 2023, Vol. 43
2023, Vol. 43文章信息
- 董彦君, 刘杰, 陈颖, 李宗善, 高光耀, 王聪, 王晓春
- DONG Yanjun, LIU Jie, CHEN Ying, LI Zongshan, GAO Guangyao, WANG Cong, WANG Xiaochun
- 锡林浩特多年生草本根部导管性状与植株生态策略的相关特征
- Vessel traits of main roots are linked to ecological strategies of the perennial herbs in Xilinhot of China
- 生态学报. 2023, 43(3): 1126-1134
- Acta Ecologica Sinica. 2023, 43(3): 1126-1134
- http://dx.doi.org/10.5846/stxb202108262399
-
文章历史
- 收稿日期: 2021-08-26





2. 中国科学院大学, 北京 100049;
3. 陕西黄土高原地球关键带国家野外科学观测研究站, 西安 710061;
4. 东北林业大学生态研究中心, 哈尔滨 150040
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. National Observation and Research Station of Earth Critical Zone on the Loess Plateau in Shaanxi, Xi'an 710061, China;
4. Center for Ecological Research, Northeast Forestry University, Harbin 150040, China
在中国干旱半干旱区, 水分是对植物生长发育最具限制性的环境因子, 水分胁迫会导致植物气孔关闭, 光合作用下降, 从而降低植物的生产力, 影响植物的生长[1—2]。干旱胁迫也会引起植物木质部栓塞, 造成水力运输故障, 最终会导致器官甚至整株植物的死亡[3]。根据哈根-泊肃叶(Hagen-Poiseuille)方程[4], 水力传导率与导管直径的四次方成正比, 因此导管的面积可直接影响植物的水分传导效率。有研究表明, 导管越宽植物生长越快, 因为更宽的导管允许更高的蒸腾从而能促进光合作用[5—6]。但在胁迫条件下, 木质部中的负压增强, 较宽的导管更容易产生空穴化, 从而导致水分运输的安全性降低[4, 7]。这种木质部安全与效率之间的权衡, 对于生长在干旱区的植物尤为重要[8]。与导管的大小相比, 导管数量对水力传导率的影响较小, 但导管密度可直接影响木质部的结构组成, 从而影响其强度及其在负压下抵抗导管内爆的能力[9]。安全的水力系统往往由更小更多的导管组成[10], 因为小导管可以降低低温或干旱胁迫引起栓塞的风险[11], 但小导管增加了水分运输的阻力, 因此水力效率较低[12], 从而可能会影响光合作用, 导致生长缓慢[13]。
通常认为植株的高度可能与木质部的水力结构之间存在联系[14], 水分运输是决定植物生长高度的主要限制性因素[15]。随植物高度增加, 水分运输阻力也会增大, 植物必须通过调节一些水力学特性, 在输水有效性和安全性之间做出权衡[15—16]。有研究发现木本植物的株高与导管面积正相关, 与导管密度负相关, 因为导管面积越大, 其水分输送效率越高, 而较小的导管密度可为起机械支撑作用的纤维提供更多的空间[5, 17]。同样地, 另一项草本植物的研究中发现高大植物根的平均导管横截面积较大, 因为导管面积对成熟根系的水力传导有重大影响, 但没有发现导管密度与株高的关系[13]。然而相关的研究大多集中在木本植物[5, 18—28], 对草本植物的研究还没有引起重视, 而这些结论需要在更广泛的物种上进行验证, 因此, 本研究选择了草本植物为研究对象。
根系解剖特征与整株植物的功能密切相关。与水力传输有关的解剖结构(如导管面积、导管数量、木质部比例、木质部面积等)[5, 19]、与机械强度有关的解剖结构(如中柱面积、中柱比例等)[13]都会影响整株植物的功能。此外, 根直径或皮层宽度也可通过影响水力传输效率进而影响植株的生长速率和株高[8, 29]。虽然解剖结构和植物功能之间的关系已被广泛认可, 但在根系水平上, 很少有对两者关系的证实[8, 30]。因此, 本研究在根系水平上, 分析了与水力传输有关的解剖性状与植物功能之间的关系。
本研究的目的是探究物种整株功能的3个关键性状(年龄、生长速率和株高)是否与导管解剖性状有关, 以了解植物对环境的适应机制及生存策略。为此本研究在中国干旱半干旱区的锡林浩特草原进行了采样, 重点研究了采集到的15个科的42种草本植物。采用石蜡切片法获取到根部导管解剖结构参量(导管数量、导管分数、导管面积、水力传导效率和水力直径)和生长参量(年龄和生长速率), 运用逐步回归法和Pearson相关分析了导管解剖性状与生长特征之间的关系, 以确定物种整株功能的种间变异背后的结构特征。
1 材料与方法 1.1 样品采集与处理本研究的采样区位于中国内蒙古的锡林浩特草原(43°02′—44°52′N, 115°18′—117°06′E)(图 1), 该地区处于内蒙古高原中部, 地势南高北低, 平均海拔988.5 m。属中温带半干旱大陆性气候, 降水量309 mm, 年均气温为2.0℃。从国家气象信息中心(http://data.cma.cn/)获取了1981—2019年的气候数据。本研究于2020年7月—8月在锡林浩特草原共设置了11个采样点(表 1), 在每个样点采集该区域内广泛分布的多年生草本物种, 每个物种挖取5株左右健康的植株, 在其主根近地面3 cm左右的位置截取一段, 迅速放入FAA固定液(70%乙醇: 35%—40%甲醛: 乙酸=90: 5: 5)中固定保存。共采集到42种多年生草本的主根, 295个样品。

|
| 图 1 锡林浩特草原草本物种采样点空间分布图 Fig. 1 Locations of sampling sites of perennial forbs species across the Xilinhot Grassland, China |
| 采样点 Site |
经度/(°E) Longitude |
纬度/(°N) Latitude |
海拔 Altitude /m |
降雨量 Precipitation /mm |
温度 Temperature/℃ |
物种数 Species number |
| S1 | 116.66 | 43.55 | 1260 | 321 | 1.51 | 5 |
| S2 | 116.67 | 43.55 | 1271 | 320 | 1.52 | 14 |
| S3 | 116.53 | 44.27 | 1140 | 282 | 1.39 | 8 |
| S4 | 114.79 | 43.94 | 1117 | 229 | 1.78 | 8 |
| S5 | 115.49 | 43.91 | 1131 | 227 | 1.80 | 3 |
| S6 | 116.1 | 44.37 | 938 | 253 | 2.58 | 5 |
| S7 | 116.09 | 44.37 | 935 | 253 | 2.59 | 2 |
| S8 | 117.6 | 44.58 | 1050 | 297 | 1.71 | 6 |
| S9 | 118.15 | 44.76 | 1125 | 342 | 0.94 | 3 |
| S10 | 118.92 | 45.04 | 1015 | 358 | 1.03 | 2 |
| S11 | 119.35 | 45.31 | 972 | 372 | 0.79 | 3 |
完成采样后, 在实验室内根据传统的石蜡切片法[31—32], 将采集到的草本植物主根样品制成石蜡切片, 切片厚度为8—12 μm, 然后采用番红—固绿二重染色法染色, 用中性树胶封片, 制成永久装片。做5个根段的重复, 每个重复切5个样片。最后在Olympus DP73 (Olympus, Tokyo, Japan)光学显微镜下用同一倍数(×40)观察并拍照。
1.2 数据处理与统计方法使用ImageJ图像处理软件, 在根的横切片上选取一个固定的木质部导管区域(850 μm× 850 μm), 自动读取被测量区域内的导管数量(Vessel number, NV)、导管分数(Vessel fraction, VF)(测量范围内导管总面积占测量面积的比例)、平均导管面积(Mean vessel area, MVA)等指标。多年生草本植株的平均高度Hmean= (Hmax+Hmin)/2, 其中Hmax和Hmin分别为植株的最大高度和最小高度, 通过查中国植物志(http://www.iplant.cn/frps)得到。通过分辨根木质部中的早材和晚材来确定年轮[33]。草本植物的年龄是由物种根的横切结构中的年轮数决定的[34]。植物平均每年的生长量即生长速率(Growth rate, GR)的计算公式为GR=W/A, 其中A为植株生长的年龄, W为A年生长的宽度(图 2)。由于导管是近似椭圆, 因此导管直径D=[3(ab)3/(2a2+2b2)]1/4, 其中a和b分别为导管长轴和短轴的值[35]。理论水力传导率(The theoretical hydraulic conductivity, Kp)的计算, 根据哈根-泊肃叶方程(Hagen-Poiseuille)[4] 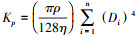
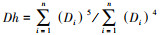

|
| 图 2 达乌里芯芭(Cymbaria dahurica)的根部横截面 Fig. 2 Root cross-sections of Cymbaria dahurica 图中黄线标出了年轮界限, W表示草本10年生长的宽度。比例尺为500 μm |
本研究中数据记录与初步整理主要用Excel 2019;数据分析主要采用软件SPSS 26.0和R语言程序;图形绘制采用Origin 2018和ArcGIS 9.2完成。
2 结果与分析 2.1 锡林浩特草原草本物种导管结构参数及变化范围本研究在锡林浩特草原共采集到42种草本, 分属于15个科, 32个属, 其中豆科、菊科、藜科和蔷薇科的物种最多(表 2)。从锡林浩特草原草本物种生长特征参量和根部导管解剖结构特征频数分布图(图 3)中可知, 采集到的草本物种的年龄范围为1—13年, 平均年龄为5年, 年龄在2到5年的草本物种约占60%;冷蒿(Artemisia frigida)的生长速率为50.61 μm/a, 是生长速率最低的物种, 蓟(Cirsium japonicum)的生长速率为842.31 μm/a, 是生长速率最高的物种;平均高度最低的物种为百里香(Thymus mongolicus), 仅0.06 m;在固定测量面积内, 泥胡菜(Hemistepta lyrata)的导管数量最少(31个), 冷蒿的导管数量最多(480个);大部分物种的导管分数在2%—9%之间, 平均值为6.56%;平均导管面积最小的物种是翻白委陵菜(Potentilla chinensis), 数值为93.92 μm2, 平均导管面积最大的物种是田旋花(Convolvulus arvensis), 数值为982.88 μm2;73%的草本物种的平均水力传导率不超过0.5 kg m MPa-1 s-1, 叉枝蓼(Polygonum tortuosum)的水力直径最大, 为52.28 μm, 水力直径在15—24 μm之间的物种占65%。
| 物种名称Species | 科名Family | 属名Genera |
| 车前Plantago asiatica | 车前科Plantaginaceae | 车前属Plantago |
| 补血草Limonium sinense | 白花丹科Plumbaginaceae | 补血草属Limonium |
| 百里香Thymus mongolicus | 唇形科Labiatae | 百里香属Thymus |
| 糙苏Phlomis umbrosa | 唇形科Labiatae | 糙苏属Phlomis |
| 达乌里胡枝子Lespedeza davurica | 豆科Leguminosae | 胡枝子属Lespedeza |
| 乳白黄耆Astragalus galactites | 豆科Leguminosae | 黄耆属Astragalus |
| 黄耆Astragalus membranaceus | 豆科Leguminosae | 黄耆属Astragalus |
| 砂珍棘豆Oxytropis psamocharis | 豆科Leguminosae | 棘豆属Oxytropis |
| 棘豆Oxytropis Myriophylla | 豆科Leguminosae | 棘豆属Oxytropis |
| 狭叶锦鸡儿Caragana stenophylla | 豆科Leguminosae | 锦鸡儿属Caragana |
| 花苜蓿Medicago ruthenica | 豆科Leguminosae | 苜蓿属Medicago |
| 飞廉Carduus nutans | 菊科Compositae | 飞廉属Carduus |
| 阿尔泰狗娃花Heteropappus altaicus | 菊科Compositae | 狗娃花属Heteropappus |
| 黄蒿Artemisia annua | 菊科Compositae | 蒿属Artemisia |
| 冷蒿Artemisia frigida | 菊科Compositae | 蒿属Artemisia |
| 长叶火绒草Leontopodium longifolium | 菊科Compositae | 火绒草属Leontopodium |
| 蓟Cirsium japonicum | 菊科Compositae | 蓟属Cirsium |
| 麻花头Serratula centauroides | 菊科Compositae | 麻花头属Serratula |
| 泥胡菜Hemistepta lyrata | 菊科Compositae | 泥胡菜属Hemistepta |
| 天葵Semiaquilegia adoxoides | 菊科Compositae | 天葵属Semiaquilegia |
| 烟管头草Carpesium cernuum | 菊科Compositae | 天名精属Carpesium |
| 栉叶蒿Neopallasia pectinata | 菊科Compositae | 栉叶蒿属Neopallasia |
| 鹤虱Lappula myosotis | 紫草科Boraginaceae | 鹤虱属Lappula |
| 田旋花Convolvulus arvensis | 旋花科Convolvulaceae | 旋花属Convolvulus |
| 木地肤Kochia prostrata | 藜科Chenopodiaceae | 地肤属Kochia |
| 灰绿藜Chenopodium glaucum | 藜科Chenopodiaceae | 藜属Chenopodium |
| 藜Chenopodium album | 藜科Chenopodiaceae | 藜属Chenopodium |
| 尖头叶藜Chenopodium acuminatum | 藜科Chenopodiaceae | 藜属Chenopodium |
| 猪毛菜Salsola collina | 藜科Chenopodiaceae | 猪毛菜属Salsola |
| 萹蓄Polygonum aviculare | 蓼科Polygonaceae | 蓼属Polygonum |
| 叉枝蓼Polygonum tortuosum | 蓼科Polygonaceae | 蓼属Polygonum |
| 翻白草Potentilla discolor | 蔷薇科Rosaceae | 委陵菜属Potentilla |
| 菊叶委陵菜Potentilla tanacetifolia | 蔷薇科Rosaceae | 委陵菜属Potentilla |
| 二裂委陵菜Potentilla bifurca | 蔷薇科Rosaceae | 委陵菜属Potentilla |
| 翻白委陵菜Potentilla chinensis | 蔷薇科Rosaceae | 委陵菜属Potentilla |
| 星毛委陵菜Potentilla acaulis | 蔷薇科Rosaceae | 委陵菜属Potentilla |
| 狼毒Euphorbia fischeriana | 瑞香科Thymelaeaceae | 狼毒属Euphorbia |
| 防风Saposhnikovia divaricata | 伞形科Umbelliferae | 防风属Saposhnikovia |
| 荠菜Capsella bursa-pastoris | 十字花科Cruciferae | 荠属Capsella |
| 女娄菜Silene aprica | 石竹科Caryophyllaceae | 蝇子草属Silene |
| 达乌里芯芭Cymbaria dahurica | 玄参科Scrophulariaceae | 芯芭属Cymbaria |
| 北芸香Haplophyllum dauricum | 芸香科Rutaceae | 芸香属Haplophyllum |

|
| 图 3 锡林浩特草原草本物种生长特征参数和根部导管解剖结构特征频数分布图 Fig. 3 Frequency Diagram of growth traits and the major anatomical traits in secondary root xylem for herb species in Xilinhot grassland |
从图 4中可以看出, 各导管参量与年龄无相关关系;植株的生长速率与导管数量(R=-0.494, P < 0.01)和导管分数(R=-0.255, P < 0.05)显著负相关关系, 与平均导管面积(R=0.274, P < 0.05)、平均水力传导率(R=0.263, P < 0.05)和水力直径(R=0.245, P < 0.05)显著正相关;植株的平均高度与导管数量(R=-0.354, P < 0.01)显著负相关, 与平均导管面积(R=0.293, P < 0.05)、平均水力传导率(R=0.289, P < 0.05)和水力直径(R=0.278, P < 0.05)显著正相关。

|
| 图 4 生长特征与导管解剖结构参量之间的相关性 Fig. 4 Relationship between growth characteristics and hydraulic conductivity traits 进行相关分析前, 对原始数据进行了log10转换; 相关系数和显著性已标出:**P < 0.01. *P < 0.05 |
以年龄、GR和Hmean分别为被解释变量, NV、VF、MVA、MKp和Dh为解释变量, 利用R语言程序, 建立多元线性回归模型, 采用逐步回归法对模型进行修正, 最终得到GR和Hmean的多元回归方程(表 3)。从模型中可以看出, NV与GR成负相关, 是影响GR的主要导管参量, 而其余的导管性状(VF、MVA、MKp和Dh)在逐步回归中被剔除, NV解释了GR 25%的变异;NV与Hmean成负相关, 影响Hmean的主要导管解剖参量也是NV, 而其余的导管性状在逐步回归中被剔除, NV解释了Hmean 13%的变异。
| 模型 Models |
可决系数 Coefficient of determination |
调整后系数 Adjusted Coefficient |
概率值 Probability value |
赤池信息量准则 Akaike infomation criterion |
| GR=-0.489 NV+3.514 | 0.245*** | 0.234*** | < 0.001 | -212.74 |
| Hmean=-0.376 NV+ 0.404 | 0.126** | 0.113** | < 0.01 | -191.53 |
| GR为生长速率Growth rate;Hmean为平均株高Mean height;NV为导管数量; **P < 0.01; ***P < 0.001 | ||||
锡林浩特草原多年生草本植物根部的MVA、MKp和Dh与GR显著正相关(图 4), 这表明生长快的草本物种具有更大的导管, 即生长快的草本物种比生长缓慢的物种需要更高的水分运输能力。锡林浩特草原位于中国的干旱半干旱区, 在该地区干旱是限制植物生长速率的主要环境因子。大导管具有更高的导水效率, 有效的水分运输允许更高的气孔导度, 从而提高光合速率, 促进生长速率[37—38], 这一现象是水力性状与光合作用协调的结果[38]。Wahl和Ryser[13]对来自不同生境的19种多年生禾本科植物的根解剖特征进行了研究, 结果发现生长快速的物种比生长缓慢的物种具有更大的平均导管横截面积, 这与本研究的结果一致。同时, 在木本植物茎中的发现也证实了本研究结果[5, 18, 22, 39]。锡林浩特草原多年生草本植物根部固定范围内的NV与GR极显著负相关(图 4), 表明生长快的物种具有更少的导管, 导管面积对水力传导率的贡献比导管数量更大[4, 40], 因此生长快的物种仍具有较高的水力传导能力;而生长缓慢的草本物种则具有较多的小导管, 这可能反映了对导管功能的更好保护[13]。导管的大小和数量与水分传导的有效性和安全性有关, 根据哈根—泊肃叶定律[4], 水力传导率与导管直径的四次方成正比。宽导管的导水效率更高, 但是脆弱易倒塌, 且容易发生栓塞[41];而窄导管的导水效率虽低, 但在胁迫条件下抵抗空穴化的能力强, 导管不易栓塞, 从而避免水力系统崩溃, 保证导管正常的水力运输功能[42]。另外, 固定面积内的导管数量越多, 水力系统的安全性越高, 因为当部分导管发生空穴化而导致水分运输功能丧失时, 仍有大量保持正常功能的导管可以继续运输水分, 以维持植株的正常生长[43]。研究还表明, 导管发生栓塞后, 小导管比大导管恢复更快, 因此空穴化的时间更短[44]。
3.2 导管解剖性状与植株高度的关系本研究中锡林浩特草原物种的株高范围为0.06—2 m。多年生草本的导管性状与Hmean之间的关系, 同其与GR之间的关系基本一致, 即表征导水能力的性状(MVA、MKp和Dh)与Hmean正相关, 而NV与Hmean负相关, 这表明高大的植株具有更强的水分传导能力和更少的导管, 而矮小的植株具有较低的导水效率和更多的导管(图 4)。另一项对多年生禾本科草本植物的相关研究发现高大植物的平均导管横截面积比矮小物种的更大, 这与本研究结果一致;但该研究中植物的高度与导管数量无关, 这与本研究的发现不同[13]。此外, 在木本植物中, 也发现了与本研究一致的结果, 比如对51种加利福尼亚被子植物的研究发现, 高的物种比矮的物种有更大的导管, 但导管密度也更低[19]。同样的, 在对玻利维亚42种热带雨林树种的研究中也发现了相似的结果[20]。还有研究发现, 乔木的导管面积比灌木的大, 导管密度更低[11], 也证实了本研究结果。这些研究均表明, 当植株的高度增加, 导管的面积也随之增加, 从而抵消了因水分运输路径变长而增加的水力运输阻力[5, 45—46]。植物液流通量与水分输导距离成反比[47], 因此当水分运输距离增加, 植株要维持一个固定的液流通量, 就需要增大导管直径[48]。
高大的植株把张力传递给根部, 这可能会改变根系的机械强度[8], 因此高大植株根部较低的导管数量, 可以补偿其较大的导管直径, 从而维持一定的机械支撑[19]。矮小植株具有多而小的导管, 水力传导率较低, 但水力运输安全性较高, 这种木质部安全与效率的权衡保证了植株的水力功能[12], 这对于生长在干旱区的植物可能尤为重要。有研究表明, 除了根导管解剖结构外, 根直径[13]、根密度[8]、皮层宽度[29]、木质部面积[8, 13]等都可能与植株的高度有关。因此需要进一步的实验, 探究根其他解剖结构与植物生长特征之间的关系。
4 结论本研究以生长在中国锡林浩特草原的多年生草本为研究对象, 为草本植物的全株功能与根系导管解剖之间的密切关系提供了证据。结果发现, 植物的生长速率与表征水力能力的性状(平均导管面积、平均水力传导率、水力直径)正相关, 与导管数量负相关, 即生长快的物种具有大而少的导管, 而生长慢的物种具有小而多的导管, 这表明生长快的物种需要较高的水分运输能力, 而生长慢的物种, 具有较安全的水力运输系统;植物的株高与表征水力能力的性状正相关, 与导管数量负相关, 即高大的物种具有大而少的导管, 而矮小的物种具有小而多的导管, 这表明高大的植株通过增大导管直径减小水力运输阻力, 较少的导管补偿较大的导管以维持一定的机械强度, 而矮小植物有较低的导水效率, 维持较高的水力运输安全性, 这种效率与安全的权衡对于生长在干旱区的植物非常重要。虽然本研究证实了多年生草本植物根导管性状与整株功能密切相关, 但根的其他结构(如根直径、木质部面积等)和其他植物器官(茎、叶)也可能影响植株的生态功能, 因此需要进一步的研究。
| [1] |
Blackman C J, Brodribb T J, Jordan G J. Leaf hydraulics and drought stress: response, recovery and survivorship in four woody temperate plant species. Plant, Cell & Environment, 2009, 32(11): 1584-1595. |
| [2] |
Bréda N, Huc R, Granier A, Dreyer E. Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Annals of Forest Science, 2006, 63(6): 625-644. DOI:10.1051/forest:2006042 |
| [3] |
Scoffoni C, Albuquerque C, Brodersen C R, Townes S V, John G P, Cochard H, Buckley T N, McElrone A J, Sack L. Leaf vein xylem conduit diameter influences susceptibility to embolism and hydraulic decline. New Phytologist, 2017, 213(3): 1076-1092. DOI:10.1111/nph.14256 |
| [4] |
Tyree M T, Zimmermann M H. Xylem Structure and the Ascent of Sap. Berlin, Heidelberg: Springer Berlin Heidelberg, 2002.
|
| [5] |
Fan Z X, Zhang S B, Hao G Y, Ferry Slik J W, Cao K F. Hydraulic conductivity traits predict growth rates and adult stature of 40 Asian tropical tree species better than wood density. Journal of Ecology, 2012, 100(3): 732-741. DOI:10.1111/j.1365-2745.2011.01939.x |
| [6] |
Ishida A, Nakano T, Yazaki K, Matsuki S, Koike N, Lauenstein D L, Shimizu M, Yamashita N. Coordination between leaf and stem traits related to leaf carbon gain and hydraulics across 32 drought-tolerant angiosperms. Oecologia, 2008, 156(1): 193-202. DOI:10.1007/s00442-008-0965-6 |
| [7] |
Davis S D, Sperry J S, Hacke U G. The relationship between xylem conduit diameter and cavitation caused by freezing. American Journal of Botany, 1999, 86(10): 1367-1372. DOI:10.2307/2656919 |
| [8] |
Hummel I, Vile D, Violle C, Devaux J, Ricci B, Blanchard A, Garnier É, Roumet C. Relating root structure and anatomy to whole-plant functioning in 14 herbaceous Mediterranean species. New Phytologist, 2007, 173(2): 313-321. DOI:10.1111/j.1469-8137.2006.01912.x |
| [9] |
Jacobsen A L, Ewers F W, Pratt R B, Paddock W A, Davis S D. Do xylem fibers affect vessel cavitation resistance?. Plant Physiology, 2005, 139(1): 546-556. DOI:10.1104/pp.104.058404 |
| [10] |
Yáñez-Espinosa L, Terrazas T, López-Mata L. Effects of flooding on wood and bark anatomy of four species in a mangrove forest community. Trees, 2001, 15(2): 91-97. DOI:10.1007/s004680000081 |
| [11] |
Carlquist S, Hoekman D A. Ecological wood anatomy of the woody southern Californian flora. IAWA Journal, 1985, 6(4): 347. |
| [12] |
Hacke U G, Sperry J S, Wheeler J K, Castro L. Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiology, 2006, 26(6): 689-701. DOI:10.1093/treephys/26.6.689 |
| [13] |
Wahl S, Ryser P. Root tissue structure is linked to ecological strategies of grasses. New Phytologist, 2000, 148(3): 459-471. DOI:10.1046/j.1469-8137.2000.00775.x |
| [14] |
King D A, Davies S J, Tan S, Noor N S M. The role of wood density and stem support costs in the growth and mortality of tropical trees. Journal of Ecology, 2006, 94(3): 670-680. DOI:10.1111/j.1365-2745.2006.01112.x |
| [15] |
Ryan M G, Yoder B J. Hydraulic limits to tree height and tree growth. BioScience, 1997, 47(4): 235-242. DOI:10.2307/1313077 |
| [16] |
Sperry J S, Meinzer F C, McCULLOH K A. Safety and efficiency conflicts in hydraulic architecture: scaling from tissues to trees. Plant, Cell & Environment, 2008, 31(5): 632-645. |
| [17] |
Martínez-Cabrera H I, Schenk H J, Cevallos-Ferriz S R S, Jones C S. Integration of vessel traits, wood density, and height in angiosperm shrubs and trees. American Journal of Botany, 2011, 98(5): 915-922. DOI:10.3732/ajb.1000335 |
| [18] |
Zhang J L, Cao K F. Stem hydraulics mediates leaf water status, carbon gain, nutrient use efficiencies and plant growth rates across dipterocarp species. Functional Ecology, 2009, 23(4): 658-667. DOI:10.1111/j.1365-2435.2009.01552.x |
| [19] |
Preston K A, Cornwell W K, DeNoyer J L. Wood density and vessel traits as distinct correlates of ecological strategy in 51 California coast range angiosperms. New Phytologist, 2006, 170(4): 807-818. DOI:10.1111/j.1469-8137.2006.01712.x |
| [20] |
Poorter L, McDonald I, Alarcón A, Fichtler E, Licona J C, Peña-Claros M, Sterck F, Villegas Z, Sass-Klaassen U. The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species. New Phytologist, 2010, 185(2): 481-492. DOI:10.1111/j.1469-8137.2009.03092.x |
| [21] |
Pertiwi Y, Aiso H, Ishiguri F, Wedatama S, Marsoem S, Ohshima J, Iizuka K, Yokota S. Effect of radial growth rate on wood properties of neolamarckia cadamba. Journal of Tropical Forest Science, 2017, 29(1): 30-36. |
| [22] |
Yan Y M, Fan Z, Fu P L, Chen H, Lin L. Size dependent associations between tree diameter growth rates and functional traits in an Asian tropical seasonal rainforest. Functional Plant Biology, 2020. |
| [23] |
Cao X, Shen Q D, Liu L, Cheng J L. Relationships of growth, stable carbon isotope composition and anatomical properties of leaf and xylem in seven mulberry cultivars: a hint to drought tolerance. Plant Biology, 2019. |
| [24] |
Ishiguri F, Wahyudi I, Takashima Y, Ohshima J, Yokota S. Effects of radial growth rate on anatomical characteristics and wood properties in peronema canescens trees planted in south Kalimantan, Indonesia. Journal of Tropical Forest Science, 2021, 33(1): 22-29. DOI:10.26525/jtfs2021.33.1.22 |
| [25] |
Anfodillo T, Olson M E. Tree mortality: testing the link between drought, embolism vulnerability, and xylem conduit diameter remains a priori ty. Frontiers in Forests and Global Change, 2021, 4: 704670. DOI:10.3389/ffgc.2021.704670 |
| [26] |
Li M Y, Leng Q N, Hao G Y. Contrasting patterns of radial growth rate between Larix principis-rupprechtii and Pinus sylvestris var. mongolica along an elevational gradient are mediated by differences in xylem hydraulics. Forest Ecology and Management, 2021, 497: 119524. DOI:10.1016/j.foreco.2021.119524 |
| [27] |
Zhao H, Jiang Z M, Zhang Y J, Jiang B, Cai J. Hydraulic efficiency at the whole tree level stably correlated with productivity over years in 9 poplar hybrids clones. Forest Ecology and Management, 2021, 496: 119382. DOI:10.1016/j.foreco.2021.119382 |
| [28] |
Qi J H, Fan Z X, Fu P L, Zhang Y J, Sterck F. Differential determinants of growth rates in subtropical evergreen and deciduous juvenile trees: carbon gain, hydraulics and nutrient-use efficiencies. Tree Physiology, 2020, 41(1): 12-23. |
| [29] |
Rieger M, Litvin P. Root system hydraulic conductivity in species with contrasting root anatomy. Journal of Experimental Botany, 1999, 50(331): 201-209. DOI:10.1093/jxb/50.331.201 |
| [30] |
Eissenstat D. Root structure and function in an ecological context. New Phytologist, 2000, 148(3): 353-354. DOI:10.1046/j.1469-8137.2000.00781.x |
| [31] |
张宝华. 聚焦植物石蜡切片制作. 吉林省教育学院学报: 下旬, 2013, 29(4): 153-154. |
| [32] |
张军周, 勾晓华, 赵志千, 刘文火, 张芬, 曹宗英, 周非飞. 树轮生态学研究中微树芯石蜡切片制作的方法探讨. 植物生态学报, 2013, 37(10): 972-977. |
| [33] |
von Arx G, Dietz H. Growth rings in the roots of temperate forbs are robust annual markers. Plant Biology, 2006, 8(2): 224-233. DOI:10.1055/s-2005-873051 |
| [34] |
von Arx G, Archer S R, Hughes M K. Long-term functional plasticity in plant hydraulic architecture in response to supplemental moisture. Annals of Botany, 2012, 109(6): 1091-1100. DOI:10.1093/aob/mcs030 |
| [35] |
Lewis A M, Boose E R. Estimating volume flow rates through xylem conduits. American Journal of Botany, 1995, 82(9): 1112-1116. DOI:10.1002/j.1537-2197.1995.tb11581.x |
| [36] |
Sperry J S, Nichols K L, Sullivan J E M, Eastlack S E. Xylem embolism in ring-porous, diffuse-porous, and coniferous trees of northern Utah and interior Alaska. Ecology, 1994, 75(6): 1736-1752. DOI:10.2307/1939633 |
| [37] |
Brodribb T J, Holbrook N M, Gutiérrez M V. Hydraulic and photosynthetic co-ordination in seasonally dry tropical forest trees. Plant, Cell & Environment, 2002, 25(11): 1435-1444. |
| [38] |
Santiago L S, Goldstein G, Meinzer F C, Fisher J B, Machado K, Woodruff D, Jones T. Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees. Oecologia, 2004, 140(4): 543-550. DOI:10.1007/s00442-004-1624-1 |
| [39] |
Castro-Díez P, Puyravaud J P, Cornelissen J H C, Villar-Salvador P. Stem anatomy and relative growth rate in seedlings of a wide range of woody plant species and types. Oecologia, 1998, 116(1/2): 57-66. |
| [40] |
McCully M E, Canny M J. Pathways and processes of water and nutrient movement in roots. Plant and Soil, 1988, 111(2): 159-170. DOI:10.1007/BF02139932 |
| [41] |
Linton M J, Sperry J S, Williams D G. Limits to water transport in Juniperus osteosperma and Pinus edulis: implications for drought tolerance and regulation of transpiration. Functional Ecology, 1998, 12(6): 906-911. DOI:10.1046/j.1365-2435.1998.00275.x |
| [42] |
Zimmermann M H. Hydraulic architecture of some diffuse-porous trees. Canadian Journal of Botany, 1978, 56(18): 2286-2295. DOI:10.1139/b78-274 |
| [43] |
Zimmermann M H. Functional xylem anatomy of angiosperm treesNew Perspectives in Wood Anatomy, 1982.
|
| [44] |
McCULLY M E, Huang C X, Ling L E C. Daily embolism and refilling of xylem vessels in the roots of field-grown maize. New Phytologist, 1998, 138(2): 327-342. DOI:10.1046/j.1469-8137.1998.00101.x |
| [45] |
Koch G W, Sillett S C, Jennings G M, Davis S D. The limits to tree height. Nature, 2004, 428(6985): 851-854. DOI:10.1038/nature02417 |
| [46] |
Mencuccini M. The ecological significance of long-distance water transport: short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms. Plant, Cell & Environment, 2003, 26(1): 163-182. |
| [47] |
刘魁英, 赵宗芸. 木质部输导阻力对苹果树体矮化的影响. 北京农学院学报, 1992, 7(2): 139-144. DOI:10.13473/j.cnki.issn.1002-3186.1992.02.018 |
| [48] |
赵琦琳, 田文斌, 郑忠, 史青茹, 由文辉, 阎恩荣. 浙江天童木本植物水力结构与树高的关联性. 生态学报, 2020, 40(19): 6905-6911. |