 2023, Vol. 43
2023, Vol. 43文章信息
- 杨光辉, 秦树林, 金光泽
- YANG Guanghui, QIN Shulin, JIN Guangze
- 小兴安岭五种林型早春草本植物多样性及其环境解释
- Early-spring herb diversity and its environmental interpretation of five forest types in Xiaoxing'an Mountains, China
- 生态学报. 2023, 43(3): 1234-1246
- Acta Ecologica Sinica. 2023, 43(3): 1234-1246
- http://dx.doi.org/10.5846/stxb202201260250
-
文章历史
- 收稿日期: 2022-01-26
- 网络出版日期: 2022-10-10





2. 吉林省林业调查规划院, 长春 130022;
3. 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040;
4. 东北林业大学东北亚生物多样性研究中心, 哈尔滨 150040
2. Jilin Provincial Institute of Forest Inventory and Planning, Changchun 130022, China;
3. Key Laboratory of Sustainable Forest Ecosystem Management of Ministry of Education, Northeast Forestry University, Harbin 150040, China;
4. Northeast Asia Biodiversity Research Center, Northeast Forestry University, Harbin 150040, China
生物多样性是人类赖以生存的物质基础, 对于维持生态系统功能和稳定性[1]、提高生态效益[2]具有重要作用。近年来人类活动的增加导致生物多样性正在迅速下降[3], 其生态后果已受到广泛关注[4]。草本植物是森林生态系统生物多样性的主要组成部分[5], 探究草本植物组成与多样性对人为活动的响应为实现生物多样性的保护提供理论依据。早春时期生长的草本植物通过对土壤氮元素和钾元素的大量积累, 作为短期的营养汇, 避免了森林土壤养分流失并为养分储存做出重要贡献[6], 除此之外, 其易于分解的植物体促进森林生态系统的物质循环, 提高森林生产力, 并能够作为药用以及食用植物, 具有极高的经济和生态价值[7]。通过对早春特殊环境的长期适应, 早春草本层植物进化出与生境相适的生活史对策, 可分为三类:(1)早春生长期植物:早春时期开始生长, 开花和结实等生活史阶段在林分郁闭之前完成, 林分郁闭后进入休眠期;(2)早春开花植物:早春时期进行生长、开花和结实等生活史阶段, 在林分郁闭后仍可继续生长;(3)早春展叶植物:生长过程在早春完成, 林分郁闭后进行开花和结实的这类植物[7]。近年来就早春草本层植物的种类和群落特征[8]、生物量[9]等方面进行了研究, 但对不同生活史对策早春草本层植物的多样性及其影响因素方面的研究较少[10], 了解其多样性变化特征及影响因素对于保护、开发以及利用早春草本层植物资源具有重要意义。
林下草本植物群落的组成及多样性随干扰条件[11]和方式[12]不同而产生差异。人为干扰导致的林分改变影响了包括林分组成和结构[13]、土壤[14]、生物多样性[15]等多种生态系统要素。在森林生态系统中, 上层植物是森林资源的主要获取组分, 森林管理导致的树种改变, 通过地球化学循环的动态变化, 明显改变了土壤的化学性质[16], 而土壤养分又是草本物种分异的主要环境因子, 对草本植物分布起主导作用[17], 对其生长发育及其多样性产生直接影响[18]。同时, 冠层通过控制太阳能的截获量来调节林下光照和水分[19], 进而使林下草本植物丰富度和多样性存在差异[20]。冠层特征和土壤特征变化均会对草本植物物种组成和多样性产生显著影响, 从而使不同林型草本植物群落生物多样性间的差异增大。因此, 探究林分类型改变后林下草本植物组成与多样性特征及其影响因素仍十分必要。
东北东部山区的阔叶红松林在20世纪初遭到大面积工业化采伐, 除有小部分还保存有原始的红松林外, 其他均被次生林和人工林替代, 其完整的原始林-次生林-人工林的干扰类型梯度[21], 为探索森林生态系统经过人为干扰或经营后草本植物多样性的差异提供了良好的研究平台。本研究以黑龙江凉水国家级自然保护区内的白桦次生林、红松人工林、落叶松人工林、阔叶红松择伐林以及阔叶红松林为研究对象, 探究五种林型林下早春草本层植物组成及多样性之间的差异以及对环境的响应, 为进一步研究干扰对小兴安岭乃至东北地区森林的生态系统的生物多样性的影响以及其保护提供理论依据。
1 材料和方法 1.1 研究区概况研究区位于黑龙江省伊春市大箐山县的黑龙江凉水国家级自然保护区(47°10′50″N, 128°53′20″E)内。该保护区资源丰富、结构复杂, 植被覆盖率高达96%, 地带性植被是以红松为主的温带针阔混交林。研究区具有明显的温带大陆性季风气候特征, 春季干燥、多大风, 夏季降水量大, 集中在6—8月, 秋季降温剧烈, 冬季严寒、多风雪。年均温-0.3℃, 年平均降水量676 mm, 年平均相对湿度78%, 无霜期100—120 d, 积雪期130—150 d。保护区属于低山丘陵地带, 海拔高度在280—707.3 m之间, 平均海拔高度在400 m左右。地带性土壤为暗棕壤, 占保护区总面积的84.91%。样地林型概况见表 1。
| 林型 Forest type |
乔木组成 Tree composition |
灌木组成 Shrub composition |
利用历史 Land-use history |
林龄 Age/a |
| 白桦次生林 Secondary birch forest |
4落叶松3白桦1红皮云杉1春榆+五角枫+大青杨+红松-紫椴-大黄柳-黄檗-臭冷杉-水曲柳-辽东桤木-花楷槭-裂叶榆-香杨-蒙古栎-稠李-硕桦-辽椴-青楷槭-花楸树-朝鲜槐 | 8暴马丁香1毛榛+瘤枝卫矛+金花忍冬-东北山梅花-刺五加-鼠李-接骨木-修枝荚蒾-光萼溲疏-狗枣猕猴桃-榆叶梅-楤木 | 1953年皆伐, 自然更新成林 | 68 |
| 红松人工林 Korean pine plantation |
6红松1白桦1落叶松+黄檗+红皮云杉+水曲柳-鱼鳞云杉-紫椴-五角枫-春榆-花楷槭-稠李-青楷槭-裂叶榆-胡桃楸 | 4暴马丁香4毛榛1光萼溲疏+榆叶梅+金花忍冬+瘤枝卫矛+刺五加-狗枣猕猴桃 | 1954年人工造林 | 67 |
| 落叶松人工林 Dahurian larch plantation |
8落叶松1水曲柳+红松+五角枫+紫椴-红皮云杉-青楷槭-花楷槭-稠李-辽东桤木-裂叶榆-硕桦 | 4毛榛2瘤枝卫矛2刺五加1东北山梅花+暴马丁香+金花忍冬-光萼溲疏 | 1954年人工造林 | 67 |
| 阔叶红松择伐林 Selectively cut mixed broadleaved-Korean pine forest |
4红松1五角枫1硕桦1水曲柳1青楷槭1裂叶榆1臭冷杉+春榆+紫椴+山杨+蒙古栎-黄檗-花楷槭-辽东桤木-大黄柳-稠李-鱼鳞云杉-红皮云杉 | 9暴马丁香+毛榛-瘤枝卫矛-东北山梅花-刺五加-楤木-鼠李-狗枣猕猴桃-金花忍冬-光萼溲疏-修枝荚蒾 | 1971年择伐, 强度30% | >200 |
| 阔叶红松林 Mixed broadleaved-Korean pine forest |
6红松1五角枫1紫椴1硕桦+水曲柳+臭冷杉+青楷槭+榆树+裂叶榆-白桦-稠李-山杨-花楷槭-辽东桤木-春榆-辽椴 | 5斑叶稠李4毛榛1暴马丁香-东北山梅花-瘤枝卫矛 | 原始林 | >300 |
| 暴马丁香(Syringa reticulata subsp. amurensis)、稠李(Padus avium)、春榆(Ulmus davidiana var. japonica)、刺五加(Eleutherococcus senticosus)、大黄柳(Salix raddeana)、大青杨(Populus ussuriensis)、东北山梅花(Philadelphus schrenkii)、硕桦(Betula costata)、狗枣猕猴桃(Actinidia kolomikta)、光萼溲疏(Deutzia glabrata)、红皮云杉(Picea koraiensis)、胡桃楸(Juglans mandshurica)、花楷槭(Acer ukurunduense)、花楸树(Sorbus pohuashanensis)、黄檗(Phellodendron amurense)、金花忍冬(Lonicera chrysantha)、接骨木(Sambucus williamsii)、辽椴(Tilia mandshurica)、臭冷杉(Abies nephrolepis)、裂叶榆(Ulmus laciniata)、瘤枝卫矛(Euonymus verrucosus)、楤木(Aralia elata)、辽东桤木(Alnus hirsuta)、毛榛(Corylus mandshurica)、蒙古栎(Quercus mongolica)、修枝荚蒾(Viburnum burejaeticum)、青楷槭(Acer tegmentosum)、朝鲜槐(Maackia amurensis)、斑叶稠李(Padus maackii)、鼠李(Rhamnus davurica)、山杨(Populus davidiana)、水曲柳(Fraxinus mandshurica)、五角枫(Acer pictum subsp. mono)、香杨(Populus koreana)、鱼鳞云杉(Picea jezoensis var. microsperma)、榆叶梅(Prunus triloba)、紫椴(Tilia amurensis)、红松(Pinus koraiensis)、落叶松(Larix gmelinii)、白桦(Betula platyphylla) | ||||
在每个林型中设置3个20 m×30 m的样方, 在每个样方中按照“X”型设立5个1 m×1 m的草本样方。于2021年5月初进行林下草本植物多样性调查。鉴定林下草本植物的种类, 测定物种个体数(以地上部分单株进行统计)、高度和盖度, 盖度采用网格法测定, 即将1 m×1 m的小样方分为100个10 cm×10 cm的小网格并用目测法估算盖度。
以每个1 m×1 m的草本小样方为采样单位, 在其周围随机选取3个采样点, 在每个采样点取表层(0—10 cm)土壤样品进行混合, 带回实验室进行分析测定。使用pH计(HANNA pH211, Italy)测定土壤pH, 使用碳氮元素分析仪(Analytic Jena AG, Jena, Germany)进行土壤有机碳(Soil organic carbon)的测定, 使用全自动连续分析仪(SEAL AA3)测定土壤的全氮(Total nitrogen, TN)和全磷(Total phosphorus, TP)。
选择阴天或晴好天气的日出或日落, 将连接180 °鱼眼镜头(Nikon FC-E8)的数码相机(Coolpix 4500, Nikon, Tokyo, Japan)利用三角架水平固定在距地面1.3 m处, 采集冠层半球面影像。拍照点与草本样方调查点保持一致, 每个样方拍摄三张照片, 采用Gap Light Analyzer 2.0(GLA)图像处理软件处理林冠影像照片[19, 22], 可直接获得林下直射光(Transmitted direct light, TDir)、林下散射光(Transmitted diffused light, TDif)和林下总光照(Transmitted total light, TTot)的数据。
1.3 数据分析 1.3.1 重要值计算林下草本植物的重要值计算公式为:重要值(Pi)= (相对多度+相对盖度+相对高度)/3[23];
1.3.2 群落百分比相似系数计算群落百分比相似系数:

|
式中, xij和xik分别表示两个不同群落中各物种的重要值。
1.3.3 物种多样性指数计算林下草本植物多样性测定采用物种丰富度指数(S)以及Shannon-Wiener多样性指数。具体计算公式如下:
Shannon-Wiener多样性指数H′:
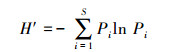
|
式中, S为群落中某生活史对策中的物种总数;Pi为样方内第i种物种或某生活史对策的重要值。
1.3.4 统计分析方法五种林型不同生活史对策植物的盖度、密度及物种多样性数据通过SPSS 26.0软件进行Levene同质性检验、单因素方差分析(One-Way ANOVA), 对于方差齐性的统计量采用LSD多重比较获得结果(P=0.05);对于方差不齐的统计量采用Tamhane法进行比较。对数据进行lg转换, 使用R.4.0.2中的hier.part包[24]对不同生活史对策草本植物的密度、盖度和物种多样性与土壤因子和光环境因子进行层次分割, 对早春草本层植物组成及其多样性产生显著影响的因子进行一般线性模型分析, 为达到最佳的拟合效果, 对二者分别进行线性和非线性拟合, 选取拟合效果最佳的进行绘图。所有图均在Origin 2019中进行绘制。
2 结果 2.1 五种林型早春草本层植物组成研究区林下草本物种丰富, 在五种林型下共发现草本植物60种, 隶属于26科52属(附表 1)。其中白桦次生林20科37属43种, 红松人工林21科36属38种, 落叶松人工林22科33属38种, 阔叶红松择伐林19科28属32种, 阔叶红松林22科34属37种。
五种林型下不同生活史对策的草本植物的科、属组成差异较小, 早春生长期植物在各林下均为7种;早春开花植物在红松人工林物种数最高(10种), 占比26.3%;早春展叶植物在白桦次生林中最高(27种), 占比62.8%(表 2)。
| 林型 Forest type |
早春生长期植物 Early-spring growing herbs |
早春开花植物 Early-spring flowering herbs |
早春展叶植物 Early-spring foliating herbs |
||||||||
| 科/% Family |
属/% Genus |
种/% Species |
科/% Family |
属/% Genus |
种/% Species |
科/% Family |
属/% Genus |
种/% Species |
|||
| 白桦次生林 Secondary birch forest |
3 (15%) |
7 (18.9%) |
7 (16.3%) |
8 (40%) |
9 (24.3%) |
9 (20.9%) |
12 (60%) |
22 (59.5%) |
27 (62.8%) |
||
| 红松人工林 Korean pine plantation |
3 (11.1%) |
7 (19.4%) |
7 (18.4%) |
9 (42.9%) |
10 (27.8%) |
10 (26.3%) |
14 (66.7%) |
20 (55.6%) |
21 (55.3%) |
||
| 落叶松人工林 Dahurian larch plantation |
3 (13.6%) |
7 (21.2%) |
7 (18.4%) |
8 (36.4%) |
9 (27.3%) |
9 (23.7%) |
15 (68.2%) |
18 (54.5) |
22 (57.9%) |
||
| 阔叶红松择伐林 Selectively cut mixed broadleaved- Korean pine forest |
4 (21.1%) |
7 (25.0%) |
7 (21.9%) |
7 (36.8%) |
7 (25.0%) |
7 (21.9%) |
12 (63.2%) |
15 (53.6%) |
18 (56.3%) |
||
| 阔叶红松林 Mixed broadleaved-Korean pine forest |
4 (18.2%) |
7 (20.6%) |
7 (18.9%) |
7 (31.8%) |
8 (23.5%) |
8 (21.6%) |
15 (68.2%) |
20 (58.8%) |
22 (59.5%) |
||
白桦次生林和红松人工林均以银莲花、蚊子草(Filipendula palmata)和东北羊角芹为林下草本优势种, 落叶松人工林的林下优势种为林金腰(Chrysosplenium lectus-cochleae)、酢浆草(Oxalis corniculata)和荷青花(Hylomecon japonica), 阔叶红松择伐林的优势种为东北羊角芹、银莲花和荷青花, 阔叶红松林的优势种为银莲花、荷青花和拟扁果草;银莲花、东北羊角芹和荷青花在五种林型下均出现且占据优势地位, 华北剪股颖(Agrostis clavata)、木贼(Equisetum hyemale)、唢呐草(Mitella nuda)、乌头(Aconitum carmichaeli)和狭叶荨麻(Urtica angustifolia)仅在单一林分中占据重要值的前十位(表 3)。五种林型早春草本层植物组成相似度均高于50%, 其中阔叶红松林和阔叶红松择伐林最高(68.37%), 阔叶红松林和红松人工林最低(51.46%)(表 4)。
| 物种 Species |
重要值Importance value/% | ||||
| 白桦次生林 Secondary birch forest |
红松人工林 Korean pine plantation |
落叶松人工林 Dahurian larch plantation |
阔叶红松择伐林 Selectively cut mixed broadleaved-Korean pine forest |
阔叶红松林 Mixed broadleaved-Korean pine forest |
|
| 白花碎米荠Cardamine leucantha | — | — | 4.1 | 7.7 | 3.8 |
| 荷青花Hylomecon japonica | 3.5 | 4.9 | 8.6 | 9.5 | 10.2 |
| 华北剪股颖Agrostis clavata | 4.2 | — | — | — | — |
| 林金腰Chrysosplenium lectus-cochleae | 5.7 | 6.4 | 12.0 | 7.5 | — |
| 龙常草Diarrhena mandshurica | 6.6 | — | — | — | 3.6 |
| 毛缘薹草Carex pilosa | — | — | — | 5.5 | 4.9 |
| 细叶孩儿参Pseudostellaria sylvatica | 4.5 | 4.5 | 3.7 | 3.0 | — |
| 木贼Equisetum hyemale | — | — | — | — | 5.4 |
| 拟扁果草Enemion raddeanum | — | 4.0 | 8.4 | 9.4 | 7.8 |
| 唢呐草Mitella nuda | — | 3.2 | — | — | — |
| 蚊子草Filipendula palmata | 11.1 | 8.4 | 7.8 | 4.3 | — |
| 乌头Aconitum carmichaelii | — | 3.5 | — | — | — |
| 五福花Adoxa moschatellina | 5.7 | — | — | — | — |
| 狭叶荨麻Urtica angustifolia | — | — | 4.1 | — | — |
| 东北羊角芹Aegopodium alpestre | 6.8 | 10.8 | 6.4 | 14.5 | 5.2 |
| 延胡索Corydalis yanhusuo | 2.8 | — | — | 3.4 | 4.2 |
| 银莲花Anemone cathayensis | 17.4 | 12.6 | 4.7 | 13.0 | 29.6 |
| 酢浆草Oxalis corniculata | — | 7.6 | 11.7 | — | 4.9 |
| 其他Others | 31.8 | 34.1 | 28.5 | 22.4 | 20.3 |
| 林型Forest type | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ |
| Ⅰ | — | 64.08 | 52.96 | 54.16 | 53.12 |
| Ⅱ | 61.73 | 63.18 | 51.46 | ||
| Ⅲ | 60.76 | 52.58 | |||
| Ⅳ | 68.37 | ||||
| Ⅴ | — | ||||
| Ⅰ: 白桦次生林Secondary birch forest; Ⅱ: 红松人工林Korean pine plantation; Ⅲ: 落叶松人工林Dahurian larch plantation; Ⅳ: 阔叶红松择伐林Selectively cut mixed broadleaved-Korean pine forest; Ⅴ: 阔叶红松林Mixed broadleaved-Korean pine forest | |||||
阔叶红松林的早春生长期植物密度和盖度最高, 密度显著高于红松人工林和落叶松人工林样地(P<0.05), 盖度显著高于除白桦次生林外的其他三种林型(P<0.05);落叶松人工林的早春开花植物密度显著高于白桦次生林、阔叶红松择伐林和阔叶红松林(P<0.05), 红松人工林的密度显著高于阔叶红松林(P<0.05);早春展叶植物的密度和盖度在五个林型之间没有显著差异(P>0.05)(图 1)。

|
| 图 1 五种林型不同生活史对策的草本植物密度和盖度的差异(平均值±标准误差) Fig. 1 Differences of herbaceous plants density and coverage under different life-history strategies in five forest types (mean ± SE) 不同小写字母表示同一生活史对策不同林型间差异显著(P<0.05), 不同大写字母表示同一林型不同生活史对策间差异显著(P<0.05); Ⅰ: 白桦次生林Secondary birch forest; Ⅱ: 红松人工林Korean pine plantation; Ⅲ: 落叶松人工林Dahurian larch plantation; Ⅳ: 阔叶红松择伐林Selectively cut mixed broadleaved-Korean pine forest; Ⅴ: 阔叶红松林Mixed broadleaved-Korean pine forest |
白桦次生林的早春展叶植物密度显著高于早春开花植物(P<0.05), 盖度表现为早春生长期植物和早春展叶植物显著高于早春开花植物(P<0.05);落叶松人工林的早春开花植物密度显著高于早春生长期植物(P<0.05);阔叶红松林林下早春生长期植物盖度和密度均显著高于其他两种植物类群(P<0.05), 且早春展叶植物的盖度显著高于早春开花植物(P<0.05)(图 1)。
2.2 五种林型草本植物的多样性五种林型不同生活史对策植物的草本物种多样性差异显著(图 2)。早春开花植物的物种多样性指数在五种林型之间存在显著差异(P<0.05), 落叶松人工林的丰富度指数显著高于白桦次生林、阔叶红松择伐林和阔叶红松林(P<0.05), 红松人工林的丰富度指数显著高于阔叶红松择伐林和阔叶红松林(P<0.05);红松人工林和落叶松人工林的Shannon-Wiener指数均显著高于阔叶红松择伐林和阔叶红松林(P<0.05)。对于早春展叶植物, 白桦次生林的丰富度指数以及Shannon-Wiener指数均显著高于红松人工林、落叶松人工林以及阔叶红松择伐林(P<0.05), 且其丰富度指数显著高于阔叶红松林(P<0.05), 阔叶红松择伐林的丰富度指数显著低于红松人工林和阔叶红松林(P<0.05), 其Shannon-Wiener指数显著低于红松人工林(P<0.05)。

|
| 图 2 五种林型不同生活史对策的草本植物多样性差异(平均值±标准误差) Fig. 2 Differences of herbaceous plants diversity under different life-history strategies in five forest types (mean±SE) |
白桦次生林和红松人工林早春展叶植物的丰富度指数以及Shannon-Wiener指数均显著高于早春生长期植物以及早春开花植物(P<0.05), 落叶人工林的早春展叶植物的丰富度指数和Shannon-Wiener指数显著高于早春生长期植物(P<0.05), 阔叶红松择伐林早春生长期植物的丰富度指数显著高于早春开花植物(P<0.05), 阔叶红松林早春展叶植物的两种多样性指数均显著高于早春开花植物(P<0.05)(图 2)。
2.3 不同生活史对策草本植物密度、盖度和多样性与环境因子的关系土壤pH对早春生长期植物的密度(50.2%)、盖度(52.8%)丰富度指数(38.9%)以及Shannon-Wiener指数(36.7%)解释率最大, 对于早春开花植物和早春展叶植物, 土壤TP、pH以及林下光照因子对其解释率较高(图 3)。

|
| 图 3 环境因子对不同生活史对策草本植物密度、盖度以及物种多样性的层次分割结果 Fig. 3 Hierarchical partitioning results of environmental factors on density, coverage and species diversity of herbaceous plants under different life-history strategies D: 密度Density; C: 盖度Coverage; S: 物种丰富度指数Species richness index; H′: Shannon-Wiener指数Shannon-Wiener index; SOC: 土壤有机碳Soil organic carbon; TN: 土壤全氮Soil total nitrogen; TP: 土壤全磷Soil total phosphorus; pH: 土壤pH Soil pH; TDir: 林下直射光Transmitted direct light; TDif: 林下散射光Transmitted diffused light; TTot: 林下总光照Transmitted total light |
早春生长期植物的密度、盖度、丰富度指数以及Shannon-Wiener指数均与土壤pH呈显著正相关关系(P<0.05), 其丰富度指数和Shannon-Wiener指数与土壤有机碳含量呈显著的负相关关系(P<0.05), 与土壤全氮含量呈显著的单峰型关系(P<0.05)(图 4)。

|
| 图 4 早春生长期植物的密度、盖度以及物种多样性与环境因子的关系 Fig. 4 Relationships of density, coverage and species diversity of early-spring growing herbs with environmental factors |
早春开花植物密度与林下直射光、林下散射光以及林下总光照呈显著的单峰型关系(P<0.05), 其盖度与土壤pH呈显著的正相关关系(P<0.05), 与林下散射光和林下总光照呈显著的负相关关系(P<0.05)(图 5)。

|
| 图 5 早春开花植物的密度和盖度与环境因子的关系 Fig. 5 Relationships of density and coverage of early-spring flowering herbs with environmental factors |
对于早春展叶植物, 其密度和盖度与土壤全磷含量呈显著的单峰型关系(P<0.05), 盖度与林下散射光呈显著的正相关关系(P<0.05), 其丰富度指数和Shannon-Wiener指数与土壤pH呈极显著的负相关关系(P<0.01), 丰富度指数和Shannon-Wiener指数与林下直射光和林下总光照呈显著的“U”型关系、与林下散射光呈显著的正相关关系(P<0.05)(图 6)。

|
| 图 6 早春展叶植物的密度、盖度以及物种多样性与环境因子的关系 Fig. 6 Relationships of density, coverage and species diversity of early-spring foliating herbs with environmental factors |
本研究区域内五种林型的早春草本层植物群落组成的相似度较高, 林内不同生活史对策物种种类组成差异较小, 可能是由于研究区域内各林分的所处环境较为相似, 且草本植物组成的差异主要受立地条件影响, 与林分组成无关[25]。但林下优势物种存在差异, 是由于在人为采伐导致林型改变的条件下, 生境间的差异引起了优势种组成及分布的改变[23], 这可能是落叶松人工林下以林金腰为优势种, 而其他四种林型以银莲花为优势草本植物的原因之一。五种林型不同生活史对策草本植物的密度、盖度以及多样性指数差异显著, 这可能是由不同林分微环境的差异所导致。植物的生活史对策是维持种生长和繁殖的资源最佳分配方式[26], 同一林型间不同生活史对策植物的生长与多样性呈现显著差异, 这可能是由于不同生活史对策植物对资源分配策略的不同[27]。
3.2 早春草本层植物数量特征及多样性与土壤因子的关系人为采伐导致的环境异质性为物种共存和多样化提供了更多的生态位空间[28]。土壤的空间异质性强烈依赖于土壤结构和空间变异性, 对植物多样性的影响较为复杂[29]。本研究区域阔叶红松林由于人为采伐更新改变了林内树种组成, 而有研究表明森林管理导致的林分植被类型的差异会对土壤有机质含量以及C/N造成影响[30], 也可能通过改变林下的细根生长以及凋落物量影响森林土壤pH[31], 进而改变不同林型下草本植物的数量特征及其多样性。本研究结果显示, 早春生长期植物的丰富度指数及Shannon-Wiener指数与土壤全氮含量呈现出显著的单峰型关系, 这与王爱霞等的研究结果一致[10], 可能是因为N是植物体内重要细胞结构的组成元素, 是植物的各项生命活动的基础, 土壤全氮含量的适量增加对于草本植物的生长及其多样性起到促进作用, 但其含量过高则会导致土壤酸化和碱基阳离子的消耗, 造成养分失衡, 从而对植物的生长造成影响[32], 进而降低植物的多样性。早春生长期植物多样性指数与土壤有机碳含量呈显著负相关关系、早春展叶植物的密度和盖度与土壤全磷含量呈显著的单峰型关系, 均说明土壤养分是影响植物生长以及分布格局的重要环境因子[33—34]。土壤pH对五种林型林下草本植物多样性产生显著影响, 研究表明, pH与草本植物的丰富度指数和Shannon-Wiener指数均呈现显著的正相关关系[35], 与本研究pH对早春生长期植物多样性的研究结果一致。由于小兴安岭地区土壤代换性能的特点, 暗棕壤轻度不饱和, 发生酸性反应, 土壤pH通常在4.8—6.0之间[36], 本研究区域土壤pH的平均值为4.43, 酸性土壤意味着H+和Al3+含量较高, 且结果发现, 土壤pH与早春生长期植物的盖度、密度以及多样性指数呈显著的正相关关系, 与早春展叶植物的丰富度指数呈显著的负相关关系, 这可能与草本植物的生活史对策特性有关, 早春生长期植物对pH较低的土壤环境耐受性较低, 更适于偏中性的土壤, 酸性土壤不利于其生长;而早春展叶植物其生长条件则更偏向于酸性土壤, 与黄庆阳等的研究结果一致[34]。
3.3 早春草本层植物数量特征及多样性与林下光环境的关系冠层及其结构的差异驱动了光异质性的变化[37], 并决定林分和树木吸收的光量[38], 进而影响林下的光照条件。草本植物的生长与林内光照条件密切相关, 光的可利用性和异质性对林下草本植物组成及丰富度指数具有重要影响[39]。在早春的非生长季, 原始林和次生林的林下光照均显著高于夏季的生长季[40], 早春生长期植物在冠层展叶前生长, 林内充足的光照可以满足其完成营养生长阶段, 林分郁闭后则进入生殖生长阶段[7], 这可能是早春生长期植物的盖度、密度及多样性与林内光照无显著相关的原因;早春开花植物在早春时期和林分郁闭后都在进行生长过程, 林下的光照对其生长发育影响过程较为复杂, 其密度与林下直射光、林下散射光和总光照均呈显著的单峰型关系, 且其盖度与林下散射光和林下总光照呈显著的负相关关系, 可能是因为早春开花植物的生活史对策更偏向于低光照, 对强光照的耐受性较弱, 在遮荫的环境下才能更好的生长;而早春展叶植物的盖度与林下散射光呈显著正相关关系, 其物种丰富度和Shannon-Wiener指数与林下直射光和林下总光照均呈显著的“U”型关系, 表明早春展叶植物在高光照条件下具有更高的多样性, 这与前人的研究结果一致[41—42], 且在低光照条件下正常生长, 作为耐荫性植物能够适应林分对季节变化的响应。
4 结论本研究探讨了五种林型不同生活史对策的早春草本层植物组成、物种多样性及环境解释, 结果显示, 阔叶红松林的早春生长期植物的密度和盖度均显著高于红松人工林和落叶松人工林, 落叶松人工林的早春开花植物的密度、丰富度指数以及Shannon-Wiener指数均显著高于阔叶红松择伐林和阔叶红松林, 白桦次生林的丰富度指数以及Shannon-Wiener指数均显著高于除阔叶红松林外的其他三种林型。这与植物的生长特性和林下环境异质性密切相关, 主要受土壤pH和林下光照条件的影响。干扰后的森林在不断演替过程中, 草本植物为满足其生长发育需求而进化出不同的资源获取与分配策略以适应森林环境的变化, 使得森林在各个演替阶段均能达到动态的平衡。
附录| 生活史对策Life history strategy | 物种名Name |
| 早春生长期植物 Early-spring growing herbs | 侧金盏花Adonis amurensis |
| 早春生长期植物 Early-spring growing herbs | 顶冰花Gagea nakaiana |
| 早春生长期植物 Early-spring growing herbs | 荷青花Hylomecon japonica |
| 早春生长期植物 Early-spring growing herbs | 拟扁果草Enemion raddeanum |
| 早春生长期植物 Early-spring growing herbs | 菟葵Eranthis stellata |
| 早春生长期植物 Early-spring growing herbs | 五福花Adoxa moschatellina |
| 早春生长期植物 Early-spring growing herbs | 延胡索Corydalis yanhusuo |
| 早春生长期植物 Early-spring growing herbs | 银莲花Anemone cathayensis |
| 早春开花植物Early-spring flowering herbs | 白花碎米荠Cardamine leucantha |
| 早春开花植物Early-spring flowering herbs | 野芝麻Lamium barbatum |
| 早春开花植物Early-spring flowering herbs | 白屈菜Chelidonium majus |
| 早春开花植物Early-spring flowering herbs | 驴蹄草Caltha palustris |
| 早春开花植物Early-spring flowering herbs | 朝鲜堇菜Viola albida |
| 早春开花植物Early-spring flowering herbs | 林金腰Chrysosplenium lectus-cochleae |
| 早春开花植物Early-spring flowering herbs | 蔓孩儿参Pseudostellaria davidii |
| 早春开花植物Early-spring flowering herbs | 种阜草Moehringia lateriflora |
| 早春开花植物Early-spring flowering herbs | 附地菜Trigonotis peduncularis |
| 早春开花植物Early-spring flowering herbs | 酢浆草Oxalis corniculata |
| 早春开花植物Early-spring flowering herbs | 北重楼Paris verticillata |
| 早春展叶植物Early-spring foliating herbs | 野豌豆Vicia sepium |
| 早春展叶植物Early-spring foliating herbs | 粗茎鳞毛蕨Dryopteris crassirhizoma |
| 早春展叶植物Early-spring foliating herbs | 大叶柴胡Bupleurum longiradiatum |
| 早春展叶植物Early-spring foliating herbs | 耳叶蟹甲草Parasenecio auriculatus |
| 早春展叶植物Early-spring foliating herbs | 和尚菜Adenocaulon himalaicum |
| 早春展叶植物Early-spring foliating herbs | 黑水当归Angelica amurensis |
| 早春展叶植物Early-spring foliating herbs | 东北蹄盖蕨Athyrium brevifrons |
| 早春展叶植物Early-spring foliating herbs | 花荵Polemonium caeruleum |
| 早春展叶植物Early-spring foliating herbs | 灰脉薹草Carex appendiculata |
| 早春展叶植物Early-spring foliating herbs | 荚果蕨Matteuccia struthiopteris |
| 早春展叶植物Early-spring foliating herbs | 假冷蕨Athyrium spinulosum |
| 早春展叶植物Early-spring foliating herbs | 尖萼耧斗菜Aquilegia oxysepala |
| 早春展叶植物Early-spring foliating herbs | 尖嘴薹草Carex leiorhyncha |
| 早春展叶植物Early-spring foliating herbs | 华北剪股颖Agrostis clavata |
| 早春展叶植物Early-spring foliating herbs | 宽叶薹草Carex siderosticta |
| 早春展叶植物Early-spring foliating herbs | 宽叶荨麻Urtica laetevirens |
| 早春展叶植物Early-spring foliating herbs | 藜芦Veratrum nigrum |
| 早春展叶植物Early-spring foliating herbs | 铃兰Convallaria majalis |
| 早春展叶植物Early-spring foliating herbs | 龙常草Diarrhena mandshurica |
| 早春展叶植物Early-spring foliating herbs | 龙江风毛菊Saussurea amurensis |
| 早春展叶植物Early-spring foliating herbs | 毛缘薹草Carex pilosa |
| 早春展叶植物Early-spring foliating herbs | 木贼Equisetum hyemale |
| 早春展叶植物Early-spring foliating herbs | 披碱草Elymus dahuricus |
| 早春展叶植物Early-spring foliating herbs | 七瓣莲Trientalis europaea |
| 早春展叶植物Early-spring foliating herbs | 槭叶蚊子草Filipendula glaberrima |
| 早春展叶植物Early-spring foliating herbs | 贝加尔唐松草Thalictrum baicalense |
| 早春展叶植物Early-spring foliating herbs | 山韭Allium senescens |
| 早春展叶植物Early-spring foliating herbs | 升麻Cimicifuga foetida |
| 早春展叶植物Early-spring foliating herbs | 水杨梅Geum chiloense |
| 早春展叶植物Early-spring foliating herbs | 唢呐草Mitella nuda |
| 早春展叶植物Early-spring foliating herbs | 蚊子草Filipendula palmata |
| 早春展叶植物Early-spring foliating herbs | 乌苏里薹草Carex ussuriensis |
| 早春展叶植物Early-spring foliating herbs | 乌头Aconitum carmichaelii |
| 早春展叶植物Early-spring foliating herbs | 舞鹤草Maianthemum bifolium |
| 早春展叶植物Early-spring foliating herbs | 狭叶荨麻Urtica angustifolia |
| 早春展叶植物Early-spring foliating herbs | 大全叶山芹Ostericum maximowiczii var. australe |
| 早春展叶植物Early-spring foliating herbs | 东北羊角芹Aegopodium alpestre |
| 早春展叶植物Early-spring foliating herbs | 大叶章Deyeuxia purpurea |
| 早春展叶植物Early-spring foliating herbs | 烟管蓟Cirsium pendulum |
| 早春展叶植物Early-spring foliating herbs | 羊须草Carex callitrichos |
| 早春展叶植物Early-spring foliating herbs | 猪殃殃Galium spurium |
| [1] |
Isbell F, Gonzalez A, Loreau M, Cowles J, Díaz S, Hector A, Mace G M, Wardle D A, O'Connor M I, Duffy J E, Turnbull L A, Thompson P L, Larigauderie A. Linking the influence and dependence of people on biodiversity across scales. Nature, 2017, 546(7656): 65-72. DOI:10.1038/nature22899 |
| [2] |
Paul C, Hanley N, Meyer S T, Fürst C, Weisser W W, Knoke T. On the functional relationship between biodiversity and economic value. Science Advances, 2020, 6(5): eaax7712. DOI:10.1126/sciadv.aax7712 |
| [3] |
Chiatante G, Pellitteri-Rosa D, Torretta E, Nonnis Marzano F, Meriggi A. Indicators of biodiversity in an intensively cultivated and heavily human modified landscape. Ecological Indicators, 2021, 130: 108060. DOI:10.1016/j.ecolind.2021.108060 |
| [4] |
Sala O E, Stuart Chapin F, Armesto J J, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke L F, Jackson R B, Kinzig A, Leemans R, Lodge D M, Mooney H A, Oesterheld M, Poff N L, Sykes M T, Walker B H, Walker M, Wall D H. Global biodiversity scenarios for the year 2100. Science, 2000, 287(5459): 1770-1774. DOI:10.1126/science.287.5459.1770 |
| [5] |
Yu M, Sun O J. Effects of forest patch type and site on herb-layer vegetation in a temperate forest ecosystem. Forest Ecology and Management, 2013, 300: 14-20. DOI:10.1016/j.foreco.2012.12.039 |
| [6] |
Muller R N, Bormann F H. Role of Erythronium americanum Ker. in energy flow and nutrient dynamics of a northern hardwood forest ecosystem. Science, 1976, 193(4258): 1126-1128. |
| [7] |
吴刚, 尹若波, 周永斌, 郝占庆, 梁秀英. 长白山红松阔叶林林隙动态变化对早春草本植物的影响. 生态学报, 1999, 19(5): 659-663. DOI:10.3321/j.issn:1000-0933.1999.05.012 |
| [8] |
夏富才, 张春雨, 赵秀海, 潘春芳. 早春草本植物群落结构及其聚类分析. 东北师大学报: 自然科学版, 2008, 40(4): 109-114. DOI:10.3321/j.issn:1000-1832.2008.04.025 |
| [9] |
高景, 王金牛, 徐波, 谢雨, 贺俊东, 吴彦. 不同雪被厚度下典型高山草地早春植物叶片性状、株高及生物量分配的研究. 植物生态学报, 2016, 40(8): 775-787. |
| [10] |
王爱霞, 马婧婧, 龚会蝶, 范国安, 王茂, 赵红梅, 程军回. 北疆一年生早春短命植物物种丰富度分布格局及其影响因素. 生物多样性, 2021, 29(6): 735-745. |
| [11] |
Onaindia M, Dominguez I, Albizu I, Garbisu C, Amezaga I. Vegetation diversity and vertical structure as indicators of forest disturbance. Forest Ecology and Management, 2004, 195(3): 341-354. DOI:10.1016/j.foreco.2004.02.059 |
| [12] |
杨勇, 刘爱军, 李兰花, 王保林, 王明玖. 不同干扰方式对内蒙古典型草原植物种组成和功能群特征的影响. 应用生态学报, 2016, 27(3): 794-802. DOI:10.13287/j.1001-9332.201603.007 |
| [13] |
Vilén T, Gunia K, Verkerk P J, Seidl R, Schelhaas M J, Lindner M, Bellassen V. Reconstructed forest age structure in Europe 1950-2010. Forest Ecology and Management, 2012, 286: 203-218. |
| [14] |
Jandl R, Lindner M, Vesterdal L, Bauwens B, Baritz R, Hagedorn F, Johnson D W, Minkkinen K, Byrne K A. How strongly can forest management influence soil carbon sequestration?. Geoderma, 2007, 137(3/4): 253-268. |
| [15] |
Paillet Y, Bergès L, Hjältén J, Ódor P, Avon C, Bernhardt-römermann M, Bijlsma R J, de bruyn L, Fuhr M, Grandin U, Kanka R, Lundin L, Luque S, Magura T, Matesanz S, Mészáros I, Sebastià M T, Schmidt W, Standovár T, Tóthmérész B, Uotila A, Valladares F, Vellak K, Virtanen R. Biodiversity differences between managed and unmanaged forests: meta-analysis of species richness in Europe. Conservation Biology, 2010, 24(1): 101-112. DOI:10.1111/j.1523-1739.2009.01399.x |
| [16] |
Augusto L, Dupouey J L, Ranger J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Annals of Forest Science, 2003, 60(8): 823-831. |
| [17] |
余敏, 周志勇, 康峰峰, 欧阳帅, 米湘成, 孙建新. 山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释. 植物生态学报, 2013, 37(5): 373-383. |
| [18] |
Lai M Y, He S C, Yu S, Jin G Z. Effects of experimental N addition on plant diversity in an old-growth temperate forest. Ecology and Evolution, 2018, 8(11): 5900-5911. |
| [19] |
区余端, 苏志尧. 粤北山地常绿阔叶林自然干扰后冠层结构与林下光照动态. 生态学报, 2012, 32(18): 5637-5645. |
| [20] |
Bartels S F, Chen H Y H. Interactions between overstorey and understorey vegetation along an overstorey compositional gradient. Journal of Vegetation Science, 2013, 24(3): 543-552. |
| [21] |
史宝库, 金光泽, 汪兆洋. 小兴安岭5种林型土壤呼吸时空变异. 生态学报, 2012, 32(17): 5416-5428. |
| [22] |
Frazer G W, Canham C D, Lertzman K P. Gap Light Analyzer (GLA), Version 2.0: Imaging software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs, users manual and program documentation. Millbrook: Simon Fraser University, Burnaby, British Columbia and the Institute of Ecosystem Studies, 1999.
|
| [23] |
张涵丹, 康希睿, 邵文豪, 杨旭, 张建锋, 刘学全, 陈光才. 不同类型杉木人工林林下草本植物多样性特征. 生态学报, 2021, 41(6): 2118-2128. |
| [24] |
Nally R M, Walsh C J. Hierarchical partitioning public-domain software. Biodiversity and Conservation, 2004, 13(3): 659-660. |
| [25] |
Ewald J. The influence of coniferous canopies on understorey vegetation and soils in mountain forests of the northern Calcareous Alps. Applied Vegetation Science, 2000, 3(1): 123-134. |
| [26] |
班勇. 植物生活史对策的进化. 生态学杂志, 1995, 14(3): 33-39. |
| [27] |
Hartemink N, Jongejans E, de Kroon H. Flexible life history responses to flower and rosette bud removal in three perennial herbs. Oikos, 2004, 105(1): 159-167. |
| [28] |
Stein A, Gerstner K, Kreft H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecology Letters, 2014, 17(7): 866-880. |
| [29] |
Wang L, Liu C, Alves D G, Frank D A, Wang D L. Plant diversity is associated with the amount and spatial structure of soil heterogeneity in meadow steppe of China. Landscape Ecology, 2015, 30(9): 1713-1721. |
| [30] |
Oyonarte C, Aranda V, Durante P. Soil surface properties in Mediterranean Mountain ecosystems: effects of environmental factors and implications of management. Forest Ecology and Management, 2008, 254(2): 156-165. |
| [31] |
Russell A, Kivlin S, Hawkes C. Tropical tree species effects on soil pH and biotic factors and the consequences for macroaggregate dynamics. Forests, 2018, 9(4): 184. |
| [32] |
Magill A H, Aber J D, Currie W S, Nadelhoffer K J, Martin M E, McDowell W H, Melillo J M, Steudler P. Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA. Forest Ecology and Management, 2004, 196(1): 7-28. |
| [33] |
丛静, 尹华群, 卢慧, 宿秀江, 杨敬元, 李迪强, 张于光. 神农架保护区典型植被的物种多样性和环境解释. 林业科学, 2013, 49(5): 30-35. |
| [34] |
黄庆阳, 曹宏杰, 谢立红, 罗春雨, 杨帆, 王立民, 倪红伟. 五大连池火山熔岩台地草本层物种多样性及环境解释. 生物多样性, 2020, 28(6): 658-667. |
| [35] |
Rodríguez-Loinaz G, Onaindia M, Amezaga I, Mijangos I, Garbisu C. Relationship between vegetation diversity and soil functional diversity in native mixed-oak forests. Soil Biology and Biochemistry, 2008, 40(1): 49-60. |
| [36] |
中国林业科学研究院林业研究所. 中国森林土壤. 北京: 科学出版社, 1986.
|
| [37] |
Matsuo T, Martínez-Ramos M, Bongers F, van der Sande M T, Poorter L. Forest structure drives changes in light heterogeneity during tropical secondary forest succession. Journal of Ecology, 2021, 109(8): 2871-2884. |
| [38] |
Binkley D, Campoe O C, Gspaltl M, Forrester D I. Light absorption and use efficiency in forests: why patterns differ for trees and stands. Forest Ecology and Management, 2013, 288: 5-13. |
| [39] |
Kumar P, Chen H Y H, Thomas S C, Shahi C. Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forest of Canada. Journal of Ecology, 2018, 106(3): 1266-1276. |
| [40] |
Zhou G, Liu Q J, Xu Z Z, Du W X, Yu J, Meng S W, Zhou H, Qin L H, Shah S. How can the shade intolerant Korean pine survive under dense deciduous canopy?. Forest Ecology and Management, 2020, 457: 117735. |
| [41] |
Márialigeti S, Tinya F, Bidló A, Ódor P. Environmental drivers of the composition and diversity of the herb layer in mixed temperate forests in Hungary. Plant Ecology, 2016, 217(5): 549-563. |
| [42] |
Dormann C F, Bagnara M, Boch S, Hinderling J, Janeiro-Otero A, Schäfer D, Schall P, Hartig F. Plant species richness increases with light availability, but not variability, in temperate forests understorey. BMC Ecology, 2020, 20(1): 43. |