 2023, Vol. 43
2023, Vol. 43文章信息
- 李永刚, 张元明
- LI Yonggang, ZHANG Yuanming
- 干旱荒漠区不同藓结皮斑块碳通量对降雨量变化的响应
- Response of carbon flux of different moss biocrust patches to rainfall in arid desert
- 生态学报. 2023, 43(4): 1584-1595
- Acta Ecologica Sinica. 2023, 43(4): 1584-1595
- http://dx.doi.org/10.5846/stxb202107181930
-
文章历史
- 收稿日期: 2021-07-18
- 网络出版日期: 2022-10-14





干旱半干旱区占地球陆地面积的40%[1], 且储存全球25%的有机碳含量[2]。因此, 旱地土壤碳库的微小变化都会显著影响大气CO2浓度, 并进一步影响全球气候变暖, 在陆地生态系统碳循环中具有重要作用[3—5]。生物结皮是由细菌、真菌、蓝绿藻、地衣和苔藓植物等隐花植物类群与土壤表层颗粒胶结形成的有机复合体, 广泛分布在干旱荒漠区, 盖度往往高于维管束植物, 局部地区甚至能达到70%, 是荒漠土壤生物类群中最为活跃的组分之一[5—8]。例如, 生物结皮能改善土壤肥力、提高土壤抗风蚀、水蚀能力, 改变土壤养分循环, 影响土壤与大气之间的水分、气体交换等[9—12];此外, 生物结皮还能通过光合作用固定大气中的CO2, 增加土壤表层有机碳[12], 同时, 生物结皮也能通过自身所包含的有机成分与呼吸作用向大气中释放CO2进而影响旱地土壤碳循环[11—14]。因此, 生物结皮是旱地土壤碳循环的重要影响因素。
斑块最早是景观生态学中重要的概念, 是生态系统与周围环境在外貌或者性质上不同, 并具有一定内部均质性空间单元[15], 斑块对斑块内群落结构, 生产力和稳定性等具有重要影响。斑块内部群落结构的形成与维持, 是群落及其多样性的基础, 其驱动力往往源于外界干扰, 干扰会导致新的异质性生境和新群落斑块的形成[16—17]。已有斑块效应的研究多集中于具有较大尺度的森林、草原、荒漠草本植物群落, 近年来, 生物结皮的斑块效应愈发受到关注, 结皮斑块是生物土壤结皮的基本功能单位, 并作为一个微小的生态系统研究荒漠生物土壤结皮分布、生长以及多功能性等[18—19]。藓结皮斑块是生物结皮的重要形式, 通常是由单一物种的藓类植物发育成群, 以大小不同的沟壑将其分隔开, 最终以斑块状分布在荒漠土壤表面。研究发现, 斑块面积小于100 cm2的藓类结皮斑块数目占调查斑块总数的94%, 其面积约占调查斑块总面积的40%[20], 说明藓类结皮斑块以小型斑块为主, 但在面积上并不占据优势。藓类结皮斑块下层土壤全碳和全氮含量表现出中间高四周低, 而土壤全磷含量则表现出中间低四周高[21]。藓类结皮斑块微生物量碳、氮从中心向边缘呈现降低趋势[22]。随斑块面积的增加, 有助于维持藓类结皮斑块水分含量, 降低斑块内蒸散量, 增加藓类植物氮、磷含量, 同时改变齿肋赤藓斑块内土壤酶活性, 进而影响藓类结皮斑块内氮、磷养分的循环[21—24]。藓类结皮斑块大小能够显著的影响荒漠藓类结皮斑块内水分、养分等土壤特性, 而斑块面积大小是否能够显著的影响荒漠藓类结皮碳通量并不清楚。
在旱地生态系统中, 水分是荒漠生态系统植物生长的主要调控和限制因子, 还能够直接影响生物结皮的生理活性[25—27]。因此, 旱地土壤表面湿润时间的长短能够影响生物结皮的生理活性时间, 决定了生物结皮土壤的碳源-碳汇效应[25]。组成生物结皮的藻类、地衣及藓类植物是典型的变水植物对水分的变化极为敏感, 在水分频繁多变的条件下具有着很强的适应能力和环境抗性, 水分缺乏时生物结皮脱水进入休眠状态, 而在水分条件适宜的情况下能够快速恢复生理活性进行光合和呼吸作用[28—31]。一般情况下, 生物结皮的碳固定和释放很大程度上是与影响其活性的降雨事件相关, 主要体现无降雨供给时, 生物结皮表层受到干旱胁迫、土壤微生物处于休眠状态, 生物结皮土壤呼吸速率较低, 降雨发生时, 土壤水分增加会迅速激活生物结皮以及土壤微生物活性, 破坏土壤团聚体结构, 释放大量呼吸底物, 致使土壤呼吸速率迅速升高, 而受到大降雨事件时, 降雨造成土壤水分饱和或积水淹没生物结皮层, 阻碍氧气进入土壤, 从而抑制微生物活性, 降低生物土壤结皮的呼吸速率[13, 32—34], 同时降水事件导致的土壤有效水分持续时间是影响生物结皮呼吸速率的重要因素[35—36], 生物结皮的碳释放和固定过程也发生在降雨后的短时期内[34, 37—38]。研究发现, 降雨量的增加或降低能够显著的影响藓结皮水分入渗[39]、土壤微生物群落结构[36, 40], 藓类植物的光合呼吸速率[41], 进一步改变荒漠藓类结皮土壤的碳通量[42]。例如, 单次降雨量 < 2 mm时藓类结皮碳通量表现为碳排放, 光合碳固定量低于呼吸碳排放量;单次降雨量>5 mm时藓类结皮表现为碳固定, 光合碳固定量高于呼吸碳排放量[35, 43]。因此, 干旱区的降水模式也就决定了土壤碳吸收和释放之间的平衡[34, 35, 37—38], 从而影响了土壤与大气之间的碳交换通量。干旱区土壤水分主要来源于降雨, 认识土壤水分对土壤碳循环的影响规律是探讨土壤与大气圈碳交换对降雨响应机制的重要前提。
干旱区大多数降雨事件具有脉冲特点, 单次降雨量的不同对藓结皮碳通量已有较多研究, 但忽略了藓结皮斑块大小对其碳通量的影响, 尤其不同大小的藓结皮斑块碳通量对降雨量变化的响应并不清楚。本文以古尔班通古特沙漠的齿肋赤藓结皮斑块为研究对象, 通过施加不同降雨量处理, 连续测定降雨量处理后齿肋赤藓斑块表观碳通量和土壤体积含水量和温度, 比较并阐明斑块大小对荒漠藓结皮斑块表观碳通量的影响, 揭示藓类结皮斑块碳通量对荒漠降雨量变化的响应特征, 有助于加深理解荒漠藓类结皮斑块环境适应性特征, 为预测未来降水格局变化下荒漠藓结皮斑块组成、分布以及荒漠生态系统碳循环研究提供科学理论基础。
1 材料与方法 1.1 研究区自然概况研究区位于新疆准噶尔盆地中部的古尔班通古特沙漠(45°14′N, 87°36′E, 海拔540 m)。该沙漠气候属于我国温带干旱气候, 年均降水量仅70—160 mm, 潜在年蒸发量在2606.6 mm左右, 年均温7.26℃, 冬季平均气温在-16.8℃。沙漠腹地年均降水量为80 mm左右, 蒸发量却高达1600 mm以上, 多年平均气温为5—5.7℃。沙漠的基底是第四纪疏松冲积物, 主要由0.1—0.25 mm粒级的沙粒组成。沙丘的高度一般均在50 m以下, 沙丘主要是固定和半固定性质, 植被以白梭梭(Haloxylon persicum) 和梭梭(Haloxylon ammodendron)等小半乔木为建群种, 沙丘中下部有蛇麻黄(Ephedra distachya)、淡枝沙拐枣(Calligonum leucocladum)等灌木广泛分布[44], 春季丰富的降雨和早春融雪水使土壤含水量较高, 植物迅速萌发生长, 对节刺(Horanowia ulicina)、尖喙牻牛儿苗(Erodium oxyrrhynchum)、囊果苔草(Carex physodes)、角果藜(Ceratocarpus arenarius)等草本植物[45—46]。该地区生物土壤结皮(包括藻结皮、地衣结皮和藓类结皮)发育良好, 主要分布于丘间低地和灌丛下。
1.2 研究方法 1.2.1 样品采集与处理在古尔班通古特沙漠中心区域, 选择地势较为平坦、远离植物灌丛、荒漠藓类结皮斑块发育良好的丘间低地, 用直径为4.6、5.6、10 cm, 深度为10 cm的PVC管(PVC管为从市场中购买, 直径固定)在丘间低地随机选择直径约为4.6、5.6、10 cm的齿肋赤藓斑块。采集斑块时对齿肋赤藓斑块进行喷水处理, 以免取样时斑块表面出现裂缝使斑块破碎, 保证齿肋赤藓斑块的完整, 用PVC管插入齿肋赤藓斑块土壤中, 从侧面将PVC管取出用与底面积相同大小的铁片封底并用胶带粘贴确保铁片不会脱落和密闭。根据古尔班通古特沙漠年降雨数据, 发现该沙漠降雨量小于2 mm降雨为该沙漠的小降雨事件, 降雨≤2—5 mm的降雨事件占全年降雨量的87.5%, 占年全年降雨频次的47.5%, 降雨10—15 mm的降雨事件占全年降雨量的4.3%, 占全年降雨频次的26.6%, 因此, 本研究设置0、2、5、15 mm模拟降雨处理, 每个处理3个重复, 且土壤水分和温度的测定需要独立测定, 避免对碳通量的测定产生影响, 共采集72个齿肋赤藓斑块。
1.2.2 表观土壤碳通量的测定与计算实验在古尔班通古特沙漠中心区域进行, 通过收集的PVC管样品放置室外一周, 使PVC管内土壤水分含量降至最低。将所有PVC管放置室外对相同面积的齿肋赤藓斑块分别施加0 mm(对照, CK)、2 mm(降雨, PRE2)、5 mm(降雨, PRE5)、15 mm(降雨, PRE15)4个不同模拟降雨处理, 其中2 mm降雨为常规小降雨事件, 5 mm降雨为中等降雨事件, 15 mm降雨为极端大降雨事件。对所有PVC管样品进行降雨处理, 降雨处理于早上6点开始, 使用喷雾器进行喷洒, 使水滴均匀撒在PVC管内齿肋赤藓斑块表面, 确保无地表径流形成。施加不同降雨量处理完后, 分别在施加降雨后0、2、4、6、8、10、12、18、24、30、36、42、48、54 h测定静碳交换速率。测量时将连接有LI-6400(Li-Cor Inc., Lincoln, NE, USA)透明通量室, 在PVC管和通量室间添加凹槽, 确保测量过程中透明通量室内气体与外界完全隔离。选择自动模式, 测定时间为90 s, 每10 s记录1次, 共九次[39]。计算CO2的浓度变化率, 为了排除因透明通量室关闭对测定的影响, 选取通化室闭合后30 s到打开前10 s的CO2浓度数据, 同时剔除R2<0.95的数据, 减小偶然误差。计算公式如下:
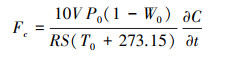
|
式中Fc是碳通量在单位面积上的速率(mmol CO2 m-2 s-1), 正值说明碳排放;V是测定时密闭空间的总体积(cm3), P0是测定时起初的气压值(kPa), W0是测定起初的水蒸气摩尔分数(mmol/mol), R是气体常数(8.312 kJ mol-1 K-1), S是测定时斑块面积(cm2), T0是空气温度(℃), 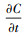
齿肋赤藓斑块内土壤体积含水量和土壤温度采用TDR测定, 测量土壤含水量和温度与齿肋赤藓斑块表观碳通量测定同时进行。为了避免TDR测量探头在测定土壤体积含水量和温度时对齿肋赤藓斑块表观碳通量测定的影响, 另选择36个齿肋赤藓斑块进行0 mm、2 mm、5 mm、15 mm 4个不同降雨量梯度处理, 将TDR探测器埋入每个PVC管内0—5 cm的土层中, 每隔10分钟进行记录1次, 测定PVC管内齿肋赤藓斑块内土壤体积含水量和土壤温度。
1.2.4 数据处理与分析利用方差分析比较不同降水强度和斑块大小对齿肋赤藓斑块累计碳通量间差异, 随后使用LSD进行多重比较。对降雨量和斑块大小进行重复测量双因素方差分析, 分析斑块大小和降雨量对齿肋赤藓斑块表观碳通量的影响。方差分解用于分析计算齿肋赤藓斑块大小和土壤体积含水量对齿肋赤藓斑块表观碳通量的解释。一般线性模型分析土壤体积含水量和齿肋赤藓斑块表观碳通量间关系, 所有的分析使用R 3.5.3。利用Origin 9.0作图分析、土壤体积含水量随时间的变化规律, 并对不同斑块面积的齿肋赤藓结皮54 h内的碳通量变化规律曲线进行积分计算, 得到累积碳通量。
2 结果与分析 2.1 不同斑块大小齿肋赤藓结皮斑块土壤体积含水量、温度间差异 2.1.1 不同斑块面积齿肋赤藓结皮斑块土壤体积含水量间差异如图 1所示, 不同降雨量处理后, 不同斑块大小的藓结皮土壤含水量差异显著, 土壤含水量随时间的变化趋势相似。直径为10 cm、5.6 cm和4.6 cm的藓结皮初始土壤含水量在2 mm降雨处理下分别为2.4%、3.9%、1.1%;5 mm降雨处理下分别为10.4%、5.4%、7.0%;15 mm降雨处理下分别为16.8%、19.2%、18.9%。不同大小的藓结皮斑块土壤含水量在不同降雨量处理下其变化趋势不同。在2 mm和5 mm降雨处理下, 土壤含水量呈现先增加后降低的变化趋势, 第4 h土壤含水量达到最大值。2 mm降雨, 直径10 cm、5.6 cm、4.6 cm藓结皮斑块土壤含水量分别在第12 h、54 h、36 h恢复到降雨处理前水平, 5 mm降雨, 直径10 cm藓结皮斑块在第54 h土壤含水量为1.9%未恢复至降雨处理前, 而直径5.6 cm和4.6 cm藓结皮斑块土壤含水量在第48 h土壤含水量恢复至处理前水平。15 mm降雨处理后土壤含水量呈现下降趋势, 10 cm、5.6 cm、4.6 cm直径斑块土壤体积含水量变化趋势相似, 且在第54 h土壤体积含水量分别为4.0%、3.5%、1.6%。

|
| 图 1 不同降雨处理后藓结皮斑块土壤含水量随时间的变化 Fig. 1 The changes of soil water content in moss crust patches with time after different rainfall treatments |
如图 2所示, 不同斑块面积土壤温度在不同降雨处理下变化趋势相同。0 mm、2 mm、5 mm、15 mm降雨处理, 10 cm直径齿肋赤藓斑块土壤温度的最低温分别是19.7℃、20.1℃、20.1℃、19.3℃, 最高温度为46.0℃、45.9℃、45.9℃、43.1℃;5.6 cm直径齿肋赤藓斑块土壤温度的最低温分别为20.2℃、20.0℃、19.5℃、19.4℃, 最高温分别为45.9℃、47.7℃、46.7℃、42.5℃;4.6 cm直径齿肋赤藓斑块土壤温度的最低温分别为20.0℃、20.1℃、18.7℃、19.2℃, 最高温分别为46.1℃、47.2℃、46.0℃、42.2℃。在15 mm降水处理后, 齿肋赤藓斑块温度显著低于0 mm、2 mm、5 mm降雨处理。

|
| 图 2 不同降雨处理后藓结皮斑块土壤温度随时间的变化 Fig. 2 The changes of soil temperature in moss crust patches with time after different rainfall treatments |
如图 3所示, 不同降雨量处理下藓结皮斑块碳通量随时间变化趋势明显不同。无降雨处理, 藓类结皮斑块净碳交换速率随时间呈现波动性变化, 表现为随时间净碳交换速率先升高后降低, 在第4 h、6 h(上午10点、12点)分别达到净碳交换速率的最大值, 第12 h(下午18点)达到该天净碳交换速率最低值, 碳交换速率在1.26和-1.51 mmol CO2 m-2 s-1间波动, 且随斑块面积增加其净碳交换速率波动范围增加。2 mm、5 mm降雨处理下藓结皮斑块净碳交换速率分别在第12 h、18 h达到最低, 随后趋于稳定。15 mm降雨处理下藓结皮斑块净碳交换速率的变化趋势与2 mm、5 mm降雨处理不同, 呈现降低-升高-降低-稳定的变化趋势。15 mm降雨处理, 直径为5.6 cm和4.6 cm的藓类结皮斑块净碳交换速率与10 cm的藓类结皮斑块净谈交换速率变化趋势不同。

|
| 图 3 不同降雨强度处理下荒漠藓类结皮斑块静碳交换速率随时间变化 Fig. 3 The changes of net carbon exchange rate of moss crust patches with time after different rainfall treatments |
降雨处理后, 不同斑块大小藓结皮碳通量变化明显不同, 直径10 cm的藓结皮斑块初始静碳交换速率显著(2 mm, P=0.010; 5 mm, P=0.045; 15 mm, P < 0.001)高于直径5.6 cm和4.6 cm藓结皮斑块。2 mm降雨处理下, 10 cm直径的藓结皮斑块净碳交换速率在第2 h—6 h显著高于5.6 cm和4.6 cm直径斑块;5 mm降雨处理下, 10 cm直径藓类结皮斑块净碳通量在2 h—10 h显著高于5.6 cm和4.6 cm直径;15 mm降雨处理下, 10 cm直径藓类结皮斑块净碳通量在整个测定过程均高于5.6 cm和4.6 cm直径藓结皮斑块, 且随时间的变化趋势明显不同。
如图 4所示, 藓结皮斑块54 h累积碳交换量在不同降雨量处理间差异显著, 随降雨量的增加藓结皮斑块累积碳交换量明显增加。直径10 cm藓结皮斑块累计谈交换量表现为15 mm降雨处理显著高于2 mm和5 mm降雨处理;5.6 cm和4.6 cm直径齿肋赤藓斑块累积碳交换量在15 mm降雨处理下表现为碳排放, 在0 mm、2 mm、5 mm降雨处理均表现为碳固定。不同斑块大小藓结皮碳通量间差异显著, 且随斑块面积增加呈增加趋势。15 mm降雨处理, 藓结皮斑块累积碳排放量均表现为碳释放, 10 cm直径斑块累积碳排放量显著高于5.6 cm和4.6 cm直径的斑块。2 mm和5 mm降雨处理下, 10 cm直径藓结皮斑块累积碳通量表现为碳排放, 而5.6 cm和4.6 cm直径藓结皮累积碳通量则表现为碳固定。无降雨处理的藓结皮斑块累积碳交换量表现为碳固定, 不同斑块大小的藓结皮累积碳固定无显著差异(P>0.05)。

|
| 图 4 不同降雨量处理后54 h荒漠藓结皮斑块累积碳交换量 Fig. 4 Accumulate carbon exchange of moss crust patches under different rainfall treatments for 54 hours 不同大写字母代表同一斑块面积藓结皮在不同降雨量下的累计碳交换量间差异显著(P < 0.05);不同小写字母代表同一降雨量下不同斑块面积藓结皮累计碳交换量间差异显著(P < 0.05) |
如表 1所示, 降雨量和斑块大小及交互效应均会显著影响藓结皮碳通量。藓结皮净碳交换速率与土壤含水量呈显著的正相关(P < 0.001, R2=0.26);不同斑块大小藓结皮净碳交换速率与土壤含水量间均呈显著(P < 0.001, R2=0.53;P < 0.001, R2=0.30;P=0.002, R2=0.17)正效应, 不同斑块大小藓结皮碳交换速率与土壤含水量间斜率具有明显差异(图 5)。藓结皮斑块在土壤温度小于25℃时, 随土壤温度升高净碳交换速率呈下降趋势, 而高于25℃时, 土壤温度对结皮斑块净碳交换速率无显著影响(图 6)。
| df | F | P | |
| 降雨量Rainfall amount | 3 | 50.60 | < 0.001 |
| 斑块大小Patch size | 2 | 56.22 | < 0.001 |
| 降雨量×斑块大小Rainfall amount × Patch size | 6 | 31.14 | < 0.001 |

|
| 图 5 藓结皮斑块净碳交换速率与土壤含水量间关系 Fig. 5 The relationship of the rate of net carbon exchange and the soil water content analyzed in the moss patch |

|
| 图 6 藓结皮斑块净碳交换速率与土壤温度间关系 Fig. 6 The relationship of the rate of net carbon exchange and the soil temperature analyzed in the moss patch |
本研究发现, 斑块大小能够显著影响藓结皮斑块碳通量。降雨处理后初始净碳交换速率表现为10 cm直径藓结皮斑块显著高于5.6 cm和4.6 cm直径藓结皮斑块, 且不同斑块面积藓结皮斑块净碳交换速率随时间的变化趋势明显不同, 这可能受到斑块内土壤含水量的变化的影响。前期研究发现, 藓结皮斑块大小能够显著影响斑块内土壤水分含量, 随斑块面积的增加斑块内土壤含水量呈现单峰模型[20]。斑块大小不仅能够显著的影响斑块内土壤含水量, 同时能够显著影响斑块内土壤酶活性[24, 47]。土壤水分和酶活性的改变可能导致不同斑块大小的藓结皮斑块内土壤微生物群落结构的改变, 影响土壤呼吸速率。10 cm直径藓结皮斑块累积净表观碳通量显著高于直径为5.6 cm和4.6 cm直径藓结皮斑块累积净表观碳通量, 这是因为斑块越大, 斑块内土壤含水量较高, 且能够维持较长时间的湿润状态, 有利于土壤微生物的生长, 产生大量的CO2气体排放。藓结皮斑块净碳交换速率与土壤含水量间呈显著的正相关, 且不同斑块大小其斜率不同, 表现为随斑块增加其净碳交换速率与土壤含水量间斜率增大。该结果说明, 随斑块面积增加土壤含水量对藓结皮斑块净碳通量的影响越大。因此, 荒漠藓结皮斑块越大越易受到荒漠水分的影响[48]。2 mm和5 mm降雨量能够显著影响10 cm直径藓结皮斑块碳通量由碳固定转变为碳释放, 而5.6 cm和4.6 cm直径的藓结皮斑块无显著影响。15 mm大降雨事件显著影响藓结皮斑块碳通量且均表现为碳释放, 随斑块面积的增加释放量增加, 大降雨事件不利于藓结皮斑块的碳固定, 小降雨事件仍维持小型藓结皮斑块碳固定。因此, 在小降雨脉冲事件频发的荒漠生态系统中, 小型藓结皮斑块有助于维持其生长和碳平衡, 避免造成由于长期湿润造成其碳损失, 降低齿肋赤藓植物死亡现象。
3.2 降雨对藓结皮斑块碳通量的影响土壤水分是限制干旱和半干旱地区生命活动的主要非生物因素, 降雨不仅是干旱区土壤水分的主要来源, 而且对地下微生物活性、土壤酶活性等具有重要的调控作用[49—50]。降雨量的变化会直接影响土壤水分有效性进而影响包括土壤呼吸在内的陆地生态系统碳循环。本研究结果表明, 不同降雨量处理能够显著的影响古尔班通古特沙漠中藓结皮斑块碳通量, 且斑块净碳交换速率随时间的变化趋势明显不同;随降雨量增加, 藓结皮斑块碳通量增加。这与前期研究结果一致, 即藓结皮斑块对降雨的响应表现为碳释放过程, 且随降雨量的增加碳排放量呈增加[51—52]。此外, 本研究发现, 2 mm降雨处理后仅在第12 h藓结皮斑块表观碳通量为正值(即表现为碳排放), 超过5 mm降雨处理下藓结皮斑块碳通量则呈负值(表现为碳固定)[35], 这与前期研究发现结果类似。不同降雨量脉冲对荒漠藓结皮植物含水量、光合、呼吸以及碳通量的影响存在显著差异[53—54]。无降雨处理时, 藓结皮能够利用凝结水激活其光合特性, 固定大气中的CO2, 干旱区生态系统表现为碳吸收过程, 但其净碳交换速率较低, 累积碳吸收量较低在0—1之间[55]。降雨事件发生后, 土壤微生物会迅速打破休眠从而恢复活性, 加速有机质等底物的分解和释放。如管超等[56]认为小于5 mm的降雨能刺激生物结皮中土壤微生物的新陈代谢活动, 增加其生物量, 从而提高土壤呼吸速率;肖波等[57]研究表明黄土高原地区小于6 mm降雨范围内藓结皮土壤呼吸速率随降雨量增加呈直线上升;韩海燕[58]在高寒沙区模拟1、2、5 mm降雨后发现藓结皮土壤呼吸速率较降雨前分别升高了11.0、6.2、4.1倍。长期干旱后, 随降雨量的增加, 土壤净碳交换速率激增程度升高。2 mm和5 mm降雨量下, 土壤净碳交换速率变化趋势由降雨初期的激增随后逐渐波动下降直至恢复到降雨处理前水平;而15 mm降雨量下, 土壤净碳交换速率在降雨初期仍出现明显的激增, 随后随时间先下降后缓慢上升, 可能受到藓结皮斑块植物生理活性(光合和呼吸作用)的影响。主要由于降雨处理后齿肋赤藓能够快速恢复光合呼吸作用, 因此降雨处理后会出现一段时间静碳交换速率的下降, 随着藓结皮斑块含水量的降低, 藓类植物脱水进入休眠状态, 藓结皮斑块碳通量逐渐进入稳定阶段, 并恢复到降雨前水平, 这一过程主要受到降雨量的影响[59]。
本研究还发现藓结皮斑块累积碳交换量随降雨量增加呈增加趋势, 可能原因是随降雨量增加, 提高了土壤含水量, 延长了藓结皮斑块湿润时间, 增加了齿肋赤藓光合和呼吸作用的时间, 且促进土壤微生物的生长, 进而释放大量的CO2[53, 60—62]。另一方面降雨处理前藓结皮斑块下层土壤处于干旱状态, 降雨处理后土壤含水量显著升高, 水分渗入填充土壤孔隙, 使土壤间隙中的CO2大量释放出来, 藓结皮由降雨前的碳吸收转变为释放碳[53, 62]。
3.3 不同降雨量下影响藓结皮斑块碳通量主要因素本研究结果表明斑块面积大小、降雨量及两者交互作用均显著改变了藓结皮斑块碳通量, 因此降雨量和斑块大小均作为影响藓结皮碳通量的重要因素。斑块面积大小能够影响藓类植物斑块内土壤含水量、蒸发量、凝结水形成等显著改变斑块内土壤含水量。降雨是土壤水分的主要来源, 土壤含水量能够促进土壤碳释放, 且在全球尺度上, 土壤呼吸随降雨量的增加呈线性增加[62—65]。因此, 降雨量和斑块大小通过影响斑块内土壤含水量进一步影响结皮斑块的碳通量[35, 40, 66]。这与本研究结果类似, 不同降雨量和斑块大小下, 土壤含水量对藓结皮斑块碳通量呈显著正效应, 因此, 土壤含水量是荒漠生态系统中藓结皮斑块碳通量的主要影响因素。除土壤水分之外, 土壤温度也是影响生物结皮土壤碳通量的重要环境因子[56]。前期研究发现土壤温度与荒漠藓类结皮表观碳通量间呈显著的正效应或负效应不同[67—71]。与前期的研究结果不同的是土壤温度对齿肋赤藓斑块表观碳通量影响存在温度阈值, 在低于25℃时, 土壤温度与藓结皮斑块碳通量间呈显著负相关, 高于25℃时, 温度与藓结皮碳通量间无显著的影响。有研究表明土壤温度的升高降低土壤水分的可利用性从而抑制了土壤呼吸, 影响藓类植物的光合呼吸作用能够显著的改变藓结皮斑块碳通量[35, 66, 72]。例如, 腾格里沙漠东南缘随土壤温度升高藓结皮和藻-地衣结皮的土壤呼吸速率呈下降趋势[61]。除了土壤含水量和土壤温度外, 斑块大小也可能通过改变土壤有机碳含量、土壤养分含量、土壤酶活性等土壤化学性质来进一步影响荒漠藓类结皮斑块表观碳通量。土壤养分和土壤有机质含量是土壤微生物呼吸的底物, 斑块面积越大其斑块下土壤有机质积累越多, 肥岛效应越明显[24, 47], 土壤释放CO2越高。土壤孔隙度、粗糙度、等土壤物理属性也可能影响土壤碳通量, 土壤孔隙中可能吸附CO2, 粗糙度等物理属性可能影响其水分入渗、土壤结构进一步影响土壤含水量和养分含量影响结皮斑块碳通量。
4 结论(1) 斑块大小能够显著影响藓结皮斑块碳通量, 斑块越小, 荒漠降雨脉冲对藓结皮碳通量的影响越小。
(2) 荒漠藓结皮斑块碳通量对降雨量的变化响应不同, 随降雨量增加藓结皮斑块碳通量和累积碳通量呈增加趋势。因此, 小型斑块对荒漠小降雨脉冲具有较强的适应性。
(3) 荒漠生态系统中, 不同降雨量和斑块大小下, 土壤含水量与藓类结皮斑块表观碳通量呈显著正相关, 因此, 土壤含水量是影响荒漠藓结皮斑块碳通量的主要因素。
| [1] |
Reynolds J F, Smith D M S, Lambin E F, Turner B L Ⅱ, Mortimore M, Batterbury S P J, Downing T E, Dowlatabadi H, Fernández R J, Herrick J E, Huber-Sannwald E, Jiang H, Leemans R, Lynam T, Maestre F T, Ayarza M, Walker B. Global desertification: building a science for dryland development. Science, 2007, 316(5826): 847-851. DOI:10.1126/science.1131634 |
| [2] |
Maestre F T, Escolar C, de Guevara M L, Quero J L, Lázaro R, Delgado-Baquerizo M, Ochoa V, Berdugo M, Gozalo B, Gallardo A. Changes in biocrust cover drive carbon cycle responses to climate change in drylands. Global Change Biology, 2013, 19(12): 3835-3847. DOI:10.1111/gcb.12306 |
| [3] |
Li B Z, Ge T D, Hill P W, Jones D L, Zhu Z K, Zhran M, Wu J S. Experimental strategies to measure the microbial uptake and mineralization kinetics of dissolved organic carbon in soil. Soil Ecology Letters, 2020, 2(3): 180-187. DOI:10.1007/s42832-020-0035-5 |
| [4] |
Chen H, Kong W D, Shi Q, Wang F, He C, Wu J S, Lin Q M, Zhang X Z, Zhu Y G, Liang C, Luo Y. Patterns and drivers of the degradability of dissolved organic matter in dryland soils on the Tibetan Plateau. Journal of Applied Ecology, 2022, 59(3): 884-894. DOI:10.1111/1365-2664.14105 |
| [5] |
Waring B G, Smith K R, Grote E E, Howell A, Reibold R, Tucker C L, Reed S C. Climatic controls on soil carbon accumulation and loss in a dryland ecosystems. Journal of Geophysical Research: Biogeosciences, 2021, 126(12): e2021JG006492. |
| [6] |
张元明, 曹同, 潘伯荣. 新疆古尔班通古特沙漠南缘土壤结皮中苔藓植物的研究. 西北植物学报, 2002, 22(1): 18-23. DOI:10.3321/j.issn:1000-4025.2002.01.003 |
| [7] |
李新荣, 张元明, 赵允格. 生物土壤结皮研究: 进展、前沿与展望. 地球科学进展, 2009, 24(1): 11-24. DOI:10.3321/j.issn:1001-8166.2009.01.002 |
| [8] |
Belnap J, Eldridge D. Disturbance and recovery of biological soil crusts. Ecological Studies. Berlin, Heidelberg: Springer Berlin Heidelberg, 2001: 363-383.
|
| [9] |
Rodriguez-Caballero E, Belnap J, Büdel B, Crutzen P J, Andreae M O, Pöschl U, Weber B. Dryland photoautotrophic soil surface communities endangered by global change. Nature Geoscience, 2018, 11(3): 185-189. DOI:10.1038/s41561-018-0072-1 |
| [10] |
张鹏, 李新荣, 张志山, 潘艳霞, 刘艳梅, 苏洁琼. 腾格里沙漠东南缘生物土壤结皮的固氮潜力. 应用生态学报, 2012, 23(8): 2157-2164. DOI:10.13287/j.1001-9332.2012.0295 |
| [11] |
Su Y G, Zhao X, Li A X, Li X R, Huang G. Nitrogen fixation in biological soil crusts from the Tengger desert, Northern China. European Journal of Soil Biology, 2011, 47(3): 182-187. DOI:10.1016/j.ejsobi.2011.04.001 |
| [12] |
Li X R, Zhang P, Su Y G, Jia R L. Carbon fixation by biological soil crusts following revegetation of sand dunes in arid desert regions of China: a four-year field study. Catena, 2012, 97: 119-126. DOI:10.1016/j.catena.2012.05.009 |
| [13] |
辜晨, 贾晓红, 吴波, 成龙, 杨占武, 杨德福, 赵雪彬. 高寒沙区生物土壤结皮覆盖土壤碳通量对模拟降水的响应. 生态学报, 2017, 37(13): 4423-4433. |
| [14] |
李玉强, 赵哈林, 赵玮, 云建英, 王少昆. 生物结皮对土壤呼吸的影响作用初探. 水土保持学报, 2008, 22(3): 106-109, 151. DOI:10.3321/j.issn:1009-2242.2008.03.022 |
| [15] |
邬建国, 李百炼, 伍业钢. 缀块性和缀块动态: Ⅰ.概念与机制. 生态学杂志, 1992, 11(4): 41-45. |
| [16] |
Bisigato A J, Bertiller M B, Ares J O, Pazos G E. Effect of grazing on plant patterns in arid ecosystems of Patagonian Monte. Ecography, 2005, 28(5): 561-572. DOI:10.1111/j.2005.0906-7590.04170.x |
| [17] |
Givnish T J, Volin J C, Owen V D, Volin V C, Muss J D, Glaser P H. Vegetation differentiation in the patterned landscape of the central Everglades: importance of local and landscape drivers. Global Ecology and Biogeography, 2008, 17(3): 384-402. DOI:10.1111/j.1466-8238.2007.00371.x |
| [18] |
Bowker M A, Maestre F T, Escolar C. Biological crusts as a model system for examining the biodiversity-ecosystem function relationship in soils. Soil Biology and Biochemistry, 2010, 42(3): 405-417. DOI:10.1016/j.soilbio.2009.10.025 |
| [19] |
Bowker M A, Maestre F T, Mau R L. Diversity and patch-size distributions of biological soil crusts regulate dryland ecosystem multifunctionality. Ecosystems, 2013, 16(6): 923-933. DOI:10.1007/s10021-013-9644-5 |
| [20] |
吉雪花, 张元明, 陶冶, 周小兵, 张静. 藓类结皮斑块面积与环境因子的关系. 中国沙漠, 2013, 33(6): 1803-1809. |
| [21] |
吉雪花, 张元明, 周小兵, 吴林, 张静. 不同尺度苔藓结皮土壤性状的空间分布特征. 生态学报, 2014, 34(14): 4006-4016. |
| [22] |
李茜倩, 张元明. 荒漠藓类结皮斑块中土壤理化性质、酶活性及微生物生物量分布的边缘效应. 生态学杂志, 2018, 37(7): 2114-2121. |
| [23] |
Yin B, Zhang Y M. Physiological regulation of Syntrichia caninervis Mitt. in different microhabitats during periods of snow in the Gurbantünggüt Desert, northwestern China. Journal of Plant Physiology, 2016, 194: 13-22. DOI:10.1016/j.jplph.2016.01.015 |
| [24] |
Li Y G, Zhou X B, Zhang Y M. Moss patch size and microhabitats influence stoichiometry of moss crusts in a temperate desert, Central Asia. Plant and Soil, 2019, 443(1/2): 55-72. |
| [25] |
Grote E E, Belnap J, Housman D C, Sparks J P. Carbon exchange in biological soil crust communities under differential temperatures and soil water contents: implications for global change. Global Change Biology, 2010, 16(10): 2763-2774. DOI:10.1111/j.1365-2486.2010.02201.x |
| [26] |
Büdel B, Williams W J, Reichenberger H. Annual net primary productivity of a cyanobacteria-dominated biological soil crust in the Gulf Savannah, Queensland, Australia. Biogeosciences, 2018, 15(2): 491-505. DOI:10.5194/bg-15-491-2018 |
| [27] |
Zhang C P, Niu D C, Song M L, Elser J J, Okie J G, Fu H. Effects of rainfall manipulations on carbon exchange of cyanobacteria and moss-dominated biological soil crusts. Soil Biology and Biochemistry, 2018, 124: 24-31. DOI:10.1016/j.soilbio.2018.05.021 |
| [28] |
国春晖, 沙伟, 李孝凯. 干旱胁迫对三种藓类植物生理特性的影响. 北方园艺, 2014(9): 78-82. |
| [29] |
吴楠, 魏美丽, 张元明. 生物土壤结皮中刺叶赤藓质膜透性对脱水、复水过程的响应. 自然科学进展, 2009, 19(9): 942-951. DOI:10.3321/j.issn:1002-008X.2009.09.008 |
| [30] |
吴玉环, 程佳强, 冯虎元, 安黎哲, 高谦, 程国栋. 耐旱藓类的抗旱生理及其机理研究. 中国沙漠, 2004, 24(1): 23-29. DOI:10.3321/j.issn:1000-694X.2004.01.004 |
| [31] |
严欢, 施亚峰, 韩正敏. 南京地区苔藓植物种类调查及耐旱性能分析. 林业调查规划, 2018, 43(4): 30-34. DOI:10.3969/j.issn.1671-3168.2018.04.007 |
| [32] |
Zhu M X, De Boeck H J, Xu H, Chen Z, Lv J, Zhang Z Q. Seasonal variations in the response of soil respiration to rainfall events in a riparian poplar plantation. The Science of the Total Environment, 2020, 747: 141222. DOI:10.1016/j.scitotenv.2020.141222 |
| [33] |
赵河聚, 成龙, 贾晓红, 吴波, 李元寿, 岳艳鹏, 周虹, 赵雪彬. 高寒沙区生物土壤结皮覆盖土壤碳释放动态. 生态学报, 2020, 40(18): 6396-6404. |
| [34] |
Yang Z L, Wei Y Y, Fu G Y, Song H Q, Li G Y, Xiao R. Asymmetric effect of increased and decreased precipitation in different periods on soil and heterotrophic respiration in a semiarid grassland. Agricultural and Forest Meteorology, 2020, 291: 108039. DOI:10.1016/j.agrformet.2020.108039 |
| [35] |
Wu L, Zhang Y M, Zhang J, Downing A. Precipitation intensity is the primary driver of moss crust-derived CO2 exchange: implications for soil C balance in a temperate desert of northwestern China. European Journal of Soil Biology, 2015, 67: 27-34. DOI:10.1016/j.ejsobi.2015.01.003 |
| [36] |
Li X J, Zhao Y, Yang H T, Zhang P, Gao Y P. Soil respiration of biologically-crusted soils in response to simulated precipitation pulses in the tengger desert, Northern China. Pedosphere, 2018, 28(1): 103-113. DOI:10.1016/S1002-0160(17)60307-2 |
| [37] |
赵洋, 齐欣林, 陈永乐, 张志山, 贾荣亮, 胡宜刚, 张鹏. 极端降雨事件对不同类型生物土壤结皮覆盖土壤碳释放的影响. 中国沙漠, 2013, 33(2): 543-548. |
| [38] |
Lee X H, Wu H J, Sigler J, Oishi C, Siccama T. Rapid and transient response of soil respiration to rain. Global Change Biology, 2004, 10(6): 1017-1026. DOI:10.1111/j.1529-8817.2003.00787.x |
| [39] |
Chamizo S, Cantón Y, Rodríguez-Caballero E, Domingo F, Escudero A. Runoff at contrasting scales in a semiarid ecosystem: a complex balance between biological soil crust features and rainfall characteristics. Journal of Hydrology, 2012, 452/453: 130-138. DOI:10.1016/j.jhydrol.2012.05.045 |
| [40] |
Cable J M, Huxman T E. Precipitation pulse size effects on Sonoran Desert soil microbial crusts. Oecologia, 2004, 141(2): 317-324. DOI:10.1007/s00442-003-1461-7 |
| [41] |
赵允格, 许明祥, Jayne Belnap. 生物结皮光合作用对光温水的响应及其对结皮空间分布格局的解译——以黄土丘陵区为例. 生态学报, 2010, 30(17): 4668-4675. |
| [42] |
Kim M, Or D. Hydration status and diurnal trophic interactions shape microbial community function in desert biocrusts. Biogeosciences, 2017, 14(23): 5403-5424. DOI:10.5194/bg-14-5403-2017 |
| [43] |
Baldauf S, Ladrón de Guevara M, Maestre F T, Tietjen B. Soil moisture dynamics under two rainfall frequency treatments drive early spring CO2 gas exchange of lichen-dominated biocrusts in central Spain. Peer J, 2018, 6: e5904. DOI:10.7717/peerj.5904 |
| [44] |
尹本丰, 张元明. 荒漠区不同微生境下齿肋赤藓对一次降雪的生理生化响应. 植物生态学报, 2014, 38(9): 978-989. |
| [45] |
张立运, 陈昌笃. 论古尔班通古特沙漠植物多样性的一般特点. 生态学报, 2002, 22(11): 1923-1932. |
| [46] |
钱亦兵, 吴兆宁, 张立运, 赵锐锋, 王小燕, 李有民. 古尔班通古特沙漠短命植物的空间分布特征. 科学通报, 2007, 52(19): 2299-2306. |
| [47] |
Li Y G, Zhou X B, Zhang Y M. Shrub modulates the stoichiometry of moss and soil in desert ecosystems, China. Journal of Arid Land, 2019, 11(4): 579-594. |
| [48] |
Borken W, Xu Y J, Davidson E A, Beese F. Site and temporal variation of soil respiration in European beech, Norway spruce, and Scots pine forests. Global Change Biology, 2002, 8(12): 1205-1216. |
| [49] |
Niu S L, Wu M Y, Han Y, Xia J Y, Li L H, Wan S Q. Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe. The New Phytologist, 2008, 177(1): 209-219. |
| [50] |
韩丹, 李玉霖, 杨红玲, 宁志英, 张子谦. 模拟增温和改变降雨频率对干旱半干旱区土壤呼吸的影响. 中国沙漠, 2021, 41(2): 100-108. |
| [51] |
Housman D C, Grote E E, Belnap J. Annual CO2 flux from a biological soil crust system on the Colorado Plateau: effects of increased temperature and summer precipitation. Ecological Society of America Annual Meeting Abstracts, 2007. |
| [52] |
Darrouzet-Nardi A, Reed S C, Grote E E, Belnap J. Patterns of longer-term climate change effects on CO2 efflux from biocrusted soils differ from those observed in the short term. Biogeosciences, 2018, 15(14): 4561-4573. |
| [53] |
Coe K K, Belnap J, Sparks J P. Precipitation-driven carbon balance controls survivorship of desert biocrust mosses. Ecology, 2012, 93(7): 1626-1636. |
| [54] |
Zhang J, Zhang Y M. Diurnal variations of chlorophyll fluorescence and CO2 exchange of biological soil crusts in different successional stages in the Gurbantunggut Desert of northwestern China. Ecological Research, 2014, 29(2): 289-298. |
| [55] |
Wilske B, Burgheimer J, Karnieli A, Zaady E, Andreae M O, Yakir D, Kesselmeier J. The CO2 exchange of biological soil crusts in a semiarid grass-shrubland at the northern transition zone of the Negev desert, Israel. Biogeosciences, 2008, 5(5): 1411-1423. |
| [56] |
管超, 张鹏, 李新荣. 腾格里沙漠东南缘生物结皮土壤呼吸对水热因子变化的响应. 植物生态学报, 2017, 41(3): 301-310. |
| [57] |
肖波, 郭成久, 赵东阳, 胡克林, 贾玉华. 黄土和风沙土藓结皮土壤呼吸对模拟降雨的响应. 生态学报, 2017, 37(11): 3724-3732. |
| [58] |
韩海燕. 高寒沙地生物结皮碳释放及其对土壤呼吸的影响[D]. 北京: 中国林业科学研究院, 2014.
|
| [59] |
贾晓红, 辜晨, 吴波, 李元寿, 成龙, 李新荣. 干旱沙区生物土壤结皮覆盖土壤CO2通量对脉冲式降雨的响应. 中国沙漠, 2016, 36(2): 423-432. |
| [60] |
Gordon H, Haygarth P M, Bardgett R D. Drying and rewetting effects on soil microbial community composition and nutrient leaching. Soil Biology and Biochemistry, 2008, 40(2): 302-311. |
| [61] |
Miralles I, Lázaro R, Sánchez-Marañón M, Soriano M, Ortega R. Biocrust cover and successional stages influence soil bacterial composition and diversity in semiarid ecosystems. Science of the Total Environment, 2020, 709: 134654. |
| [62] |
Zhao Y, Zhang P. Rainfall characteristics determine respiration rate of biological soil crusts in drylands. Ecological Indicators, 2021, 124: 107452. |
| [63] |
Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B: Chemical and Physical Meteorology, 1992, 44(2): 81-99. |
| [64] |
Belnap J, Weber B, Büdel B. Biological soil crusts as an organizing principle in drylands. Springer International Publishing Switzerland, 2016. |
| [65] |
吴林, 苏延桂, 张元明. 模拟降水对古尔班通古特沙漠生物结皮表观土壤碳通量的影响. 生态学报, 2012, 32(13): 4103-4113. |
| [66] |
Yao X M, Bowker M A, Xiao B. Estimation of annual CO2 efflux of moss biocrust through measuring and simulating its respiration rate in a semiarid climate. Geoderma, 2020, 376: 114560. |
| [67] |
Luo C Y, Xu G P, Chao Z G, Wang S P, Lin X W, Hu Y G, Zhang Z H, Duan J C, Chang X F, Su A L, Li Y N, Zhao X Q, Du M Y, Tang Y H, Kimball B. Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan Plateau. Global Change Biology, 2010, 16(5): 1606-1617. |
| [68] |
徐冰鑫, 胡宜刚, 张志山, 陈永乐, 张鹏, 李刚. 模拟增温对荒漠生物土壤结皮-土壤系统CO2、CH4和N2O通量的影响. 植物生态学报, 2014, 38(8): 809-820. |
| [69] |
Thomas A D, Hoon S R, Dougill A J. Soil respiration at five sites along the Kalahari Transect: effects of temperature, precipitation pulses and biological soil crust cover. Geoderma, 2011, 167/168: 284-294. |
| [70] |
Darrouzet-Nardi A, Reed S C, Grote E E, Belnap J. Observations of net soil exchange of CO2 in a dryland show experimental warming increases carbon losses in biocrust soils. Biogeochemistry, 2015, 126(3): 363-378. |
| [71] |
Tucker C L, Ferrenberg S, Reed S C. Climatic sensitivity of dryland soil CO2 fluxes differs dramatically with biological soil crust successional state. Ecosystems, 2019, 22(1): 15-32. |
| [72] |
Garcia-Pichel F, Belnap J. Microenvironments and microscale productivity of cyanobacterial desert crusts1. Journal of Phycology, 1996, 32(5): 774-782. |