钱善勤. 稳定同位素技术应用于水域食物网的方法学研究进展. 湖泊科学, 2008, 20(1): 13-20. DOI:10.3321/j.issn:1003-5427.2008.01.002 |
| [21] |
陈大庆. 河流水生生物调查指南. 北京: 科学出版社, 2014.
|
| [22] |
Post D M. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology, 2002, 83(3): 703-718. DOI:10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2 |
| [23] |
Parnell A C, Inger R, Bearhop S, Jackson A L. Source partitioning using stable isotopes: coping with too much variation. PLoS One, 2010, 5(3): e9672. DOI:10.1371/journal.pone.0009672 |
| [24] |
McCutchan Jr J H, Lewis Jr W M, Kendall C, McGrath C C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos, 2003, 102(2): 378-390. DOI:10.1034/j.1600-0706.2003.12098.x |
| [25] |
Mao Z G, Gu X H, Zeng Q F, Zhou L H, Sun M B. Food web structure of a shallow eutrophic lake (Lake Taihu, China) assessed by stable isotope analysis. Hydrobiologia, 2012, 683(1): 173-183. DOI:10.1007/s10750-011-0954-3 |
| [26] |
Currin C A, Newell S Y, Paerl H W. The role of standing dead Spartina alterniflora and benthic microalgae in salt marsh food webs: considerations based on multiple stable isotope analysis. Marine Ecology Progress Series, 1995, 121: 99-116. DOI:10.3354/meps121099 |
| [27] |
Layman C A, Quattrochi J P, Peyer C M, Allgeier J E. Niche width collapse in a resilient top predator following ecosystem fragmentation. Ecology Letters, 2007, 10(10): 937-944. DOI:10.1111/j.1461-0248.2007.01087.x |
| [28] |
Jackson A L, Inger R, Parnell A C, Bearhop S. Comparing isotopic niche widths among and within communities: SIBER-stable isotope Bayesian ellipses in R. Journal of Animal Ecology, 2011, 80(3): 595-602. DOI:10.1111/j.1365-2656.2011.01806.x |
| [29] |
Layman C A, Arrington D A, Montaña C G, Post D M. Can stable isotope ratios provide for community-wide measures of trophic structure?. Ecology, 2007, 88(1): 42-48. DOI:10.1890/0012-9658(2007)88[42:CSIRPF]2.0.CO;2 |
| [30] |
Kendall C, Silva S R, Kelly V J. Carbon and nitrogen isotopic compositions of particulate organic matter in four large river systems across the United States. Hydrological Processes, 2001, 15(7): 1301-1346. DOI:10.1002/hyp.216 |
| [31] |
Thorp J H, Delong M D. Dominance of autochthonous autotrophic carbon in food webs of heterotrophic rivers. Oikos, 2002, 96(3): 543-550. DOI:10.1034/j.1600-0706.2002.960315.x |
| [32] |
Carassou L, Whitfield A K, Bergamino L, Moyo S, Richoux N B. Trophic dynamics of the cape stumpnose (Rhabdosargus holubi, Sparidae) across three adjacent aquatic habitats. Estuaries and Coasts, 2016, 39(4): 1221-1233. DOI:10.1007/s12237-016-0075-3 |
| [33] |
Cazzanelli M, Soria-Barreto M, Castillo M M, Rodiles-Hernández R. Seasonal variations in food web dynamics of floodplain lakes with contrasting hydrological connectivity in the southern Gulf of Mexico. Hydrobiologia, 2021, 848(4): 773-797. DOI:10.1007/s10750-020-04468-8 |
| [34] |
Abrantes K G, Barnett A, Bouillon S. Stable isotope-based community metrics as a tool to identify patterns in food web structure in east African estuaries. Functional Ecology, 2014, 28(1): 270-282. DOI:10.1111/1365-2435.12155 |
| [35] |
Jackson M C, Donohue I, Jackson A L, Britton J R, Harper D M, Grey J. Population-level metrics of trophic structure based on stable isotopes and their application to invasion ecology. PLoS One, 2012, 7(2): e31757. DOI:10.1371/journal.pone.0031757 |
| [36] |
Abrantes K G, Barnett A, Marwick T R, Bouillon S. Importance of terrestrial subsidies for estuarine food webs in contrasting East African catchments. Ecosphere, 2013, 4(1): 1-33. |
| [37] |
肖协文. 基于稳定同位素技术的饶河食物网结构特征研究[D]. 北京: 中国科学院大学, 2014.
|
| [38] |
Junk W J, Bayley P B, Sparks R E. The flood pulse concept in river-floodplain systems. Canadian Journal of Fisheries and Aquatic Sciences, 1989, 106: 110-127. |
| [39] |
Pool T, Holtgrieve G, Elliott V, McCann K, McMeans B, Rooney N, Smits A, Phanara T, Cooperman M, Clark S, Phen C, Chhuoy S. Seasonal increases in fish trophic niche plasticity within a flood-pulse river ecosystem (Tonle Sap Lake, Cambodia). Ecosphere, 2017, 8(7): e01881. |
| [40] |
Kaymak N, Winemiller K O, Akin S, Altuner Z, Polat F, Dal T. Spatial and temporal variation in food web structure of an impounded river in Anatolia. Marine and Freshwater Research, 2018, 69(9): 1453-1471. DOI:10.1071/MF17270 |
| [41] |
Thomaz S M, Bini L M, Bozelli R L. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia, 2007, 579(1): 1-13. |
| [42] |
Jardine T D, Pettit N E, Warfe D M, Pusey B J, Ward D P, Douglas M M, Davies P M, Bunn S E. Consumer-resource coupling in wet-dry tropical rivers. Journal of Animal Ecology, 2012, 81(2): 310-322. DOI:10.1111/j.1365-2656.2011.01925.x |
| [43] |
Humphries P, Keckeis H, Finlayson B. The river wave concept: integrating river ecosystem models. BioScience, 2014, 64(10): 870-882. |
| [44] |
Cole J J, Carpenter S R, Kitchell J, Pace M L, Solomon C T, Weidel B. Strong evidence for terrestrial support of zooplankton in small lakes based on stable isotopes of carbon, nitrogen, and hydrogen. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(5): 1975-1980. |
| [45] |
Lowe-McConnell R H. Ecological Studies in Tropical Fish Communities. Cambridge: Cambridge University Press, 1987.
|
| [46] |
张欢, 何亮, 张培育, 沙永翠, 徐军. 食物链长度理论研究进展. 生态学报, 2013, 33(24): 7630-7643. |
| [47] |
Lawler S P, Morin P J. Food web architecture and population dynamics in laboratory microcosms of protists. The American Naturalist, 1993, 141(5): 675-686. |
| [48] |
Vander Zanden M J, Shuter B J, Lester N, Rasmussen J B. Patterns of food chain length in lakes: a stable isotope study. The American Naturalist, 1999, 154(4): 406-416. |
| [49] |
Post D M, Pace M L, Hairston Jr N G. Ecosystem size determines food-chain length in lakes. Nature, 2000, 405(6790): 1047-1049. |
| [50] |
刘军, 曹文宣, 常剑波. 长江上游主要河流鱼类多样性与流域特征关系. 吉首大学学报(自然科学版), 2004, 25(1): 42-47. |
| [51] |
Vander Zanden M J, Fetzer W W. Global patterns of aquatic food chain length. Oikos, 2007, 116(8): 1378-1388. |
| [52] |
Hershey A E, Northington R M, Finlay J C, Peterson B J. Stable isotopes in stream food webs//Lamberti G A, Hauer F R, eds. Methods in Stream Ecology. 3rd ed. San Diego: Academic Press, 2017: 3-20.
|