 2023, Vol. 43
2023, Vol. 43文章信息
- 束承继, 蔡文博, 杜凯伟, 江南, 林立, 李先源
- SHU Chengji, CAI Wenbo, DU Kaiwei, JIANG Nan, LIN Li, LI Xianyuan
- 深圳市入侵植物区系与风险分布
- Study on the flora and risk distribution of dominant invasive plants in Shenzhen
- 生态学报. 2023, 43(3): 1113-1125
- Acta Ecologica Sinica. 2023, 43(3): 1113-1125
- http://dx.doi.org/10.5846/stxb202108042129
-
文章历史
- 收稿日期: 2021-08-04
- 网络出版日期: 2022-10-10





2. 中国科学院生态环境研究中心, 城市与区域生态国家重点实验室, 北京 100085;
3. 重庆人文科技学院, 建筑与设计学院, 重庆 401573;
4. 福建农林大学园林学院, 福州 350002
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
3. College of Architecture and Design, Chongqing College of Humanities, Science & Technology, Chongqing 401573, China;
4. College of horticulture, Fujian Agriculture and Forestry University, Fuzhou 350002, China
生物入侵是一个世界性的生态问题, 随着我国城市化的发展, 外来植物入侵问题愈演愈烈。生物入侵(biological invasion)是指某种生物由原来的自然分布区域扩展到一个新的(通常也是遥远的)地区, 在新的地区里, 其后代可以繁殖、扩散并维持下去[1], 入侵植物扩散问题已然成为一个世界性的生态问题, 外来入侵植物在当地适宜水热条件的助长下, 凭借其极强的生态适应能力、繁殖能力、生长发育速度和产生的化感物质[2], 严重危害当地原生植被群落的生物多样性, 从而对当地的林业经济[3—4]、牧业经济[5]、果树经济[6]、生态环境质量[7]造成巨大损失。随着我国城市化的发展, 入侵植物也逐渐在我国各大城市扩散, 甚至形成斑块内优势种, 相关研究一直是学界关注的重点, 侯新星等[8]对江苏外来入侵植物区系、生活型及繁殖特性进行了研究, 为进一步的防治工作提供了技术支撑;周先叶等[9]调查了重点入侵植物薇甘菊在广东的传播途径及危害状况, 发现长途交通运输等人为因素是造成薇甘菊远距离传播的主要原因, 且薇甘菊对灌木危害极大。
解决外来植物入侵问题的技术前提是要实现入侵植物特征的透明化和入侵风险分布的空间化。目前对于入侵植物的研究, 内容上大多集中于区域内几种典型入侵植物的生理研究[10], 或是基于样点仅做入侵植物区系研究, 或是着眼于个别典型入侵植物的入侵机制和防治策略研究[11—14];方法上部分学者基于文献分析进行全国大尺度范围内的入侵植物风险评估研究[15—16], 或结合样点数据, 进行部分区域定性的入侵风险鉴定, 如野外通过定点调查、线路调查、文献分析相结合的方法, 记录和统计发现的外来入侵植物的种类, 在有限区域内研究入侵植物的风险分布[17], 缺少城市整体尺度范围内关于入侵植物风险值空间化的研究。
深圳是我国高度城市化的窗口城市, 城市发展带动了当地园林绿化部门和相关科研单位的引种驯化工作。持续从国外引入的园林栽培植物, 使得深圳市外来植物种类不断增加, 加之华南地区亚热带季风气候的水热条件适合入侵植物的扩散, 深圳市在外来种入侵方面面临的压力增大, 这使得深圳成为我国外来种入侵的脆弱地区, 同时也是我国外来植物入侵最为严重的地区之一[18]。所以本文在植被普查的基础上, 旨在研究深圳全市优势入侵植物区系及地理成分来源特征;构建入侵风险评估公式, 计算深圳全市斑块、种群入侵风险值并进行空间化制图。
1 研究方法 1.1 研究区概况深圳市陆域面积1997.47km2, 位于广东省南部沿海, 珠江入海口东侧, 南邻香港, 北靠东莞, 东接惠州, 是中国最早改革开放的窗口, 粤港澳大湾区四大中心城市之一, 也是国家支持建设的中国特色社会主义先行示范区。深圳全域地势由东南向西北逐渐降低, 地面坡度缓和, 整体形状为南北窄、东西宽的狭长形, 由北向南逐渐分布着谷地丘陵、山脉海岸和海湾半岛三种地形[19]。深圳气候属于南亚热带季风气候区, 东南偏东风为常年主导风向, 雨季集中在每年的4—9月份, 雨量充足, 年均降雨量1933.3mm;日照时间长2120.5h, 年平均气温22.4℃, 适合植物的全年生长和逸生入侵植物的扩散。
1.2 研究方法 1.2.1 调查方法本研究是在深圳陆域优势植被普查项目的基础上, 对其优势植物普查结果就入侵风险进行更深一步的研究分析。
利用高分辨率遥感卫星图片与地面实地调查相结合的方法对深圳陆域优势植被开展了快速普查[20]:根据手持0.5m空间分辨率的2018年谷歌卫星影像, 识别卫星图上俯视视角下城区大于25m2的纹理异质性斑块(即面积大于25m2的绿地斑块被标记识别, 纹理异质的斑块被分开识别), 识别郊区大于100m2的纹理异质性斑块;并到现场对每个斑块目视识别其中盖度排名前5, 且累计盖度超过80%的植被物种, 并按照盖度大小顺序记录为S1、S2、S3、S4和S5。经调查, 最终共划分植被群丛斑块83834个, 统计优势植物物种182科858属1443种[20]。
1.2.2 入侵物种判定与分析方法(1) 入侵植物及入侵等级的判定方法主要是依据《中国外来入侵植物名录》[21]、《中国外来入侵植物彩色图鉴》等[22]相关论著和部分学者的局地研究成果[8, 23—27], 对深圳陆域植被调查的1443种优势植物进行比对、遴选、入侵等级判定, 同时以国家有关政策法规文件作为补充。
(2) 入侵植物科、属的数量级别统计分析主要依据李锡文[28]对于种子植物科、属的大小顺序排列方法, 根据各科、各属所含种数的多少, 将各科、属进行大小科、大小属的划分;入侵种子植物科、属的地理成分分析主要依据吴征镒[29—30]关于世界种子植物科的分布区划分标准和关于中国种子植物属的分布区类型划分标准, 参照生态因素和地质因素, 可将世界所有科划分为18个大分布区类型, 中国种子植物属划分为15个分布区类型。
(3) 入侵植物来源特点分析主要依据《中国外来入侵植物名录》[21]中对于原产地的论述, 且对于原产地的划分按照地理学中七大洲的划分区域, 对于原产于美洲的入侵植物, 统计时南美洲与北美洲各计数一次;对于原产于热带美洲的入侵植物, 统计时归于南美洲计数;对于原产于地中海地区的入侵植物, 统计时, 则亚洲、欧洲和非洲各计数一次;对于原产于某个具体国家的入侵植物, 统计时则按照国家主体所在的大洲进行计数。
1.2.3 植被斑块风险值评估方法本文基于深圳陆域项目所取得的基础数据[20], 采取专家意见法, 在征询相关研究领域专家的前提下, 综合考虑“斑块面积”、“风险等级”、“斑块内入侵植物影响力总和”3种变量, 构建植被斑块风险值(VPRV, Vegetation patch risk values)评估公式, 研究深圳全域入侵植物风险的空间分布。其中为了使计算结果更具有直观性, 考虑到斑块之间面积差值较大, 令所有斑块的面积在范围1—35之间进行重分布缩放处理, 得到斑块面积指数;选择斑块内部入侵等级最高的植物, 将其入侵等级作为斑块风险等级, 1级风险最高, 7级风险最低, 且1—7级的入侵等级分别赋值35、30、25、20、15、10和5参与计公式计算;斑块内入侵植物影响力为斑块内每个入侵植物所处S1—S5中各个优势位置所对应的不同危害影响力赋值, 优势度越高危害影响力越高, 反之则越低, 赋值分别为35、28、21、14、7, 斑块内存在多种入侵植物时, 便求其赋值总和作为影响力数值;运用Excel中计算排序功能和ArcGIS软件分别对所有受入侵植物影响的群丛斑块进行风险值计算和空间分布图的制作。斑块风险值为该植被斑块受入侵植物影响的程度, 其值的大小仅作为各斑块间风险值排序的依据, 不具有实际的风险量化意义。公式如下:
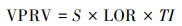
|
式中, VPRV为斑块风险值, 无量纲;S为斑块面积指数, 无量纲;LOR为斑块风险等级, 无量纲;TI为斑块内入侵植物影响力总和, 无量纲。
1.2.4 物种种群风险值评估方法种群风险值(PRV, Population risk values)为各物种种群入侵风险的大小, 采取专家意见法, 在征询相关研究领域专家的前提下, 构建物种种群风险值评估公式。综合考虑每一种入侵植物的入侵等级、入侵面积和在所有斑块内优势位置, 对深圳258种优势入侵植物进行种群风险值计算, 同样, 值的大小仅作为各物种种群间风险值排序的依据, 不具有实际的风险量化意义。公式如下:

|
式中, PRV为种群风险值, 无量纲;S为种群总面积指数, 无量纲;IIL为个体入侵等级, 无量纲;SIII为个体影响力总和, 无量纲。
种群总面积指数为包含该入侵植物种的斑块面积指数总和, 此处的斑块面积指数计算方法见本文1.2.3;个体入侵等级为该入侵植物种在《中国外来入侵植物名录》[21]中规定的入侵等级;个体影响力总和为该入侵植物种在含有它的每个群丛斑块中, 所处S1—S5的某一位置所对应的影响力总和, S1—S5各自对应的影响力赋值参考1.2.3。经Excel计算、统计、排序、分析可得出深圳地区个体风险值排名前10的入侵植物。
2 结果与分析 2.1 入侵物种区系组成分析经统计, 初步遴选出深圳陆域范围内入侵植物258种, 隶属于63科、200属;其中蕨类植物2科、2属、2种, 分别占入侵植物总科、属、种总数的3.2%、1.0%和0.8%;被子植物61科、198属、256种, 分别占入侵植物总科、属、种总数的96.8%、99.0%和99.2%;入侵植物中未发现裸子植物。总体上, 被子植物在入侵植物中占绝对优势(见表 1)。
| 门类Types | 科数 Number of families |
所占比例% Proportion |
属数 Numberof genera |
所占比例/% Proportion |
种数 Numberof species |
所占比例/% Proportion |
| 蕨类植物Pteridophyte | 2 | 3.2 | 2 | 1.0 | 2 | 0.8 |
| 被子植物Angiosperm | 61 | 96.8 | 198 | 99.0 | 256 | 99.2 |
| 总计Sum | 63 | 100.00 | 200 | 100.00 | 258 | 100.00 |
在258种入侵植物中, 豆科、菊科、禾本科所含植物种类数量分别为39种、38种、29种, 占据前三, 共计106种, 分别占入侵植物总种数的15.1%、14.7%和11.2%, 共计占比41.0%(见表 2)。相较于其他植物, 这3科植物不仅在入侵植物的种类数量上占据绝对优势, 对本地生态系统质量构成的威胁也较大。其他科总占比高达20.54%, 且每科只包含1—2种入侵植物种, 如紫茉莉科、桑寄生科等。
| 科名 Family |
种数 Number of species |
占总种数比例 Proportion/% |
科名 Family |
种数 Number of species |
占总种数比例 Proportion/% |
|
| 豆科Fabaceae | 39 | 15.12 | 爵床科Acanthaceae | 2 | 0.78 | |
| 菊科Asteraceae | 38 | 14.73 | 凤仙花科Balsaminaceae | 2 | 0.78 | |
| 禾本科Poaceae | 29 | 11.24 | 酢浆草科Oxalidaceae | 1 | 0.39 | |
| 苋科Amaranthaceae | 12 | 4.65 | 竹芋科Marantaceae | 1 | 0.39 | |
| 大戟科Euphorbiaceae | 12 | 4.65 | 樟科Lauraceae | 1 | 0.39 | |
| 旋花科Convolvulaceae | 11 | 4.26 | 鸢尾科Iridaceae | 1 | 0.39 | |
| 茄科Solanaceae | 9 | 3.49 | 雨久花科Pontederiaceae | 1 | 0.39 | |
| 唇形科Labiatae | 7 | 2.71 | 野牡丹科Melastomataceae | 1 | 0.39 | |
| 莎草科Cyperaceae | 5 | 1.94 | 荨麻科Urticaceae | 1 | 0.39 | |
| 桃金娘科Myrtaceae | 4 | 1.55 | 悬铃木科Platanaceae | 1 | 0.39 | |
| 茜草科Rubiaceae | 4 | 1.55 | 无患子科Sapindaceae | 1 | 0.39 | |
| 马鞭草科Verbenaceae | 4 | 1.55 | 天南星科Araceae | 1 | 0.39 | |
| 锦葵科Malvaceae | 4 | 1.55 | 石竹科Caryophyllaceae | 1 | 0.39 | |
| 紫葳科Bignoniaceae | 3 | 1.16 | 商陆科Phytolaccaceae | 1 | 0.39 | |
| 鸭跖草科Commelinaceae | 3 | 1.16 | 桑科Moraceae | 1 | 0.39 | |
| 仙人掌科Cactaceae | 3 | 1.16 | 桑寄生科Loranthaceae | 1 | 0.39 | |
| 西番莲科Passifloraceae | 3 | 1.16 | 秋海棠科Begoniaceae | 1 | 0.39 | |
| 马齿苋科Portulacaceae | 3 | 1.16 | 千屈菜科Lythraceae | 1 | 0.39 | |
| 龙舌兰科Agavaceae | 3 | 1.16 | 漆树科Anacardiaceae | 1 | 0.39 | |
| 蓼科Polygonaceae | 3 | 1.16 | 木贼科Equisetaceae | 1 | 0.39 | |
| 夹竹桃科Apocynaceae | 3 | 1.16 | 木麻黄科Casuarinaceae | 1 | 0.39 | |
| 葫芦科Cucurbitaceae | 3 | 1.16 | 美人蕉科Cannaceae | 1 | 0.39 | |
| 紫茉莉科Nyctaginaceae | 2 | 0.78 | 毛茛科Ranunculaceae | 1 | 0.39 | |
| 玄参科Scrophulariaceae | 2 | 0.78 | 牻牛儿苗科Geraniaceae | 1 | 0.39 | |
| 石蒜科Amaryllidaceae | 2 | 0.78 | 萝藦科Asclepiadaceae | 1 | 0.39 | |
| 十字花科Cruciferae | 2 | 0.78 | 卷柏科Selaginellaceae | 1 | 0.39 | |
| 伞形科Umbelliferae | 2 | 0.78 | 桔梗科Campanulaceae | 1 | 0.39 | |
| 葡萄科Vitaceae | 2 | 0.78 | 景天科Crassulaceae | 1 | 0.39 | |
| 木棉科Bombacaceae | 2 | 0.78 | 海桑科Sonneratiaceae | 1 | 0.39 | |
| 落葵科Basellaceae | 2 | 0.78 | 百合科Liliaceae | 1 | 0.39 | |
| 柳叶菜科Onagraceae | 2 | 0.78 | 白花丹科Plumbaginaceae | 1 | 0.39 | |
| 藜科 | Chenopodiaceae | 2 | 0.78 |
查阅国家政府部门发布的关于入侵生物的政策法规性文件(见表 3), 将其中所包含的需重点关注入侵植物种类与深圳陆域优势植物种类进行对比。结果如下:
| 政策法规文件 Policy and Regulatory documents |
存在种数 Number of species |
代表性植物 Representative plants |
| 环保部四批入侵生物名录(39) The Ministry of Environmental Protection lists four batches of invasive species |
18 | 五爪金龙Ipomoea cairica、凤眼蓝Eichhornia crassipes、薇甘菊Mikania micrantha等 |
| 国家进境植物检疫性有害生物名录(31) List of Phytosanitary Pests of the People′s Republic of China |
5 | 菟丝子Cuscuta chinensis、飞机草Chromolaena odorata、薇甘菊Mikania micrantha等 |
| 林业局《全国林业检疫性有害生物名单》(7) National List of Quarantine Pests of Forestry by Forestry Administration |
4 | 无根藤Cassytha filiformis、菟丝子Cuscuta chinensis、五爪金龙Ipomoea cairica等 |
| 农业农村部《国家重点管理外来物种名录(第一批)》(21) List of Foreign Species under National Key Management (First Batch), Ministry of Agriculture and Rural Affairs |
9 | 喜旱莲子草Alternanthera philoxeroides、三裂叶豚草Ambrosia trifida、薇甘菊Mikania micrantha等 |
数据上看, 环保部四批入侵生物名录、林业局《全国林业检疫性有害生物名单》、农业农村部《国家重点管理外来物种名录(第一批)》三份文件中涉及的入侵植物, 近一半分布于深圳, 占比分别为46.2%、57.1%、42.9%, 具有一定的分布优势, 深圳市相关部门在各自的行政施策过程中需引起重视。
2.2 入侵植物科、属数量级别统计及分析参考李锡文[28]对于种子植物科的大小顺序排列方法, 考虑到各植物科所含有的植物种类数量, 深圳陆域入侵植物的63个科可划分为5种类型, 分别为单种科(1种)、少种科(2—5种)、中型科(6—10种)、较大科(11—20种)和大型科(>20种)(见表 4)。其中单种科和少种科分别为29科(百合科、卷柏科、毛茛科、酢浆草科等)和26科(藜科、十字花科、茜草科、莎草科等), 共计55科, 两种类别分别占总科数比例46.0%和41.3%, 共占比87.3%;中型科、较大科和大型科所含科数较少, 分别为2科(唇形科、茄科)、3科(旋花科、大戟科、苋科)和3科(豆科、菊科、禾本科), 共计8科, 三种类别分别占总科数比例3.1%、4.8%和4.8%, 共占比12.7%。不难发现, 单种科与少种科在科、属的水平上集中分布, 大型科虽然只有3科, 却在种的水平上分布了106种, 占比高达41.1%, 说明, 深圳全域入侵植物区系内部结构较为复杂, 种对于科的分布存在集聚现象。
| 分级标准 The classification standard |
数量特征Quantitative characteristics | |||||
| 科数 Number of families |
占总科数比例 Proportion/% |
属数 Number of genera |
占总属数比例 Proportion/% |
种数 Number of species |
占总种数比例 Proportion/% |
|
| 单种科(1种) Single-species families |
29 | 46.0 | 29 | 14.5 | 29 | 11.2 |
| 少种科(2—5种) Few-species families |
26 | 41.3 | 58 | 29.0 | 72 | 27.9 |
| 中型科(6—10种) Medium-species family |
2 | 3.1 | 12 | 6.0 | 16 | 6.2 |
| 较大科(11—20种) Larger-species family |
3 | 4.8 | 14 | 7.0 | 35 | 13.6 |
| 大型科(>20种) Gigantic-species family |
3 | 4.8 | 87 | 43.5 | 106 | 41.1 |
| 总计Sum | 63 | 100.0 | 200 | 100.0 | 258 | 100.0 |
参考李锡文[28]对于种子植物属的大小顺序排列方法, 考虑到各植物属所含有的植物种类数量, 深圳陆域入侵植物的200个属可划分为5种类型, 分别为单种属(1种)、少种属(2—4种)、中型属(5—8种)、较大属(9—12种)和大型属(>13种)(表 5)。其中单种属高达167属, 主要为野甘草属、鼠尾草属、芒属、藜属、藿香蓟属、鬼针草属、假泽兰属等, 占总属数比例为83.5%;少种属29属, 主要为猪屎豆属、马缨丹属、狼尾草属、飞蓬属、耳草属等, 占总属数比例为14.5%;中型属4属, 分别为苋属、决明属、番薯属和大戟属, 占总属数比例为2.0%;入侵植物种缺乏较大属和大型属。从上可以看出, 单种属在深圳入侵植物区系中占绝对优势, 远高于其他类型之和, 说明该区系缺乏多样性, 另外在中型属中, 属种比为1:6, 说明中型属在种的水平上具有一定的丰富度。
| 分级标准 The classification standard |
数量特征Quantitative characteristics | |||
| 属数 Number of genera |
占总属数比例 Proportion/% |
种数 Number of species |
占总种数比例 Proportion/% |
|
| 单种属(1种)Single-species genus | 167 | 83.5 | 167 | 64.7 |
| 少种属(2—4种)Few-species genus | 29 | 14.5 | 67 | 26.0 |
| 中型属(5—8种)Medium-species genus | 4 | 2.0 | 24 | 9.3 |
| 较大属(9—12种)Larger-species genus | 0 | 0.0 | 0 | 0.0 |
| 大型属(>13种)Gigantic-species genus | 0 | 0.0 | 0 | 0.0 |
| 总计Sum | 200 | 100.0 | 258 | 100.0 |
根据吴征镒[29]关于世界种子植物科的分布区划分标准, 深圳陆域入侵植物中共有种子植物61科, 可划分为6个分布区(表 6)。世界分布科对入侵植物区系的性质反应作用不大, 故将其排除以计算其他分布类型科的占比[8]。其中泛热带分布科数量占绝对优势, 主要为豆科、天南星科、野牡丹科、西番莲科等, 占总科数的76.4%(除去世界分布科);东亚(热带、亚热带)及热带南美间断分布仅有4科, 分别为马鞭草科、紫茉莉科、仙人掌科和龙舌兰科, 占总科数的10.5%;旧世界热带分布和热带亚洲至热带大洋洲分布均仅有1科, 分别为海桑科和木麻黄科, 各自占比2.6%;热带分布共计35科, 占总科数的92.1%, 热带区系属性非常明显。深圳入侵植物中, 温带分布仅有3科, 分别为百合科、悬铃木科和牻牛儿苗科, 占比7.9%(见表 6)。
| 分布区类型 Areal-types |
科数 Number of families |
占本区科比例/% Proportion |
属数 Number of genera |
占本区属比例/% Proportion |
| 1.世界分布Widespread | 23 | — | 25 | — |
| 2.泛热带分布Pantropic | 29 | 76.4 | 62 | 35.8 |
| 3.热带亚洲和热带美洲间断分布 Trop. Asia &Trop.Amer.disjuncted |
4 | 10.5 | 34 | 19.7 |
| 4.旧世界热带分布Old World Tropics | 1 | 2.6 | 15 | 8.7 |
| 5.热带亚洲至热带大洋洲分布 Trop.Asia to Trop.Australasia Oceania |
1 | 2.6 | 3 | 1.7 |
| 6.热带亚洲至热带非洲分布 Trop.Asia to Trop.Africa |
— | — | 10 | 5.8 |
| 7.热带亚洲分布(印度-马来西亚)Trop.Asia | — | — | 7 | 4.1 |
| 8.北温带分布N.Temp. | 3 | 7.9 | 14 | 8.1 |
| 9.东亚和北美洲间断分布 E.Asia & N.Amer.disjuncted |
— | — | 9 | 5.2 |
| 10.旧世界温带分布 | — | — | 12 | 6.9 |
| 12.地中海区、西亚至中亚分布Medit., W.to C.Asia | — | — | 3 | 1.7 |
| 14.东亚分布(东喜马拉雅-日本)E.Asia | — | — | 4 | 2.3 |
| 合计(不含世界分布)Sum (Not including the Widespread) |
38 | 100.0 | 173 | 100.0 |
根据吴征镒[30]关于中国种子植物属的分布区类型划分标准, 深圳陆域200属入侵种子植物可划分为12个分布区(见表 6), 占世界15个分布区类型的80.0%, 说明深圳陆域范围内的入侵种子植物属地理成分较为复杂;世界分布属对入侵植物区系的性质反应作用不大, 故将其排除以计算其他分布类型属的占比[8]。
深圳陆域入侵植物中, 热带分布属共计131属, 占总属数比例高达75.7%, 热带区系属性非常明显。其中泛热带分布62属, 约占本分布类型的47.3%, 占总属数的35.8%, 主要为马齿苋属、蟛蜞菊属、银合欢属、泽兰属、猪屎豆属、莲子草属等;热带亚洲和热带美洲间断分布34属, 约占本分布类型的26.0%, 占总属数的19.7%, 主要为凤眼莲属、野甘草属、落花生属、百日菊属、藿香蓟属、地毯草属等。这两种分布类型在热带分布中占据优势, 其中泛热带分布优势更大。
深圳陆域入侵植物中, 温带分布属共计42属, 占总属数的24.3%, 低于热带成分75.7%。其中北温带分布和旧世界温带分布两种分布类型占在温带分布中占据优势, 分别有14属和12属, 主要为芸薹属(8)、稗属(8)、夏枯草属(8)、苦苣菜属(8)、唐松草属(8)、雏菊属(10)、萱草属(10)、鹅肠菜属(10)、苜蓿属(10)等。温带区系属性的入侵种子植物在本地区依然占有一定比例(见表 6), 可能较为适应当地的水热气候条件。
2.3.2 入侵植物来源特点分析按照上述原产地划分原则, 统计得258种深圳陆域入侵植物共计302频次。原产于亚洲的入侵植物频次高达128次, 占比最高, 为42.4%;其次是原产于南美洲的入侵植物, 为76频次, 占比约25.2%;原产于北美洲的入侵植物凭借48频次, 占比15.9%位列第三;原产于非洲、欧洲、大洋洲的入侵植物频次分别为26、12和7, 分别占比8.6%、4.0%和2.3%。除以上起源地外, 另有3种入侵植物起源不详, 两种入侵植物为杂交起源, 占比均不足1%(图 1)。

|
| 图 1 深圳入侵植物原产地统计 Fig. 1 Origin statistics of invasive plants in Shenzhen |
原产于亚洲的代表性入侵植物有棕叶狗尾草Setaria palmifolia、节节草Equisetum ramosissimum、翠云草Selaginella uncinata、鳢肠Eclipta prostrata、蓝猪耳Torenia fournieri、凤仙花Impatiens balsamina等;原产于南美洲的代表性入侵植物有凤眼蓝Eichhornia crassipes、南美蟛蜞菊Sphagneticola trilobata、喜旱莲子草Alternanthera philoxeroides、光荚含羞草Mimosa bimucronata、假臭草Praxelis clematidea、皱果苋Amaranthus viridis、金腰箭Synedrella nodiflora、马缨丹Lantana camara等;原产于北美洲的代表性入侵植物有薇甘菊Mikania micrantha、五叶地锦Parthenocissus quinquefolia、小蓬草Erigeron canadensis、三裂叶豚草Ambrosia trifida、飞机草Chromolaena odorata、垂序商陆Phytolacca americana等;原产于非洲的代表性入侵植物有苏丹凤仙花Impatiens walleriana、蓖麻Ricinus communis、光萼猪屎豆Crotalaria trichotoma、落地生根Bryophyllum pinnatum、象草Pennisetum purpureum等;原产于欧洲的代表性入侵植物有猫尾草Uraria crinita、铺地黍Panicum repens、黑麦草Lolium perenne、凹头苋Amaranthus blitum等;原产于大洋洲的代表性入侵植物有桉Eucalyptus robusta、大叶相思Acacia auriculiformis、木麻黄Casuarina equisetifolia、田菁Sesbania cannabina、黄槐决明Senna surattensis等;芥菜Brassica juncea、金盏花Calendula officinalis和金凤花Impatiens cyathiflora三个物种起源不详;唐菖蒲、碧冬茄两种植物为杂交起源。
总体来说, 深圳作为一座国际人口流动、贸易频繁的城市, 陆域入侵植物中还是以原产亚洲本地的植物物种为主, 但原产于南、北美洲的入侵植物也能占据一定优势, 说明原产于这三个大洲的入侵植物能较好的适应深圳当地的亚热带海洋性气候。
2.4 深圳市植被斑块风险值评估及分布深圳陆域优势植被普查工作将深圳几乎全域植被覆盖共划分成83834个群丛斑块, 通过对斑块内部优势植物的鉴别与遴选, 共得到受入侵植物影响的植被群丛斑块48852个。
经斑块风险值(VPRV, Vegetation patch risk values)的计算、比较、栅格化处理、自然断点法分级, 将深圳全域植被斑块风险分为7级, 1级最高, 7级最低。从图 2可以看出, 受入侵植物影响高风险值区域主要集中在郊区或近自然地带, 以1、2级为主。这些区域多位于航运交通枢纽、人类活动干扰少、雨热条件好、行政界线等处, 为入侵植物的进入和扩散提供了有利条件。由东南向西北分别为, 大鹏半岛七娘山片区的三角山、大雁顶、鹿嘴山庄、大鹏坳区域;坪山区的赤澳水库、田头山、石牙头、火烧天区域;盐田区的东部华侨城、梧桐山区域;龙岗区的松子坑水库、十二岭顶、屋头角、洞其寨区域;罗湖区的深圳水库、梧桐山区域;福田区的塘朗山部分区域;宝安区的铁岗水库区域;龙华区的分水坳、石峡岭区域;龙华区与东莞交界的大屏障森林公园区域。由于城区植物生长环境受人类活动干扰较大, 以及相关部门园林养护工作的有序推进, 城区斑块的入侵植物风险值普遍较低(图 2)。

|
| 图 2 深圳入侵植物风险值分级图 Fig. 2 A risk value distribution map of invasive plants in Shenzhen |
综合考虑每一种入侵植物的入侵等级、入侵面积和在所有入侵斑块内优势位置, 采取种群风险值(PRV, Population risk value)计算公式对深圳258种入侵植物进行种群风险值计算、比较, 可得出深圳地区种群风险值排名前13的入侵植物(表 7)。
| 排序 Order |
物种 Species |
科名 Family |
生活型 Life form |
排序 Order |
物种 Species |
科名 Family |
生活型 Life form |
|
| 1 | 白花鬼针草Bidens pilosa | Asteraceae | 一年生草本 | 8 | 叶子花Bougainvillea spectabilis | Nyctaginaceae | 灌木 | |
| 2 | 薇甘菊Mikania micrantha | Asteraceae | 多年生草本 | 9 | 糖胶树Alstonia scholaris | Apocynaceae | 乔木 | |
| 3 | 银合欢Leucaena leucocephala | Fabaceae | 灌木或小乔木 | 10 | 地毯草Axonopus compressus | Poaceae | 多年生草本 | |
| 4 | 南美蟛蜞菊 Sphagneticola trilobata |
Asteraceae | 多年生草本 | 11 | 五爪金龙Ipomoea cairica | Convolvulaceae | 多年生草本 | |
| 5 | 桉Eucalyptus robusta Smith | Myrtaceae | 乔木 | 12 | 马缨丹Lantana camara | Verbenaceae | 灌木 | |
| 6 | 杧果Mangifera indica | Anacardiaceae | 乔木 | 13 | 光荚含羞草Mimosa bimucronata | Fabaceae | 灌木 | |
| 7 | 龙眼Dimocarpus longan | Sapindaceae | 乔木 | |||||
入侵植物的入侵途径一般可以分为有意引入、无意引入和自然扩散3类[25]。
结合上文国家相关政策法规中所提及的高风险入侵植物, 决定对白花鬼针草Bidens pilosa、薇甘菊Mikania micrantha、银合欢Leucaena leucocephala、南美蟛蜞菊Sphagneticola trilobata、桉Eucalyptus robusta、五爪金龙Ipomoea cairica、马缨丹Lantana camara、光荚含羞草Mimosa bimucronata这8种入侵植物的入侵途径及其他相关信息进行分析:
这8种高风险入侵植物中, 银合欢、南美蟛蜞菊、桉、五爪金龙、马缨丹、光荚含羞草均是出于提升园林绿化的目的有意引入的, 后逸生才形成了大范围的恶意入侵分布, 白花鬼针草和薇甘菊为无意引入。这8种植物中有7种原产于美洲大陆, 仅桉一种原产于大洋洲, 结合图 1可知原产于美洲大陆的入侵植物对深圳当地的生态环境危害较大。这些植物具有极强的繁殖能力和竞争能力。薇甘菊、南美蟛蜞菊依靠茎节间不定根可实现快速无性繁殖[9, 31], 同时和五爪金龙、马缨丹一起均具有化感作用, 抑制其他植物的正常生长发育[31—34];白花鬼针草产种率、结实量、种子萌发率均处于高位, 短时间内便可形成大面积种群[13], 危害农作物生长;有“抽水机”之称的桉树生长过程中消耗地力速度过快, 破坏林地水分平衡, 根部还会产生抑制其他植物生长的有毒分泌物[35];光荚含羞草有极强的适应性、抗旱、耐瘠薄和再生能力, 在野外广发分布, 会对当地的生态环境和生物多样性造成严重破坏[36];银合欢极强的繁殖能力和环境适应能力使其在逸生状态下易疯长, 破坏种间平衡[37]。表 7中6—10名植物均为深圳地区常见的经济果树或园林绿化植物, 主动扩散能力较弱, 分布在可控的范围内, 暂不作讨论。由上可知, 我们在今后的引种驯化过程中, 要坚持以科学为导向, 在摸清入侵植物整体风险值分布格局的情况下, 深入研究目标物种潜在的入侵机制和生理特性, 同时在园林应用过程中, 针对高风险值的入侵植物种群要重点开展防治工作。
3 结论(1) 深圳陆域范围内共有63科200属258种入侵植物, 其中被子植物占有绝对的分布优势;豆科、菊科、禾本科为所有植物科中占比较高的三大科;对比四类国家相关政策法规文件, 环保部四批入侵生物名录、林业局《全国林业检疫性有害生物名单》、农业农村部《国家重点管理外来物种名录(第一批)》三份文件中所提及的入侵植物, 近一半出现在了深圳。
(2) 单种科和少种科占比较高, 在科、属的水平上集中分布, 深圳全域入侵植物区系内部结构较为复杂, 种对于科的分布存在集聚现象;单种属在深圳入侵植物区系中占绝对优势, 远高于其他类型之和, 该区系缺乏多样性, 另外在中型属中, 属种比为1:6, 说明中型属在种的水平上具有一定的丰富度。
(3) 入侵植物科、属的地理成分具有相似性, 热带区系属性非常明显;温带分布依然占有一定比例, 可能是由于部分温带区系属性的入侵种子植物较为适应当地的水热气候条件;此外深圳陆域范围内的入侵种子植物属地理成分较为复杂。
(4) 入侵植物以原产亚洲本地的植物物种为主, 但原产于南、北美洲的入侵植物也能占据一定优势, 说明原产于这三个大洲的入侵植物能较好的适应深圳当地的亚热带海洋性气候。
(5) 深圳范围内入侵植物高风险区主要分布在临近港口、人类活动干预少、行政区交界处、水热条件好的郊区或近自然地带, 由东南向西北分别涉及大鹏半岛、坪山区、盐田区、龙岗区、宝安区、龙华区、龙华区与东莞交界的大屏障森林公园区域等范围, 具体位置分布可见图 2。城区受园林养护和人为活动的影响, 风险值较低。
(6) 经公式计算可得入侵风险值排前十的入侵植物, 综合考虑国家相关政策法规文件和植物实际园林应用情况, 文中对白花鬼针草、薇甘菊、银合欢、南美蟛蜞菊、桉、五爪金龙、马缨丹和光荚含羞草这8类高风险值入侵植物进行了入侵途径及危害分析, 发现这8种植物多为园林绿化目的人为引入, 后逸生泛滥成灾。
4 讨论与防治建议 4.1 讨论入侵植物极大的阻碍了本地生态环境发挥其应有的生态功能, 如何全面而系统的对区域内入侵植物现状进行评估和地理空间化是一大研究难点, 也是面向实际植被管理应用时的亟需研究方向。相较于传统的文献分析法和样点调查法中结果不全面的局限性, 本文基于植被普查的方法所得数据[20], 探索性地引入植被斑块风险值评估方法和物种种群风险值评估方法, 从优势入侵植物层面, 对于地级市陆域范围内总体入侵植物的地理分布、来源特点、入侵途径、种群风险、全域风险分布等进行了研究。
优势植被普查方法[20]所获得的基础数据更为全面、详实;斑块风险值计算方法和种群风险值计算方法, 充分征求专家意见, 考虑了分布面积、入侵等级、入侵植物影响力总和3个参数, 在城市全域入侵植物风险空间化研究中具有探索性, 为未来研究中大尺度区域入侵植物风险分布的研究打下方法基础;基于此方法的研究结果准确性较高, 与深圳市生态环境局发布的《深圳市生物多样性白皮书》[38]和深圳市环境科学研究院在《深圳市陆域生态调查评估报告》中发布的入侵植物样地调查结果。
国际上对入侵植物风险地理空间分布的研究也存在其他方法, 例如Jenica M. Allen等[39]曾基于丰富全面入侵植物数据库和最大熵模型对全美大陆入侵植物的地理空间分布情况进行了长时间序列的对比研究并绘制了分布图, Rodríguez-Merino等[40]同样参考已有入侵植物文献利用最大熵模型对欧洲的水生植物入侵风险进行了研究并绘制了分布图, 但与本文基于实际本底调查获得的地理空间分布数据相比, 均不具有精度优势, 不过, 实地全域植被普查周期较长, 成本较高, 将来可结合遥感技术、树种识别、机器学习等方法开展深入研究, 实现年际间结果的快速获取。
从结果看, 深圳市入侵植物现状不容乐观。亚热带地区良好的水热因子为热带区系属性明显的入侵植物扩散创造了条件;不过根据数量级别分析可知, 深圳入侵植物种在科、属的分类水平上较为集聚, 依据同科或同属植物生理特征、植株形态、生活型、繁殖方式、内部结构以及基因的相似性, 在充分研究的基础上可进行整体防治;另外深圳作为国际化窗口城市, 应加强各个出境口岸、港口、机场等地区的检疫工作。
4.2 防治建议(1) 总体观原则。在全面排查的基础上, 深圳各行政区相关部门应建立入侵植物管理数据库, 总体上对本区入侵植物的种类、入侵机制、生理特性、来源特点、扩散分布等特点进行研究并记录, 加强深圳不同行政区、不同部门之间的工作协调度实施入侵植物动态监测。
(2) 突出重点。对现状入侵危害严重、入侵风险高、扩散速度快、分布范围广的入侵植物要重点治理, 制定具有针对性的防治策略。如在深圳清林径水库、铁岗水库、排牙山、梧桐山、塘朗山等地区广布的草本入侵植物薇甘菊、白花鬼针草、五爪金龙等植物。
(3) 因种施策。入侵植物防治时要考虑到不同入侵植物之间入侵机制、生理特性等方面的差异性, 因种施策, 达到精准防治、高效防治的目的。目前针对白花鬼针草, 主要采取化学除草剂防治和生物除草剂防治两种方法, 生物除草剂是指利用自然界中的生物(包括微生物、植物和动物)或其组织、代谢物工业化生产的用于除草的生物制剂[13];薇甘菊、凤眼蓝适合采取物理防治的方法, 人工清除后任其干枯。
(4) 加强检疫。扩散节点防控也是入侵植物防治工作的重要一步。多数外来入侵植物均是通过各种交通工具运输货物和全球性旅游的游客经海关口岸无意间带入的[41]。从2.4植被斑块风险值分布来看, 针对盐田区周围山体的入侵植物风险分布现状, 深圳应重点加强盐田港区进出口货物的入侵植物检疫力度。除此之外宝安机场、福田口岸、罗湖口岸、车站等外来旅客较多的地方也应重点防护, 守好防止入侵植物入侵的第一道防线。
(5) 变废为宝。随着对入侵植物的不断研究, 要挖掘其可能存在的生态修复价值和经济价值。由2.5高风险值入侵植物统计表可知, 目前深圳较为严重白花鬼针草、薇甘菊、南美蟛蜞菊等。经研究发现, 白花鬼针草不仅可以用来进行工业区受重金属污染土壤的修复[42—43], 还可用于饲料开发发展牧业经济[44], 同时白花鬼针草也有一定的药用价值, 可用于相关疾病药物的研制[45—47];薇甘菊提取物可用于入侵物种椰心叶甲的防治, 对其具有一定的胃毒活性[48];薇甘菊与南美蟛蜞菊提取液对非洲大蜗牛(深圳常见入侵生物)幼体与成体地触杀能力较强, 或许可在深圳用于非洲大蜗牛地防治工作[49];也已经有学者开始研究南美蟛蜞菊作为食用菌的栽培原料的可行性, 且结果良好, 如能顺利实现, 将变废为宝, 实现南美蟛蜞菊生物质的有效利用[50]。
| [1] |
Elton C S. The Ecology of Invasions by Animals and Plants. Chicago: University of Chicago Press, 1958.
|
| [2] |
桑卫国, 朱丽, 马克平. 外来种入侵现象、问题及研究重点. 地球科学进展, 2006, 21(3): 305-312. DOI:10.3321/j.issn:1001-8166.2006.03.013 |
| [3] |
Waterhouse D F. Biological Control of Weeds: Southeast Asian Prospects. Canberra: ACIAR, 1994: 125-126.
|
| [4] |
Waterhouse D F. The Major Arthropod Pests and Weeds of Agriculture in Southeast Asia: Distribution, Importance and Origin. Canberra: Australian Centre for International Agricultural Research, 1993, 124-135. |
| [5] |
Haysom K A, Murphy S T. The status of invasiveness of forest tree species outside their natural habitat: a global review and discussion paper. Rome: FAO, 2003: 5-8.
|
| [6] |
陈奕杉. 入侵植物紫茎泽兰(Ageratina adenophora)叶内生优势炭疽真菌(Colletotrichum spp.)对经济作物影响的研究[D]. 昆明: 云南大学, 2016.
|
| [7] |
符建伟. 外来物种入侵的生态哲学研究[D]. 苏州: 苏州科技大学, 2019.
|
| [8] |
侯新星, 辛建攀, 陆梦婷, 田如男. 江苏外来入侵植物区系、生活型及繁殖特性. 生态学杂志, 2019, 38(7): 1982-1990. DOI:10.13292/j.1000-4890.201907.026 |
| [9] |
周先叶, 昝启杰, 王勇军, 李鸣光, 廖文波, 王伯荪. 薇甘菊在广东的传播及危害状况调查. 生态科学, 2003, 22(4): 332-336. DOI:10.3969/j.issn.1008-8873.2003.04.009 |
| [10] |
王宇涛, 李春妹, 李韶山. 华南地区3种具有不同入侵性的近缘植物对低温胁迫的敏感性. 生态学报, 2013, 33(18): 5509-5515. |
| [11] |
玄祖迎, 陈雪梅, 李镇魁, 薛克娜. 佛山市主要外来入侵植物及其防治对策. 防护林科技, 2018(3): 49-51. DOI:10.13601/j.issn.1005-5215.2018.03.018 |
| [12] |
邓玲姣, 邹知明. 三叶鬼针草生长、繁殖规律与防除效果研究. 西南农业学报, 2012, 25(4): 1460-1463. DOI:10.3969/j.issn.1001-4829.2012.04.068 |
| [13] |
尚春琼, 朱珣之. 外来植物三叶鬼针草的入侵机制及其防治与利用. 草业科学, 2019, 36(1): 47-60. |
| [14] |
王宇涛, 麦菁, 李韶山, Björn L O. 华南地区严重危害入侵植物薇甘菊和五爪金龙入侵机制研究. 华南师范大学学报: 自然科学版, 2012, 44(4): 1-5. |
| [15] |
Liu J, Liang S C, Liu F H, Wang R Q, Dong M. Invasive alien plant species in China: regional distribution patterns. Diversity and Distributions, 2005, 11(4): 341-347. DOI:10.1111/j.1366-9516.2005.00162.x |
| [16] |
Xu H G, Qiang S, Genovesi P, Ding H, Wu J, Meng L, Han Z M, Miao J L, Hu B S, Guo J Y, Sun H Y, Huang C, Lei J C, Le Z F, Zhang X P, He S P, Wu Y, Zheng Z, Chen L, Jarošík V, Pyšek P. An inventory of invasive alien species in China. NeoBiota, 2012, 15: 1-26. DOI:10.3897/neobiota.15.3575 |
| [17] |
王苏铭, 张楠, 于琳倩, 赵容慧, 郝鹏, 李景文, 姜英淑, 沙海峰, 刘义, 张志翔. 北京地区外来入侵植物分布特征及其影响因素. 生态学报, 2012, 32(15): 4618-4629. |
| [18] |
邵志芳, 赵厚本, 邱少松, 杨义标, 彭少麟, 陆宠芳, 陈卓全. 深圳市主要外来入侵植物调查及治理状况. 生态环境, 2006, 15(3): 587-593. DOI:10.3969/j.issn.1674-5906.2006.03.029 |
| [19] |
唐瑜聪. 深圳市极端气候时空特征及其灾害风险评估[D]. 北京: 中国地质大学(北京), 2017.
|
| [20] |
束承继, 蔡文博, 韩宝龙, 李先源, 江南, 欧阳志云. 基于快速普查方法的深圳植被优势种特征研究. 生态学报, 2020, 40(23): 8516-8527. |
| [21] |
马金双, 李惠茹. 中国外来入侵植物名录. 北京: 高等教育出版社, 2018.
|
| [22] |
严靖, 闫小玲, 马金双. 中国外来入侵植物彩色图鉴. 上海: 上海科学技术出版社, 2016.
|
| [23] |
闫小玲, 寿海洋, 马金双. 浙江省外来入侵植物研究. 植物分类与资源学报, 2014, 36(1): 77-88. |
| [24] |
严岳鸿, 邢福武, 黄向旭, 付强, 秦新生, 陈红锋. 深圳的外来植物. 广西植物, 2004(3): 232-238. DOI:10.3969/j.issn.1000-3142.2004.03.007 |
| [25] |
曹婧, 徐晗, 潘绪斌, 戎郁萍. 中国草地外来入侵植物现状研究. 草地学报, 2020, 28(1): 1-11. |
| [26] |
林敏, 郝建华, 陈国奇. 苏州地区外来入侵植物组成及分布. 植物资源与环境学报, 2012, 21(3): 98-104. |
| [27] |
殷茜, 汪洪江, 刘兴剑. 南京中山植物园外来入侵植物分布特征及其入侵途径. 华东师范大学学报: 自然科学版, 2019(2): 128-134, 163-163. |
| [28] |
李锡文. 中国种子植物区系统计分析. 云南植物研究, 1996, 18(4): 363-384. |
| [29] |
吴征镒, 周浙昆, 李德铢, 彭华, 孙航. 世界种子植物科的分布区类型系统. 云南植物研究, 2003, 25(3): 245-257. |
| [30] |
吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1991(S4): 1-139. |
| [31] |
吴彦琼, 胡玉佳, 廖富林. 从引进到潜在入侵的植物——南美蟛蜞菊. 广西植物, 2005, 25(5): 413-418. |
| [32] |
Ismail B S, Mah L S. Effects of Mikania micrantha H.B.K. On germination and growth of weed species. Plant and Soil, 1993, 157(1): 107-113. |
| [33] |
邓业成, 孔卓玮, 骆海玉, 宁雷, 秦卉. 外来入侵植物五爪金龙对5种蔬菜种子萌发和幼苗生长的化感效应. 种子, 2010, 29(7): 8-11. |
| [34] |
李玉霞, 尚春琼, 朱珣之. 入侵植物马缨丹研究进展. 生物安全学报, 2019, 28(2): 103-110. |
| [35] |
项东云. 华南地区桉树人工林生态问题的评价. 广西林业科学, 2000, 29(2): 57-64, 86-86. |
| [36] |
蓝来娇, 马涛, 朱映, 茅裕婷, 董瑞利, 温秀军. 外来入侵植物光荚含羞草的研究进展. 河北林业科技, 2019(1): 47-52. |
| [37] |
刘国道, 何华玄. 我国银合欢的发展现状与前景. 中国草业科学, 1988(2): 56-60. |
| [38] |
深圳市生态环境局. 《深圳市生物多样性白皮书》. 2021.
|
| [39] |
Allen J M, Bradley B A. Out of the weeds? Reduced plant invasion risk with climate change in the continental United States. Biological Conservation, 2016, 203: 306-312. |
| [40] |
Rodríguez-Merino A, García-Murillo P, Cirujano S, Fernández-Zamudio R. Predicting the risk of aquatic plant invasions in Europe: how climatic factors and anthropogenic activity influence potential species distributions. Journal for Nature Conservation, 2018, 45: 58-71. |
| [41] |
李成宏, 苑景淇, 于忠亮, 周梅妹, 王梅芳, 杜凤国. 吉林省外来入侵植物的现状与防治对策. 安徽农业科学, 2019, 47(14): 51-54. |
| [42] |
赖彩秀, 潘伟斌, 张太平, 詹淑威, 吴俭. 苹果酸和草酸对两种植物吸收土壤中Cd、Zn的影响. 生态科学, 2016, 35(4): 31-37. |
| [43] |
Salazar M J, Rodriguez J H, Cid C V, Pignata M L. Auxin effects on Pb phytoextraction from polluted soils by Tegetes minuta L. and Bidens pilosa L.: extractive power of their root exudates. Journal of Hazardous Materials, 2016, 311: 63-69. |
| [44] |
邹知明, 邓玲姣, 钱雪明, 王伟, 陈建培, 陈卓, 黄串妮, 叶梦芝. 广西几种野生植物草粉饲喂家兔的试验. 饲料研究, 2011(8): 28-29. |
| [45] |
Chien S C, Young P H, Hsu Y J, Chen C H, Tien Y J, Shiu S Y, Li T H, Yang C W, Marimuthu P, Tsai L F L, Yang W C. Anti-diabetic properties of three common Bidens pilosa variants in Taiwan. Phytochemistry, 2009, 70(10): 1246-1254. |
| [46] |
Hsu Y J, Lee T H, Chang C L T, Huang Y T, Yang W C. Anti-hyperglycemic effects and mechanism of Bidens pilosa water extract. Journal of Ethnopharmacology, 2009, 122(2): 379-383. |
| [47] |
Chen Y, Luo J G, Zhang Q, Kong L Y. Identification of active substances for dually modulating the renin-angiotensin system in Bidens pilosa by liquid chromatography-mass spectrometry-based chemometrics. Journal of Functional Foods, 2016, 21: 201-211. |
| [48] |
钱军, 岑选才, 杜尚嘉, 刘盛波, 冯巧, 林作武. 薇甘菊提取物对椰心叶甲胃毒和拒食活性研究. 中国农学通报, 2015, 31(27): 258-261. |
| [49] |
邢树文, 朱慧, 查广才, 吴荣浩, 陈萍, 潘瀚, 林银芳. 四种入侵植物提取液对褐云玛瑙蜗牛的防控效果. 生态学报, 2015, 35(9): 3067-3075. |
| [50] |
张国广, 黄家福. 南美蟛蜞菊作为食用菌栽培原料的可行性探索试验. 食药用菌, 2020, 28(4): 259-261, 267-267. |