 2023, Vol. 43
2023, Vol. 43文章信息
- 杨永, 卫伟, 王琳, 刘泽漫
- YANG Yong, WEI Wei, WANG Lin, LIU Zeman
- 中国旱区样带尺度植物多样性和生产力与环境因子的关系
- Relationship between plant diversity, productivity and environmental factors at a transect scale in the dryland of China
- 生态学报. 2023, 43(4): 1563-1571
- Acta Ecologica Sinica. 2023, 43(4): 1563-1571
- http://dx.doi.org/10.5846/stxb202202110335
-
文章历史
- 收稿日期: 2022-02-11
- 网络出版日期: 2022-10-13

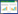




2. 中国科学院大学, 北京 100049;
3. 陕西黄土高原地球关键带国家野外科学观测研究站, 西安 710061
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. National Observation and Research Station of Earth Critical Zone on the Loess Plateau in Shaanxi, Xi'an 710061, China
生态系统植被特征与环境因子的关系研究一直是生态学研究的热点[1]。气温、光照、降水、土壤性质等环境因子的异质性对植物群落的物种组成、结构、多样性和生物量等具有重要影响[2—3]。普遍认为, 气候变化塑造大尺度上植被的分布。如Harrison等[4]认为气候变暖可能导致温带或凉爽地区的植物物种丰富度增加, 而干旱或热带地区的植物物种丰富度减少。Sun等[5]研究发现, 气候因素是驱动中国沙漠植物群落物种多样性定向损失的主要因素。同时, 植被的生长发育与其生境条件密切相关, 土壤理化性质的差异会影响群落结构、物种多样性和生产力水平[6], 如土壤水分[7]、土壤养分[8]和质地[9]等的变化会造成物种丰富度和群落生产力的改变。Yang等[10]研究发现, 中国荒漠草原地区植物多样性和生产力显著受土壤总氮、土壤总碳、土壤微生物生物量氮、土壤微生物生物量碳等因素的影响。从区域尺度而言, 土壤特征具有明显的时空异质性, 这对于植物多样性和生产力的空间分布格局有重要影响。因此, 定量解析环境因素对植物多样性和生产力的影响程度及解释作用, 对区域生态系统恢复和可持续发展具有重要指导意义。
生物多样性、生产力与环境因子间的相互作用十分复杂。研究认为, 植物多样性通过物种相互作用对生产力产生影响, 而物种相互作用的大小和方向在区域尺度上沿环境梯度变化[11]。因此, 环境因子和生物因子共同作用形成群落生产力[12]。Ali等[1]研究发现, 气候和土壤因素通过物种多样性和林分结构间接决定森林地上生物量。Li等[13]和Ma等[14]研究认为, 使用偏相关分析去除环境因素的影响后, 区域物种丰富度与生产力在区域尺度下的关系由显著正相关转为不相关。而Duffy等[15]对全球相关实验的荟萃分析结果表明, 植物多样性对生物量的驱动作用不因环境的影响而降低。因此, 环境因子和植物多样性对生产力的综合调控机制仍存在争议, 在我国旱区区域尺度上的相关研究结论尚未揭示。
我国旱区面积约660万km2, 承载着5.8亿人的生存发展[16], 而旱区生态系统极其脆弱且稳定性低, 是对气候变化和人类活动最敏感的地区之一[16—17]。现有研究多基于定点观测或遥感数据来分析生态系统对环境变化的响应, 而较大尺度的实地调查研究较为缺乏。基于此, 本研究以中国北方旱区广泛分布的典型灌草生态系统为研究对象, 沿水热梯度自东向西开展样带调查, 探究植物多样性和生产力间的关系, 揭示旱区植物多样性和生产力变化的影响因素及其解释作用。本研究有助于加深对旱区植物多样性、生产力和环境间关系的理解, 为高效利用和管理旱区生态系统提供理论指导, 为预测未来气候变化下旱区生态系统的响应提供依据。
1 材料与方法 1.1 研究区域概况本研究位于中国北方干旱半干旱生态系统, 涉及内蒙古自治区、陕西省、宁夏回族自治区、甘肃省、新疆维吾尔自治区5个省(自治区)。该区域主要属于温带大陆性气候, 年降雨量多在600 mm以下, 由东向西递减;年均温从-4℃到13℃, 区域内潜在蒸散发较高。内蒙古草原区主要土壤类型为钙积土, 黄土高原区主要土壤类型为黄绵土, 荒漠地区主要土壤类型为荒漠土和钙积土。该区域内植被盖度较低, 以耐旱抗风的灌草丛为主[18—19]。根据干旱指数划分研究区域的干旱程度(图 1), 干旱指数为年降水量与年潜在蒸发量的比值[20—21]。沿中国北方干旱梯度带, 选择12个市(盟)为调查点, 调查点的干旱程度由东向西逐渐加深, 干旱梯度明显。样带调查在2020年8月和2021年8月进行, 在每个市(盟)布设若干调查样地, 尽可能地包含该地区典型植物群落, 共计布设41块调查样地(图 1、表 1)。

|
| 图 1 采样点分布图 Fig. 1 Distribution map of sampling points |
| 行政区划 Administrative area |
布设样地数 Sampling numbers |
主要物种 Main species |
| 通辽市 | 1 | 黄花蒿Artemisia annua、小叶锦鸡儿Caragana microphylla、胡枝子Lespedeza bicolor |
| 锡林郭勒盟 | 4 | 羊草Leymus chinensis、针茅Stipa capillata、冰草Agropyron cristatum、碱韭Allium polyrhizum、委陵菜Potentilla chinensis |
| 乌兰察布市 | 2 | 针茅Stipa capillata、冷蒿Artemisia frigida、红砂Reaumuria songarica、盐爪爪Kalidium foliatum、碱韭Allium polyrhizum |
| 包头市 | 1 | 羊草Leymus chinensis、针茅Stipa capillata、糙隐子草Cleistogenes squarrosa |
| 阿拉善盟 | 4 | 红砂Reaumuria songarica、霸王Zygophyllum xanthoxylon、油蒿Artemisia ordosica、白刺Nitraria tangutorum、沙冬青Ammopiptanthus mongolicus、糙隐子草Cleistogenes squarrosa |
| 延安市 | 4 | 鼠李Rhamnus davurica、大油芒Spodiopogon sibiricus、甘菊Chrysanthemum lavandulifolium、铁杆蒿Artemisia gmelinii、猪毛蒿Artemisia scoparia、委陵菜Potentilla chinensis |
| 榆林市 | 2 | 沙蒿Artemisia desertorum、蒙古岩黄耆Hedysarum fruticosum、猪毛蒿Artemisia scoparia、狗娃花Heteropappus hispidus |
| 吴忠市 | 2 | 柠条锦鸡儿Caragana korshinskii、油蒿Artemisia ordosica、苦豆子Sophora alopecuroides |
| 武威市 | 6 | 沙拐枣Calligonum mongolicum、白刺Nitraria tangutorum、油蒿Artemisia ordosica、红砂Reaumuria songarica、梭梭Haloxylon ammodendron |
| 张掖市 | 6 | 红砂Reaumuria songarica、油蒿Artemisia ordosica、梭梭Haloxylon ammodendron、泡泡刺Nitraria sphaerocarpa |
| 酒泉市 | 5 | 红砂Reaumuria songarica、骆驼刺Alhagi sparsifolia、膜果麻黄Ephedra przewalskii、小果白刺Nitraria sibirica |
| 阜康市 | 4 | 梭梭Haloxylon ammodendron、白梭梭Haloxylon Persicum、红砂Reaumuria songonica、柽柳Tamarix chinensis |
调查样地选择尽可能排除人工干扰, 为多年围封地区或远离道路与居民的地区, 可以认为是无人工干预的自然生态系统。样地大小45 m×45 m, 调查方法按照BIODESERT国际规范化标准进行[22]。首先, 在每个样地布设4条间隔10 m的样线, 然后于每条样线随机设置5个1.5 m×1.5 m的调查样方, 调查并记录样方内出现的全部物种。
1.2 多样性指标计算本研究计算Shannon-Wiener(H)和群落性状加权值(CWM)两种指数, 分别作为物种多样性和功能多样性的表征。Shannon-Wiener指数基于物种的相对重要值计算。群落性状加权值基于6个叶片性状计算, 首先基于叶片性状的测量, 计算各性状的群落加权值;然后基于主成分分析, 提取其前两轴作为群落性状加权值的综合表征, 即多维群落性状加权值(CWM-leaf), 该指数包含了群落叶片性状的主要信息。叶片性状测量见相关文献[18]。
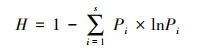
|
(1) |
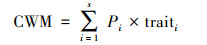
|
(2) |
式中, H代表样地内物种数;Pi代表样地内某物种的相对重要值, 相对重要值Pi = (相对多度+相对盖度+相对频度)/3;traiti为种i的性状特征值。
1.3 生产力和土壤养分数据采集与测定生产力使用地上生物量来表征。地上生物量是群落生产力的重要组成部分和直观表现, 是最为重要的生态系统功能之一。采用齐地面刈割法计算草本层的地上生物量, 选取3组1 m×1 m草本样方, 将草本植物地面以上部分剪下, 称其鲜重, 装入密封袋带回室内。采用标准株法计算灌木层的地上生物量, 由带队指导老师确定2株灌木标准株, 并估测样地的灌木植物多度, 剪下标准株灌木地面以上部分的四分之一, 称其鲜重, 装入密封袋带回室内。草本植物样品和灌木植物样品均在烘箱烘干(75℃, 48 h)至恒重后称其干重量(0.01 g)。计算3组草本植物样品生物量均值, 记为草本植物平均地上生物量(g/m2);由灌木植物标准株干重经换算得到灌木平均地上生物量(g/m2)。
在每个样地均匀设置3个土壤样品取样点, 采集0—20 cm的土壤并混合均匀, 分别采用重铬酸钾氧化法、氮分析仪测定法和碱熔消解-钼锑抗分光光度法测定土壤有机碳、土壤总氮、土壤总磷, 详细测定流程同样按照BIODESERT国际规范化方法进行[22]。测定后, 计算各样地的土壤养分指标的平均值。
1.4 气象数据来源气候因子选择年均气温和年均降水量, 数据来源于CHELSA生物气候年值数据集(Climatologies at high resolution for the earth′s land surface areas, http://chelsa-climate.org/), 该数据是基于1979—2013年全球气象数据的平均值进行插值得到, 分辨率为30弧秒。使用ArcGIS 10.2将气象数据按经纬度提取至各个样点。
1.5 数据处理与分析采用回归分析探究气候、土壤养分与植物多样性和生产力间关系以及多样性和生产力间关系。然后进一步使用层次分割的方法量化各环境因子对植物多样性和生产力的解释率, 该方法可有效避免变量间共线性的影响, 并通过置换检验的方法检验解释率是否显著[23]。该部分在R的“rdacca.hp”包中完成。
为探究环境因子和植物多样性对生产力的复杂作用机制, 本文使用了基于有向分离方法的验证性路径分析, 这种方法允许放宽标准结构方程模型的一些限制, 包括数据非正态, 变量之间的非线性关系和小样本量[24]。首先构建包含筛选后所有变量的全模型, 然后逐步去除P值最高即不显著的路径, 直到模型的赤池信息量准则指标(AIC)不再降低, 选择此时的模型作为最优模型。该部分在R的“piecewiseSEM”包中完成。根据相关研究, 本文设置初始模型包含以下路径:(1)气候因子作用于土壤因子;(2)气候因子和土壤因子共同作用于植物多样性;(3)气候因子、土壤因子和植物多样性共同作用于地上生物量。
2 结果与分析 2.1 植物多样性与生物量间关系由图 2可知, 地上生物量与群落性状加权值、Shannon-Wiener指数呈显著的线性正相关, 拟合度分别为0.46(P < 0.001)和0.21(P = 0.001), 表明植物多样性对生产力存在明显的正向效应。

|
| 图 2 植物多样性和生物量的关系 Fig. 2 Relationship between plant diversity and biomass |
由表 2可知, 群落性状加权值和Shannon-Wiener指数均与年均降水量、土壤有机碳、土壤总氮呈线性正相关(均为P < 0.001), 均与年均气温呈线性负相关(P < 0.05、P < 0.001)。地上生物量与年均降水量、土壤有机碳、土壤总氮呈线性正相关(P < 0.001、P < 0.01、P < 0.01), 与年均气温关系不显著。土壤总磷与群落性状加权值、Shannon-Wiener指数、地上生物量间关系均不显著。
| 环境因子 Environmental factors |
回归分析 Regression analysis |
群落性状加权值 Community weighted mean trait value |
Shannon-Wiener指数 Shannon-Wiener index |
地上生物量 Aboveground biomass |
| 年均降雨量 | R2-线性 | 0.604 | 0.260 | 0.443 |
| Mean annual precipitation | P-线性 | 0.000 | 0.000 | 0.000 |
| 关系 | PL | PL | PL | |
| 年均温 | R2-线性 | 0.101 | 0.546 | 0.002 |
| Mean annual temperature | P-线性 | 0.025 | 0.000 | 0.303 |
| 关系 | NL | NL | NS | |
| 土壤有机碳 | R2-线性 | 0.405 | 0.369 | 0.146 |
| Soil organic carbon | P-线性 | 0.000 | 0.000 | 0.008 |
| 关系 | PL | PL | PL | |
| 土壤总氮 | R2-线性 | 0.333 | 0.363 | 0.143 |
| Soil total nitrogen | P-线性 | 0.000 | 0.000 | 0.009 |
| 关系 | PL | PL | PL | |
| 土壤总磷 | R2-线性 | 0.001 | 0.027 | 0.001 |
| Soil total phosphorus | P-线性 | 0.550 | 0.154 | 0.898 |
| 关系 | NS | NS | NS | |
| NS:无显著关系Non-significant relationship;PL:正线性关系Positive linear relationship;NL:负线性关系Negative linear relationship | ||||
采用层次分割的方法定量分析环境因子对植物多样性变化的解释率(图 3), 结果表明, 年均降水量为影响群落性状加权值的主要环境因子, 年均气温为影响Shannon-Wiener指数的主要环境因子。具体来看, 环境因子对群落性状加权值、Shannon-Wiener指数的解释率分别为72.2%、66.2%。年均降水量和土壤有机碳对群落性状加权值的解释率显著, 分别为40.9%、18.5%。年均气温、年均降水量和土壤有机碳对Shannon-Wiener指数的解释率显著, 分别为28.3%、15.4%、10.6%。土壤总氮对两者均有一定的解释作用但均不显著, 土壤总磷对两者有很小或没有解释作用。

|
| 图 3 环境因子对植物多样性的解释率 Fig. 3 Interpretation rate of environmental factors on plant diversity ** P < 0.05, * P < 0.01 |
同样的, 采用层次分割的方法定量分析环境因子和植物多样性对生物量变化的解释率(图 4), 结果表明, 所有因子共解释了生物量52.85%的变化。其中, 群落性状加权值和年均降水量为解释率较高的两个因子, 解释率分别为20.51%和17.37%, 而年均气温和三个土壤养分变量的解释率相对较小。

|
| 图 4 环境因子和植物多样性对生产力的解释率 Fig. 4 Interpretation rate of environmental factors and plant diversity on productivity |
本文进一步构建结构方程模型探究环境因子和植物多样性对生产力的复杂作用机制。综合植物多样性和生产力的影响因素层次分割分析结果, 初始模型包含了年均降水量、土壤有机碳、年均气温、群落性状加权值、Shannon-Wiener指数和地上生物量共6个变量。结果(图 5)表明, 模型共解释了地上生物量59%的变异。年均气温和年均降水量对地上生物量的直接效应不显著, 其主要通过间接路径作用于地上生物量。土壤有机碳通过影响群落性状加权值进而影响地上生物量而非直接作用。群落性状加权值和Shannon-Wiener指数均直接影响地上生物量, 其中, 群落性状加权值对地上生物量的影响显著, 起决定作用。此外, 群落性状加权值主要受年均降水量的正向影响, 而Shannon-Wiener指数主要受年均气温的负向影响。

|
| 图 5 环境因子和植物多样性对生产力影响的结构方程模型 Fig. 5 Structural equation model of the effects of environmental factors and plant diversity on productivity 实线和虚线分别表示变量间路径系数在统计上显著和不显著, 线的粗细表示变量间关系的强度大小, 以标准化路径系数衡量; ** P < 0.05, * P < 0.01 |
植物多样性和生产力与气候因子密切相关, 大尺度上水热因子的变化是导致陆地生态系统植被变化的主要原因。本研究发现, 年均降水量是影响旱区植物功能多样性(解释率40.9%)和地上生物量(解释率17.37%)变化的主要环境因子, 是影响物种多样性(解释率15.4%)变化的次要环境因子, 说明降水在决定旱区植物群落特征方面具有重要作用。普遍认为, 水分胁迫是旱地自然生态系统植物群落组成和生产力变化的决定因素[25—26], 而降水作为该地区最主要的水分来源, 其时空分布格局很大程度上决定了该区域生态系统的变化, 本文的研究结果佐证了此观点。同时, 年均气温是影响旱区植物物种多样性(解释率28.3%)的主要环境因子。年均气温与群落性状加权值和Shannon-Wiener指数呈负相关关系, 与孙小丽等[27]和白永飞等[28]在内蒙古荒漠草原的研究结果相同。这可能是由于在旱区, 降水量往往与气温呈负相关关系(图 6), 如敦煌极干旱地区气温较高但降水量极低, 内蒙古半干旱草原区气温较低而降水量较高。高温往往意味着更恶劣的生存环境, 从而表现为随着气温增加, 植物多样性反而降低的状况。

|
| 图 6 调查样点年均降水量与年均气温的关系 Fig. 6 Relationship between mean annual precipitation and mean annual temperature of sample plots |
在陆地生态系统中, 土壤和植被是两个相互依存、相互作用的因子[27]。本研究发现, 土壤有机碳和土壤总氮均与植物多样性和地上生物量显著正相关, 并且土壤有机碳的解释率高于土壤总氮, 而土壤总磷与植物多样性和地上生物量均无显著相关性且解释率极低。就碳氮元素而言, 土壤有机碳对于维持土壤结构和土壤肥力非常重要, 土壤氮含量与植物生物量积累直接相关[29], 故土壤碳氮含量的增加能够促进植物对养分的吸收, 进而提高群落生产力。杨崇曜等[30]对内蒙古地区的相关研究认为土壤全磷含量与物种多样性没有显著相关性, 本文的研究与之得到了相同的结果, 这可能与植物吸收利用的土壤磷含量相对碳氮而言较少有关[31]。从整体而言, 本研究发现土壤养分对植物多样性和生产力的解释作用明显小于气候因子, 这可能是因为土壤的大范围格局主要由气候因素所控制[32—33], 本文所构建的结构方程模型同样表明了这一点。
3.3 旱区生态系统气候、土壤养分、植物多样性对生产力的调控机制生产力是一个关键的生态系统功能指标, 许多不同的非生物和生物因素决定了生产力, 包括气候条件、土壤条件、群落结构、物种组成、物种多样性和功能特征等[34]。本文研究结果表明, 当综合考虑环境因子和生物因子后, 气候因子和土壤养分状况主要通过间接路径调控群落地上生物量, 而群落性状加权值对群落地上生物量具有显著的直接作用, 是决定群落地上生物量的最主要的因子。许多研究证明了群落性状加权值对生产力的重要作用, 如Xu等[7]认为群落性状加权值和功能离散度介导了水分添加和氮添加对群落生产力的影响, 朱羚等[35]认为群落性状加权值对内蒙古草原及其三个草原亚区的生产力具有最高的解释作用。这反映出, 植物功能多样性对于旱区生态系统生产力的维持具有十分重要的作用。目前研究认为, 多样性对生产力影响的两个基本机制是“互补”效应和“选择”效应, 而群落性状加权值突出了群落优势种的一些功能特征, 强调其对生产力的作用, 进而适用于揭示选择效应背景下, 功能多样性对于维持群落生产力的贡献大小[36—37]。因此, 本研究认为选择效应是旱区植物多样性对生产力维系的主导机制。这可能与旱区具有较强烈的环境过滤作用有关, 胁迫环境下, 性状和功能相近的物种被过滤筛选进入相近的生态位, 从而使群落内各物种的功能特征趋同[38]。也就是说, 在旱区生态系统由相对湿润到相对干旱的过程中, 不仅仅是物种数量导致生产力降低, 同时更重要的是关键功能性状的变化(在本研究中为更小的叶面积和比叶面积)导致了生产力的降低。
4 结论基于样带调查, 利用回归分析、层次分割和结构方程模型等方法, 系统分析了中国旱区生态系统植物多样性和生产力与环境因子间的复杂关系。旱区样带群落物种多样性和功能多样性变化的驱动因素不同, 功能多样性由降水主导, 物种多样性由气温主导, 相对来说土壤养分对物种多样性和功能多样性的影响较低。地上生物量受气候因子、土壤养分和植物多样性等因子的综合影响, 群落性状加权值、年均降水量和Shannon-Wiener指数对地上生物量具有较高的解释率。结构方程模型表明, 旱区样带群落地上生物量变化由功能多样性直接决定, 而气候和土壤养分通过影响群落功能多样性进而间接影响地上生物量, 体现了旱区选择效应的优势地位和环境过滤的重要性。本研究增强了对旱区植物群落与环境间关系的认识, 研究认为, 在气候变化背景下, 需综合考虑水、热因子对旱区植物群落的不同效应, 同时应充分认识到植物功能多样性在维持旱区生态系统生产力方面的重要作用。
| [1] |
Ali A, Lin S L, He J K, Kong F M, Yu J H, Jiang H S. Climate and soils determine aboveground biomass indirectly via species diversity and stand structural complexity in tropical forests. Forest Ecology and Management, 2019, 432: 823-831. DOI:10.1016/j.foreco.2018.10.024 |
| [2] |
Pan Q M, Symstad A J, Bai Y F, Huang J H, Wu J G, Naeem S, Chen D M, Tian D S, Wang Q B, Han X G. Biodiversity-productivity relationships in a natural grassland community vary under diversity loss scenarios. Journal of Ecology, 2022, 110(1): 210-220. DOI:10.1111/1365-2745.13797 |
| [3] |
Chu C J, Bartlett M, Wang Y S, He F L, Weiner J, Chave J, Sack L. Does climate directly influence NPP globally?. Global Change Biology, 2016, 22(1): 12-24. DOI:10.1111/gcb.13079 |
| [4] |
Harrison S P, Gornish E S, Copeland S. Climate-driven diversity loss in a grassland community. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(28): 8672-8677. DOI:10.1073/pnas.1502074112 |
| [5] |
Sun Y, Sun Y, Yao S R, Akram M A, Hu W G, Dong L W, Li H L, Wei M H, Gong H Y, Xie S B, Aqeel M, Ran J Z, Degen A A, Guo Q F, Deng J M. Impact of climate change on plant species richness across drylands in China: from past to present and into the future. Ecological Indicators, 2021, 132: 108288. DOI:10.1016/j.ecolind.2021.108288 |
| [6] |
刘晓琴, 张翔, 张立锋, 李英年, 赵亮, 徐世晓, 李红琴, 马荣荣, 牛犇, 高玉葆, 古松. 封育年限对高寒草甸群落组分和物种多样性的影响. 生态学报, 2016, 36(16): 5150-5162. |
| [7] |
Xu Z W, Li M H, Zimmermann N E, Li S P, Li H, Ren H Y, Sun H, Han X G, Jiang Y, Jiang L. Plant functional diversity modulates global environmental change effects on grassland productivity. Journal of Ecology, 2018, 106(5): 1941-1951. DOI:10.1111/1365-2745.12951 |
| [8] |
Ali A, Chen H Y H, You W H, Yan E R. Multiple abiotic and biotic drivers of aboveground biomass shift with forest stratum. Forest Ecology and Management, 2019, 436: 1-10. DOI:10.1016/j.foreco.2019.01.007 |
| [9] |
何楷迪, 孙建, 陈秋计. 气候要素和土壤质地对青藏高原草地净初级生产力和降水利用率的影响. 草业科学, 2019, 36(4): 1053-1065. |
| [10] |
Yang Y, Liu B R. Testing relationship between plant productivity and diversity in a desertified steppe in Northwest China. PeerJ, 2019, 7: e7239. DOI:10.7717/peerj.7239 |
| [11] |
Li X, Xiao J F. Global climatic controls on interannual variability of ecosystem productivity: similarities and differences inferred from solar-induced chlorophyll fluorescence and enhanced vegetation index. Agricultural and Forest Meteorology, 2020, 288/289: 108018. DOI:10.1016/j.agrformet.2020.108018 |
| [12] |
Ding Y, Zang R G. Determinants of aboveground biomass in forests across three climatic zones in China. Forest Ecology and Management, 2021, 482: 118805. DOI:10.1016/j.foreco.2020.118805 |
| [13] |
Li Z J, Li Z Y, Tong X Z, Zhang J H, Dong L, Zheng Y, Ma W H, Zhao L Q, Wang L X, Wen L, Dang Z H, Tuvshintogtokh I, Liang C Z, Li F Y. Climatic humidity mediates the strength of the species richness-biomass relationship on the Mongolian Plateau steppe. Science of the Total Environment, 2020, 718: 137252. DOI:10.1016/j.scitotenv.2020.137252 |
| [14] |
Ma W H, He J S, Yang Y H, Wang X P, Liang C Z, Anwar M, Zeng H, Fang J Y, Schmid B. Environmental factors covary with plant diversity-productivity relationships among Chinese grassland sites. Global Ecology and Biogeography, 2010, 19(2): 233-243. DOI:10.1111/j.1466-8238.2009.00508.x |
| [15] |
Duffy J E, Godwin C M, Cardinale B J. Biodiversity effects in the wild are common and as strong as key drivers of productivity. Nature, 2017, 549(7671): 261-264. DOI:10.1038/nature23886 |
| [16] |
Li C J, Fu B J, Wang S, Stringer L C, Wang Y P, Li Z D, Liu Y X, Zhou W X. Drivers and impacts of changes in China's drylands. Nature Reviews Earth & Environment, 2021, 2(12): 858-873. |
| [17] |
Maestre F T, Quero J L, Gotelli N J, Escudero A, Ochoa V, Delgado-Baquerizo M, García-Gómez M, Bowker M A, Soliveres S, Escolar C, García-Palacios P, Berdugo M, Valencia E, Gozalo B, Gallardo A, Aguilera L, Arredondo T, Blones J, Boeken B, Bran D, Conceição A A, Cabrera O, Chaieb M, Derak M, Eldridge D J, Espinosa C I, Florentino A, Gaitán J, Gatica M G, Ghiloufi W, Gómez-González S, Gutiérrez J R, Hernández R M, Huang X W, Huber-Sannwald E, Jankju M, Miriti M, Monerris J, Mau R L, Morici E, Naseri K, Ospina A, Polo V, Prina A, Pucheta E, Ramírez-Collantes D A, Romão R, Tighe M, Torres-Díaz C, Val J, Veiga J P, Wang D L, Zaady E. Plant species richness and ecosystem multifunctionality in global drylands. Science, 2012, 335(6065): 214-218. DOI:10.1126/science.1215442 |
| [18] |
杨永, 黄玥, 卫伟. 西北旱区典型灌草叶片性状沿气候梯度的变化. 生态学杂志, 2021, 40(12): 3769-3777. |
| [19] |
周丹, 沈彦俊, 陈亚宁, 郭英, 张勃. 西北干旱区荒漠植被生态需水量估算. 生态学杂志, 2015, 34(3): 670-680. |
| [20] |
García-Palacios P, Gross N, Gaitán J, Maestre F T. Climate mediates the biodiversity-ecosystem stability relationship globally. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(33): 8400-8405. DOI:10.1073/pnas.1800425115 |
| [21] |
Allen R G, Pereira L S, Raes D, Smith M J F, Rome. Crop evapotranspiration-Guidelines for computing crop water requirements-FAO Irrigation and drainage paper 56. 1998, 300(9): D05109.
|
| [22] |
Maestre F T, Quero J L, Gotelli N J, Escudero A, Ochoa V, Delgado-Baquerizo M, García-Gómez M, Bowker M A, Soliveres S, Escolar C, García-Palacios P, Berdugo M, Valencia E, Gozalo B, Gallardo A, Aguilera L, Arredondo T, Blones J, Boeken B, Bran D, Conceição A A, Cabrera O, Chaieb M, Derak M, Eldridge D J, Espinosa C I, Florentino A, Gaitán J, Gatica M G, Ghiloufi W, Gómez-González S, Gutiérrez J R, Hernández R M, Huang X W, Huber-Sannwald E, Jankju M, Miriti M, Monerris J, Mau R L, Morici E, Naseri K, Ospina A, Polo V, Prina A, Pucheta E, Ramírez-Collantes D A, Romão R, Tighe M, Torres-Díaz C, Val J, Veiga J P, Wang D L, Zaady E. Plant species richness and ecosystem multifunctionality in global drylands. Science, 2012, 335(6065): 214-218. DOI:10.1126/science.1215442 |
| [23] |
Ren T T, He N P, Liu Z G, Li M X, Zhang J H, Li A, Wei C Z, Lü X T, Han X G. Environmental filtering rather than phylogeny determines plant leaf size in three floristically distinctive plateaus. Ecological Indicators, 2021, 130: 108049. DOI:10.1016/j.ecolind.2021.108049 |
| [24] |
Valencia E, Maestre F T, Le Bagousse-Pinguet Y, Quero J L, Tamme R, Börger L, García-Gómez M, Gross N. Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands. New Phytologist, 2015, 206(2): 660-671. DOI:10.1111/nph.13268 |
| [25] |
He L, Li Z L, Wang X M, Xie Y W, Ye J S. Lagged precipitation effect on plant productivity is influenced collectively by climate and edaphic factors in drylands. Science of the Total Environment, 2021, 755: 142506. DOI:10.1016/j.scitotenv.2020.142506 |
| [26] |
Liu L, Zhang Y, Wu S, Li S, Qin D. Water memory effects and their impacts on global vegetation productivity and resilience. Scientific Reports, 2018, 8: 2962. DOI:10.1038/s41598-018-21339-4 |
| [27] |
孙小丽, 康萨如拉, 张庆, 常昌明, 马文静, 牛建明. 荒漠草原物种多样性、生产力与气候因子和土壤养分之间关系的研究. 草业学报, 2015, 24(12): 10-19. DOI:10.11686/cyxb2015014 |
| [28] |
白永飞, 李凌浩, 王其兵, 张丽霞, 张焱, 陈佐忠. 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究. 植物生态学报, 2000, 24(6): 667-673. DOI:10.3321/j.issn:1005-264X.2000.06.005 |
| [29] |
Chen D, Wei W, Chen L D. Effects of terracing on soil properties in three key mountainous regions of China. Geography and Sustainability, 2021, 2(3): 195-206. DOI:10.1016/j.geosus.2021.08.002 |
| [30] |
杨崇曜, 李恩贵, 陈慧颖, 张景慧, 黄永梅. 内蒙古西部自然植被的物种多样性及其影响因素. 生物多样性, 2017, 25(12): 1303-1312. DOI:10.17520/biods.2017140 |
| [31] |
曹娟, 闫文德, 项文化, 谌小勇, 雷丕锋, 向建林. 湖南会同不同年龄杉木人工林土壤磷素特征. 生态学报, 2014, 34(22): 6519-6527. |
| [32] |
Chen S P, Wang W T, Xu W T, Wang Y, Wan H W, Chen D M, Tang Z Y, Tang X L, Zhou G Y, Xie Z Q, Zhou D W, Shangguan Z, Huang J H, He J S, Wang Y F, Sheng J D, Tang L S, Li X R, Dong M, Wu Y, Wang Q F, Wang Z H, Wu J G, Chapin F S Ⅲ, Bai Y F. Plant diversity enhances productivity and soil carbon storage. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(16): 4027-4032. |
| [33] |
Luo Z K, Feng W T, Luo Y Q, Baldock J, Wang E L. Soil organic carbon dynamics jointly controlled by climate, carbon inputs, soil properties and soil carbon fractions. Global Change Biology, 2017, 23(10): 4430-4439. |
| [34] |
Li Y, Bao W K, Bongers F, Chen B, Chen G K, Guo K, Jiang M X, Lai J S, Lin D M, Liu C J, Liu X J, Liu Y, Mi X C, Tian X J, Wang X H, Xu W B, Yan J H, Yang B, Ma K P. Drivers of tree carbon storage in subtropical forests. Science of the Total Environment, 2019, 654: 684-693. |
| [35] |
朱羚, 金一兰, 丛日慧, 刘庆福, 丁勇, 张庆. 环境因素及种间竞争在群落多样性格局中的作用. 干旱区研究, 2018, 35(6): 1427-1435. |
| [36] |
Mahaut L, Fort F, Violle C, Freschet G T. Multiple facets of diversity effects on plant productivity: species richness, functional diversity, species identity and intraspecific competition. Functional Ecology, 2020, 34(1): 287-298. |
| [37] |
de Bello F, Lavorel S, Hallett L M, Valencia E, Garnier E, Roscher C, Conti L, Galland T, Goberna M, Májeková M, Montesinos-Navarro A, Pausas J G, Verdú M, E-Vojtkó A, Götzenberger L, Lepš J. Functional trait effects on ecosystem stability: assembling the jigsaw puzzle. Trends in Ecology & Evolution, 2021, 36(9): 822-836. |
| [38] |
许驭丹, 董世魁, 李帅, 沈豪. 植物群落构建的生态过滤机制研究进展. 生态学报, 2019, 39(7): 2267-2281. |