 2023, Vol. 43
2023, Vol. 43文章信息
- 马文婧, 李英年, 张法伟, 韩琳
- MA Wenjing, LI Yingnian, ZHANG Fawei, HAN Lin
- 青海湖北岸草甸草原CO2通量年际动态及其驱动机制
- Interannual dynamics and driving mechanism of CO2 flux in meadow grassland on the north shore of Qinghai Lake
- 生态学报. 2023, 43(3): 1102-1112
- Acta Ecologica Sinica. 2023, 43(3): 1102-1112
- http://dx.doi.org/10.5846/stxb202107292057
-
文章历史
- 收稿日期: 2021-07-29
- 网络出版日期: 2022-10-11





2. 成都信息工程大学大气科学学院, 成都 610200;
3. 中国科学院西北高原生物研究所, 西宁 810001
2. College of Atmospheric Sciences, Chengdu University of Information Technology, Chengdu 610200, China;
3. Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810001, China
20世纪70年代以来, 全球气温以逐10年0.17 ℃的速度上升, 扭转了过去8000年的降温趋势, 并影响了生态系统的健康[1—2]。在过去260年里, 从1750年到2018年大气CO2浓度约增加了47%[3]。因此, 碳通量作为生态系统不可或缺的一部分, 已经成为全球气候变化研究领域最广泛的课题之一。青藏高原是重要的气候调节器和生态关键区, 有“中华水塔”和“世界第三级”之称, 在当今全球气候变化情形下, 草原作为高原主体生态系统之一, 在全球生态系统碳源汇和碳循环过程中扮演着重要角色, 因此得到国内外学者的广泛关注, 成为全球变化研究领域的热点问题。草地以其高敏感性和分布面积广泛在生态系统中扮演着极其重要的作用, 高寒草地因处于强辐射、低气温地区, 常被预期为“碳汇”区, 但实质上高寒草地碳源汇状况有较大的年际变化。研究表明克氏针茅草原生态系统生长季表现为明显的碳汇[4];Barcza等测定了2年匈牙利西部半天然草地的碳通量, 均为碳汇[5];而温带草地生态系统大多数为碳平衡状态[6];高寒草地随水热等环境因子的季节分配和年际变异而发生“源-汇”和“汇-源”的转变[7], 这将对气候变化情形下碳通量的预测带来诸多的不确定性。草甸草原主要分布于青藏高原海拔相对较低, 温度相对较高的地区, 是高原重要的植被类型之一, 就维持生态系统健康和资源可持续利用的地位不容忽视[8]。因此, 研究青藏高原草甸草原生态系统的CO2通量特征及其影响因子对评价青藏高原生物地球化学循环以及全球变化的响应和反馈具有重要的科学意义[9]。通过分析2009年到2016年8年的碳通量变化规律以及环境因子的调控机制, 探讨不同时间尺度的碳通量动态变化状况, 以期为高纬度高海拔地区草地碳循环研究提供科学依据。
1 材料与方法 1.1 研究区域概述研究地点位于青海省海北牧业气象试验站观测基地, 观测场地理位置位于100°51′E, 36°57′N, 海拔高度为3140.0 m, 年均降水量为391.9 mm, 年均气温为0.5 ℃, 年日照时数为2912.7 h。该地区位于青藏高原腹地, 青海湖北岸, 为典型高原大陆性半干旱气候。其观测场地势平坦, 植被分布均匀, 下垫面为高寒草甸化草原植被。观测场植被以西北针茅(Stipa sareptana var. krylovii)为优势种, 伴生种有猪毛蒿(Artemisia scoparia)、矮嵩草(Kobresia humilis)、冷地早熟禾(Poa crymophila)、斜茎黄芪(Astragalus adsurgens)等, 植物生长末期平均高度在20—30 cm, 盖度90%以上。土质为沙壤土, 土壤发育年轻, 粗骨性强[10]。
1.2 数据观测和处理研究区CO2通量以涡度相关系统方法观测, 通量塔设置于观测场地势平坦开阔且“风浪区”足够大的中心区域。该涡度相关系统包括开路CO2/H2O快速红外分析仪(LI-7500, LI-CorInc., Nebraska, USA)和三维超声风速仪(CSAT-3, Campbell Scientific, Utah, USA)。观测高度设置为离地面2.5 m高, 采样频率为10 Hz, 每隔半小时输出一次观测平均值, 每年4月底用CO2标准气体和高纯氮对LI-7500开路CO2/H2O快速红外分析仪进行标定。所用气象要素数据为附近5 m处的同步气象观测值。由于供电间断、仪器故障、探头结露等非可控因素, 通量塔采集到的原始数据缺失或异常现象是不可避免的[11]。本文中所有通量初始数据均经过质量控制, 处理标准按照ChinaFLUX推荐的流程进行[12], 即, 先经过WPL校正[13], 再以|NEE| < 1.0 mg CO2 m-2 s-1作为阈值, 设10天数据为窗口, 3δ(标准差)为标准进行异常数据的剔除。对于夜间观测(PPFD < 10 μmol m-2 s-1)湍流发生不理想的情况, 筛除摩擦速度u* < 0.15 m/s所对应的通量数据。对于较短时间缺失(小于2小时)的通量数据, 用线性内插法进行插补处理。对于长时间缺失的通量数据, 先用有效通量数据和环境因子构建非线性回归方程, 然后利用缺失通量数据对应的环境因子和此方程完成数据插补。
本研究夜间通量数据(Reco, n)采用5 cm土壤温度(Ts)的Van′t Hoff函数(公式(1))进行插补, 生长季白天的通量数据利用与PPFD的Michaelis-Menten函数(公式(2))进行插补[14—15]。
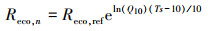
|
(1) |
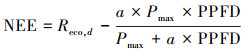
|
(2) |
式中, Reco, ref为10 ℃时生态系统的参考呼吸速率, Q10为生态系统呼吸熵(呼吸随温度升高10 ℃所增加的倍数), Reco, d为生态系统白天的呼吸速率, a和Pmax分别为生态系统表观光量子效率和潜在最大CO2同化速率(即PPFD趋向极大时生态系统净光合速率)。以上参数均为方程拟合值。
涡度相关系统无法直接测得生态系统总初级生产力(GPP)和生态系统呼吸(Re), 需要公式外推得到。涡度相关系统观测的CO2通量为NEE, 将夜间呼吸函数外延至白天(Reco, d), 就可得到生态系统的总呼吸量(Re), 进而日GPP由Re减去NEE可得生态系统总初级生产力(GPP), 如式(3)所示, 日Re为白天呼吸(Reco, d)和夜间呼吸(Reco, n)之和。NEE正负值分别代表生态系统对CO2的释放和吸收[16]。
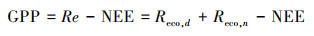
|
(3) |
数据运用SPSS 21.0和Excel 2010软件对数据进行统计分析, 用Origin 2016软件绘图。结构方程模型(SEM)是基于变量的协方差矩阵来分析变量之间关系的一种统计方法。SEM将多个变量结合在一个因果关系网络中, 从而允许同时测试多个假设。由于变量既可以是预测变量, 也可以是响应变量, 并且SEM以图形展示和数学分析相结合, 因此是一种量化直接和间接效应的有用工具。采用SEM来分析逐8日的环境因子(气温Ta、5cm土壤温度Ts、光合有效辐射PPFD、水汽饱和亏VPD、降水量Rain、增强型植被指数EVI、风速Ws、水汽压e)对CO2通量的调控机制, 用PiecewiseSEM软件包在RStudio软件中分析绘图。
2 结果与分析 2.1 主要环境因子的月际变化特征从2009年到2016年8年的环境因子的月均图(图 1)分析可得:月均气温(Ta)在-11.9—13.1 ℃之间变化, 1月平均气温最低, 从1月开始平均气温呈上升趋势, 到7月盛夏时节平均气温最高, 8月至12月平均气温逐渐降低。月均净辐射(Rn)的变化趋势与月均气温(Ta)类似, 由于太阳辐射的原因, 呈现季节性变化规律, 从1月到7月Rn呈上升趋势, 从8月至12月呈下降趋势, 1月Rn最低为17.1 W/m2, 7月最高为164.4 W/m2。月均增强型植被指数(EVI)与月均气温(Ta)和月均净辐射(Rn)的变化趋势基本一致, 变化幅度从1月开始增长到7月最高值0.5, 从8月至12月开始下降, 最小值为0.1。7月份月均降水量(Rain)最大, 为105.4 mm, 降水主要集中在生长季(5—9月), 生长季月均降水量均超过48 mm, 生长季总降水量为387.5 mm, 占全年降水量的90%, 非生长季降水量相对偏少。月均水汽饱和亏(VPD)春夏季较秋冬季高且变化幅度大, 月均水汽饱和亏(VPD)在0.18—0.59 kPa之间浮动。受降水和土壤蒸散影响, 秋冬季月均5 cm土壤含水量(SWC)稳定偏低, 春夏季月均SWC随着降水量的增大而增大且变幅较大, 秋冬季土壤较干燥, 土壤含水量在0.1 cm3/cm3以下, 春夏季SWC在0.2 cm3/cm3以上, 9月份SWC最大为0.27 cm3/cm3。

|
| 图 1 青海湖北岸草甸草原2009—2016年月均气温(Ta)、水汽饱和亏(VPD)、净辐射(Rn)、降水量(Rain)、5cm土壤体积含水量(SWC)、增强型植被指数(EVI)月际变化特征 Fig. 1 Intermonthly variation characteristics of mean air temperature (Ta), water vapor saturation deficit (VPD), net radiation (Rn), precipitation (Rain), 5cm soil volumetric water content (SWC), and enhanced vegetation index (EVI) in the meadow steppe on the north shore of Qinghai Lake from 2009 to 2016 |
分析2009—2016年8年的年均温(Ta)得出(图 2), 8年平均温度为2 ℃, 2011年、2012年在这8年期间年均气温偏低, 分别为1.44 ℃和1.39 ℃, 2016年均温最高为2.58 ℃, 其余几年变幅不大。8年平均降水量(Rain)为439.5 mm, 2010年、2015年、2016年降水量偏低, 2010年降水量最少为371.6 mm, 2009年降水量最多为499.2 mm, 2011年、2012年、2013年、2014年降水量均超过440 mm。年均水汽饱和亏(VPD)8年平均值为0.39 kPa, 2014年VPD最低为0.36 kPa, 2013年VPD最高为0.44 kPa, 2010—2012年有持续降低趋势, 2014—2016年为持续升高趋势。8年年均净辐射(Rn)为94.6 W/m2, 2014年Rn最大为106.9 W/m2, 2009—2013年Rn稳定在90 W/m2左右, 2014—2016年Rn上升至100 W/m2以上。8年5 cm土壤含水量(SWC)平均值为0.17 cm3/cm3, 在0.15—0.19 cm3/cm3之间浮动, 2012年SWC最高, 2010年SWC最低, 除2010年、2014年SWC均高于平均水平, 其余年份较平均值都偏低。8年增强型植被指数(EVI)平均值为0.28, 2011年EVI最低为0.26, 2012年EVI最高为0.29, 2013—2016年EVI均在0.28以上。

|
| 图 2 青海湖北岸草甸草原2009—2016年气温(Ta)、降水量(Rain)、水汽饱和亏(VPD)、净辐射(Rn)、5 cm土壤含水量(SWC)、增强型植被指数(EVI)年际变化特征 Fig. 2 Interannual variation characteristics of air temperature (Ta), Precipitation (Rain), water vapor saturation deficit (VPD), net radiation (Rn), 5 cm soil moisture content (SWC) and enhanced vegetation index (EVI) in the meadow steppe on the north shore of Qinghai Lake from 2009 to 2016 |
从图 3分析得出:研究期逐8日NEE均值在-2.02—0.88 gC m-2 d-1之间, 平均值为-1.73 gC m-2 d-1, 日均NEE最小值出现在6月26日(-2.02 gC m-2 d-1), 最大值出现在11月2日(0.88 gC m-2 d-1), 在观测期内5—9月NEE为负值, 表现为碳吸收, NEE负值日数约占全年的48%, 10月—翌年4月为正值, 表现为碳释放;生态系统呼吸Re日均值为1.69 gC m-2 d-1, 受季节温度的影响, 呈现夏季强, 冬季弱的态势, 日均Re在0.29—4.91 gC m-2 d-1之间, Re从1月一直加强至7月, 随着气温的降低, 从8月起又呈下降趋势, 夏季(6—8月)Re均在4 gC m-2 d-1以上, 夏季的生态系统呼吸强度大约是冬季的8倍左右, 秋季较春季呼吸强度更大;总初级生产力GPP日均值为3.15 gC m-2 d-1, 最高值为7.3 0 gC m-2 d-1, 最低值为0.01 gC m-2 d-1, 随着光照辐射强度不断增大, GPP从冬季到夏季持续增大至峰值, 随后进入秋季非生长期, 植物光合作用减小, GPP又呈不断减小的态势, 整体上生长季的光合生产能力显著强于非生长期, 夏季最强, 秋季较春季更强。

|
| 图 3 青海湖北岸草甸草原2009—2016年净生态系统碳交换(NEE)、生态系统呼吸(Re)、总初级生产力(GPP)日均值变化特征 Fig. 3 Variation characteristics of daily mean values of net ecosystem carbon exchange capacity (NEE), ecosystem respiratory (Re) and total primary productivity (GPP) in the meadow steppe on the north shore of Qinghai Lake from 2009 to 2016 |
多年平均NEE月变化呈现明显的季节变化规律(图 4), 从春末5月起为CO2为吸收状态, 至盛夏7月达到吸收峰值, 8月起吸收强度又呈缓慢下降态势, 从秋季10月到次年4月后转变为CO2释放状态。8年平均来看5—9月NEE为负值, 表现为碳吸收, 平均CO2吸收强度为29.78 gC m-2 month-1, 雨热同期的6、7、8月是CO2净吸收最强的时期, 平均每月吸收CO2 39.85 gC m-2 month-1, 7月生态系统向大气吸收CO2能力最强, NEE值为-43.03 gC m-2 month-1。10月—翌年4月为正值, 表现为碳释放, 平均每月CO2释放强度为12.21 gC m-2 month-1, 初春3月和秋末11月是CO2净释放最强的时期, 分别释放CO2 19.17 gC m-2 month-1和18.07 gC m-2 month-1。多年平均Re月变化呈现明显的季节变化规律, 冬季生态系统呼吸强度最弱, 从春季开始生态系统呼吸逐渐加强, 盛夏生长季Re表现为最强, 然后到秋季又呈现逐渐减弱的态势。8年Re月平均在14.24—120.03 gC m-2 month-1之间, 生长盛期的6、7、8、9月是生态系统呼吸最强的时期, 月平均Re为105.05 gC m-2 month-1, 7月生态系统呼吸能力最强, Re值为120.03 gC m-2 month-1。冬季(12月—翌年2月)是生态系统呼吸最弱的时期, 月平均Re为15.83 gC m-2 month-1, 2月生态系统呼吸最弱, Re为14.24 gC m-2 month-1。多年平均GPP月变化呈现明显的季节变化规律, 冬季1、2月和初春3月GPP为0, 到4月逐渐增大至7月达到峰值, 从8月又呈下降态势到秋季, 直至秋末初冬GPP为0。4—10月平均月GPP为26.85、69.68、133.71、163.06、151.41、105.62、36.09 gC m-2 month-1, 其中7月GPP最大(163.06 gC m-2 month-1), 4月GPP最小(26.85 gC m-2 month-1), 4—9月平均每月GPP为98.06 gC m-2 month-1。

|
| 图 4 青海湖北岸草甸草原2009—2016年净生态系统碳交换(NEE)、生态系统呼吸(Re)、总初级生产力(GPP)月均值变化特征 Fig. 4 Monthly mean values of net ecosystem carbon exchange (NEE), ecosystem respiration(Re)and total primary productivity (GPP) in the meadow grassland on the north shore of Qinghai Lake from 2009 to 2016 |
从NEE年累积值分析可得, 2009—2016年8年NEE均为负值, 研究区碳收支表现为较强的碳汇功能。其中2015年碳吸收最强NEE为-95.80 gC m-2 a-1, 2016年碳吸收最弱NEE为-30.60 gC m-2 a-1, 8年平均碳吸收强度为63.51 gC m-2 a-1。年总生态系统呼吸Re在2010—2013年较弱, 2009年和2014—2016年较强, 平均呼吸强度为623.60 gC m-2 a-1, 8年呼吸强度范围在579.91—686.16 gC m-2 a-1之间, 2016年呼吸强度最大为686.16 gC m-2 a-1, 2013年呼吸强度最小为579.91 gC m-2 a-1, 2010—2013年Re均低于600 gC m-2 a-1。8年的年总初级生产力GPP变异性较高, 但没有明显的变化趋势, 变化范围在645.32—756.97 gC m-2 a-1, 8年平均值为687.10 gC m-2 a-1, 2015年GPP最大(756.97 gC m-2 a-1), 2011年最小(645.32 gC m-2 a-1)。

|
| 图 5 青海湖北岸草甸草原2009—2016年净生态系统碳交换(NEE)、生态系统呼吸(Re)、总初级生产力(GPP)年际变化特征 Fig. 5 Interannual variation characteristics of net ecosystem carbon exchange capacity (NEE), ecosystem respiratory (Re) and total primary productivity (GPP) in the meadow steppe on the north shore of Qinghai Lake from 2009 to 2016 |
运用SEM分析逐8日环境因子对NEE、GPP和Re的贡献率和调控机制, 通过2009—2016年8年的数据分析可得:Ta对Re的有显著提高的作用, 相对贡献量为1.10(R2=0.95), 而VPD通过Re对NEE有极显著间接负反馈, 相对贡献量为-0.39。EVI、Rain和e有提高Re的作用, 但不显著且贡献量较小, 路径系数分别为0.16、0.05、0.03。Ta显著提高GPP, 相对贡献量为0.74, Rain和Ts与GPP正相关, 但不显著, 相对贡献量都为0.06。EVI与GPP有极显著正相关, EVI通过GPP对NEE有间接极显著负反馈作用, EVI与Re正相关, 但没有像GPP和NEE那么显著。GPP和Re对NEE有极显著影响, GPP对NEE有负响应, 相对贡献量为-2.65, Re对NEE有显著提高作用, 相对贡献量为1.76, GPP和Re相互有极显著正相关。

|
| 图 6 青海湖北岸草甸草原生态系统2009—2016年环境因子和CO2通量(NEE、GPP、Re)的结构方程模型图 Fig. 6 The structural equation model of CO2 flux (NEE, GPP, Re) induced by environmental factors in the meadow grassland ecosystem of the north shore of Qinghai Lake from 2009 to 2016 Ta: 气温; Ts: 5cm土壤温度; PPFD: 光合有效辐射; VPD: 水汽饱和亏; Rain: 降水量; EVI: 增强型植被指数; Ws: 风速; e: 水汽压; GPP:总初级生产力;NEE: 净生态系统CO2交换量; Re: 生态系统呼吸黑色实线箭头是正相关, 黑色虚线箭头是负相关, 箭头上的系数代表通径系数, 箭头粗细代表通径系数的大小, ***、**代表极显著, *代表显著, ns代表不显著 |
青海湖北岸草甸草原生态系统在2009—2016年8年均表现为碳汇, 高海拔、高纬度地区由于气温低, 较高的植被层根茎比, 并且地下死根和凋零物由于环境温度低而不容易分解, 生态系统积累的有机碳可以较长时间地储存于地下根系和土壤中。因此, 有大量研究表明高纬度和高海拔的天然草地生态系统可能是全球重要的碳汇[17]。研究区8年平均NEE为-63.51 gC m-2 a-1, 比2009、2010年贝加尔针矛草原年NEE为-181.67 gC m-2 a-1、-66.38 gC m-2 a-1碳净吸收能力稍弱[18], 但年均碳净吸收能力比NEE为-39.59 gC m-2 a-1长白山高山冻原强, 美洲北方大草原在无放牧条件下年NEE为-34.13—-57.06 gC m-2 a-1之间, 在放牧状态下, 表现为碳释放, NEE为51.6 gC m-2 a-1[19], 其播种草原也表现为碳释放, NEE为34.94 gC m-2 a-1, 而在半干旱草地生态系统中, 碳释放更强, 科尔沁沙质草地为109.79 gC m-2 a-1(2015年)、72.99 gC m-2 a-1(2016年)[20], 温带半干旱草原为131.59 gC m-2 a-1[21], 综上所述研究区高寒草甸草原生态系统具有较强的碳汇功能。研究区昼夜温差大, 有利于植被碳同化作用, 再加之海拔较高气温偏低, 尤其在夜间和非生长季的低温有效抑制了土壤微生物和植物的呼吸作用[22];生长季期间水热条件较好, 促进生态系统高的碳吸收量, 上述原因共同导致了青海湖北岸高寒草甸是一个较大的碳汇区。2015年碳吸收最强NEE为-95.80 gC m-2 a-1, 2016年碳吸收最弱NEE为-30.60 gC m-2 a-1, 相邻年份NEE差值达65 gC m-2 a-1, 说明生态系统碳通量受生物及非生物等诸多因子的影响, 即使是同一个生态系统不同年际间碳收支也存在很大的波动。如三江源退化高寒草甸在相邻年际间分别表现为弱碳源和弱碳汇[23];吉林通榆退化草原也出现类似的现象, 原因是该生态系统出现碳释放的年份降水异常偏少, 日均净碳交换速率较低, 导致生长季净碳吸收较弱[24];还有内蒙古典型草原在湿润年呈弱碳汇, 而在干旱年则转变为碳源[25]。因为年降水格局变异大, 植被较稀疏[26], 草原上土壤的持水性相对较差, 湿度较小和风速较大等多方面因素导致蒸发较强, 土壤水分有效性较低, 因此半干旱高寒草甸以及内蒙古[27]、欧洲等地[28]年际NEE在“碳源”和“碳汇”之间转换。
3.2 CO2源汇收支的调控原因研究区Rn最大值一般出现在6月下旬至7月, 随降水量增加或太阳高度角降低而逐渐下降, 在12月底或1月初下降至最小值, 这与西藏高原草原生态系统的Rn变化趋势一致[29]。太阳辐射差异受到地形和下垫面的影响, 白天在边界层造成显著的大气湍流, 有学者分析2014年科科斯卡冰川的气候变化中分析得出白天大气处于不稳定分层状态, 湍流强烈, 而夜间近地面大气湍流相对较弱[30]。土壤温度升高直接影响植物和土壤的呼吸作用[31], 本研究区中Ta与Re为正相关关系, 与王海波等人研究得出青藏高原高寒草甸生态系统呼吸与温度具有较好的指数相关关系, 土壤呼吸随温度升高而增加结果一致[32]。水分因子对于提高草地生产力有重要作用[33]。研究区在8年期间2012年SWC值最大, 净碳吸收也最强, 说明土壤含水量较大对碳吸收有显著影响作用, 这与黄河小浪底的人工林碳通量的年际变异与SWC和碳吸收显著相关结论一致[34], 当旱季时期生态系统有水分胁迫时, 其呼吸就土壤含水量的变化显得更为敏感[35], 由于土壤水分含量低、温度高会导致植物气孔关闭以及气孔导度和比叶面积下降[36], 进而抑制生态系统对水分的有效利用[37], 最终引起生态系统CO2净交换降低。NEE除了与高SWC相关性较高之外, 与低VPD也显著相关, 唐详等[38]在北京八达岭林场的人工林观测也证实了这一点, 这与本研究结果一致。这可能是因为这一阶段植物的叶面积较大, 对空气中的湿度更为敏感[39], 而土壤含水量较低时, 根系可以从较深的土壤中吸水而不影响碳吸收[40]。本研究中VPD对Re有显著影响, VPD越高Re越小, 可能是由于VPD越高, 植物叶片气孔导度越大, 植物的CO2交换越活跃[41], 植物呼吸作用相对较弱有关。从结构方程模型中可分析出GPP与Re相互影响较显著, 对于GPP和Re季节动态的相关性在诸多研究中都有发现[42—43], GPP是进行呼吸作用的主要供应基质, 必然会影响Re的季节和年际变化。已有研究表明气候变暖, 会延长植被的生长季, 而“温室效应”也会造成高寒植被提前进入生长季[44], 均暗示气候变暖有利于增强高寒草甸草原的碳汇能力。本文分析得出Ta对Re和GPP的有显著提高的作用, GPP和Re对NEE有极显著影响, GPP对NEE有负响应, GPP越大, NEE值越小, 碳吸收能力越强, 并且Re对NEE有显著提高作用。因此在气候变暖的背景下, 未来气温升高, 青海湖北岸草甸草原生态系统碳汇功能可能会加强, 与大量相关研究结论一致。
4 结论(1) 研究区5—9月NEE为负值, 表现为碳吸收, 6、7、8月是CO2净吸收最强的时期, NEE负值日数约占全年的48%, 10月—翌年4月为正值, 表现为碳释放, 初春3月和秋末11月是CO2净释放最强的时期;Re呈夏季强, 冬季弱的态势, 夏季的生态系统呼吸强度大约是冬季的8倍, 秋季较春季呼吸强度更大;GPP从冬季到夏季持续增大至峰值, 随后进入秋季非生长期, 植物光合作用减小, GPP又呈不断减小的态势。8年监测可得研究区为一碳汇区, 8年平均NEE为-63.51 gC m-2 a-1, 2015年碳吸收最强NEE为-95.80 gC m-2 a-1。
(2) Ta对Re和GPP均有显著提高的作用, GPP和Re对NEE有极显著影响, GPP对NEE有负响应, 而Re对NEE有显著提高作用。暗示在气候变暖的背景下, 未来气温升高, 青海湖北岸草甸草原生态系统碳汇功能可能会加强。
| [1] |
Marcott S A, Shakun J D, Clark P U, Mix A C. A reconstruction of regional and global temperature for the past 11, 300 years. Science, 2013, 339(6124): 1198-1201. DOI:10.1126/science.1228026 |
| [2] |
Zou F L, Li H D, Hu Q W. Responses of vegetation greening and land surface temperature variations to global warming on the Qinghai-Tibetan Plateau, 2001-2016. Ecological Indicators, 2020, 119: 106867. DOI:10.1016/j.ecolind.2020.106867 |
| [3] |
Friedlingstein P, Jones M W, O'Sullivan M, Andrew R M, Zaehle S. Global Carbon Budget 2019. Earth System Science Data, 2019, 11(4): 1783-1838. DOI:10.5194/essd-11-1783-2019 |
| [4] |
杨娟, 周广胜, 王云龙, 王玉辉. 内蒙古克氏针茅草原生态系统-大气通量交换特征. 应用生态学报, 2008, 19(3): 533-538. DOI:10.13287/j.1001-9332.2008.0147 |
| [5] |
Barcza Z, Haszpra L, Kondo H, Saigusa N, Yamamoto S, Bartholy J. Carbon exchange of grass in Hungary. Tellus B, 2003, 55(2): 187-196. DOI:10.3402/tellusb.v55i2.16695 |
| [6] |
Hurtt G C, Thomas R Q, Fisk J P, Dubayah R O, Sheldon S L. The impact of fine-scale disturbances on the predictability of vegetation dynamics and carbon flux. PLoS One, 2016, 11(4): e0152883. DOI:10.1371/journal.pone.0152883 |
| [7] |
Chai X, Shi P L, Zong N, Niu B, He Y T, Zhang X Z. Biophysical regulation of carbon flux in different rainfall regime in a northern Tibetan alpine meadow. Journal of Resources and Ecology, 2017, 8(1): 30-41. DOI:10.5814/j.issn.1674-764x.2017.01.005 |
| [8] |
周兴民, 王质彬, 杜庆. 青海植被. 西宁: 青海人民出版社, 1986.
|
| [9] |
李东, 曹广民, 胡启武, 吴琴, 李月梅, 王迎红. 高寒灌丛草甸生态系统CO2释放的初步研究. 草地学报, 2005, 13(2): 144-148. |
| [10] |
师生波, 贲桂英, 韩发, 李英年, 沈振西. 青藏高原地区矮嵩草草甸植物群落生长分析. 生态学报, 2001, 21(6): 871-876. |
| [11] |
Cao S K, Cao G C, Chen K L, Han G Z, Liu Y, Yang Y F, Li X D. Characteristics of CO2, water vapor, and energy exchanges at a headwater wetland ecosystem of the Qinghai Lake. Canadian Journal of Soil Science, 2019, 99(3): 227-243. DOI:10.1139/cjss-2018-0104 |
| [12] |
李春, 何洪林, 刘敏, 苏文, 伏玉玲, 张雷明, 温学发, 于贵瑞. ChinaFLUX CO2通量数据处理系统与应用. 地球信息科学, 2008, 10(5): 557-565. DOI:10.3969/j.issn.1560-8999.2008.05.002 |
| [13] |
Webb E K, Pearman G I, Leuning R. Correction of flux measurements for density effects due to heat and water vapour transfer. Quarterly Journal of the Royal Meteorological Society, 1980, 106(447): 85-100. DOI:10.1002/qj.49710644707 |
| [14] |
Kato T, Tang Y H, Gu S, Hirota M, Du M Y, Li Y N, Zhao X Q. Temperature and biomass influences on interannual changes in CO2 exchange in an alpine meadow on the Qinghai-Tibetan Plateau. Global Change Biology, 2006, 12(7): 1285-1298. DOI:10.1111/j.1365-2486.2006.01153.x |
| [15] |
Yu G R, Ren W, Chen Z, Zhang L M, Wang Q F, Wen X F, He N P, Zhang L, Fang H J, Zhu X J, Gao Y, Sun X M. Construction and progress of Chinese terrestrial ecosystem carbon, nitrogen and water fluxes coordinated observation. Journal of Geographical Sciences, 2016, 26(7): 803-826. DOI:10.1007/s11442-016-1300-5 |
| [16] |
于贵瑞, 孙晓敏. 陆地生态系统通量观测的原理与方法. 北京: 高等教育出版社, 2006.
|
| [17] |
Kato T, Tang Y H, Gu S, Cui X Y, Hirota M, Du M Y, Li Y N, Zhao X Q, Oikawa T. Carbon dioxide exchange between the atmosphere and an alpine meadow ecosystem on the Qinghai-Tibetan Plateau, China. Agricultural and Forest Meteorology, 2004, 124(1/2): 121-134. |
| [18] |
张盼弟. 温带草甸草原碳循环组分分析研究[D]. 北京: 中国农业科学院, 2014.
|
| [19] |
牛亚毅, 李玉强, 王旭洋, 龚相文, 罗永清, 田德宇. 干旱年份沙质草地生态系统净CO2通量年变化特征. 草业学报, 2018, 27(1): 215-221. |
| [20] |
Frank A B. Carbon dioxide fluxes over a grazed prairie and seeded pasture in the Northern Great Plains. Environmental Pollution, 2002, 116(3): 397-403. DOI:10.1016/S0269-7491(01)00216-0 |
| [21] |
Balogh J, Czóbel S, Juhász A, Fóti S, Tuba Z. Seasonal carbon-balance of a semi-desert temperate grassland ecosystem over a year period. Acta Biologica Szegediensis, 2002, 46(3): 221-222. |
| [22] |
Chang S C, Tseng K H, Hsia Y J, Wang C P, Wu J T. Soil respiration in a subtropical montane cloud forest in Taiwan. Agricultural and Forest Meteorology, 2008, 148(5): 788-798. DOI:10.1016/j.agrformet.2008.01.003 |
| [23] |
王斌. 三江源区退化和人工草地生态系统CO2通量及其影响机制的研究[D]. 天津: 南开大学, 2014.
|
| [24] |
杜群, 刘辉志, 冯健武, 王雷, 黄建平, 张武, Christian BERNHOFER. 半干旱区草原生态系统的碳交换特征. 中国科学: 地球科学, 2012, 42(5): 711-722. |
| [25] |
Hao Y B, Wang Y F, Sun X M, Huang X Z, Cui X Y, Niu H S, Zhang Y H, Yu G R. Seasonal variation in carbon exchange and its ecological analysis over Leymus chinensis steppe in Inner Mongolia. Science in China Serirs D: Earth Sciences, 2006, 49(2): 186-195. |
| [26] |
Ma W L, Shi P L, Li W H, He Y T, Zhang X Z, Shen Z X, Chai S Y. Changes in individual plant traits and biomass allocation in alpine meadow with elevation variation on the Qinghai-Tibetan Plateau. Science China Life Sciences, 2010, 53(9): 1142-1151. DOI:10.1007/s11427-010-4054-9 |
| [27] |
Liu R, Pan L P, Jenerette G D, Wang Q X, Cieraad E, Li Y. High efficiency in water use and carbon gain in a wet year for a desert halophyte community. Agricultural and Forest Meteorology, 2012, 162/163: 127-135. DOI:10.1016/j.agrformet.2012.04.015 |
| [28] |
Gilmanov T G, Soussana J F, Aires L, Allard V, Ammann C, Balzarolo M, Barcza Z, Bernhofer C, Campbell C L, Cernusca A, Cescatti A, Clifton-Brown J, Dirks B O M, Dore S, Eugster W, Fuhrer J, Gimeno C, Gruenwald T, Wohlfahrt G. Partitioning European grassland net ecosystem CO2 exchange into gross primary productivity and ecosystem respiration using light response function analysis. Agriculture, Ecosystems & Environment, 2007, 121(1/2): 93-120. |
| [29] |
石培礼, 孙晓敏, 徐玲玲, 张宪洲, 何永涛, 张东秋, 于贵瑞. 西藏高原草原化嵩草草甸生态系统CO2净交换及其影响因子. 中国科学: D辑, 2006(S1): 194-203. |
| [30] |
Wang M, Chen H, Wu N, Peng C H, Zhu Q A, Zhu D, Yang G, Wu J H, He Y X, Gao Y H, Tian J Q, Zhao X Q. Carbon dynamics of peatlands in China during the Holocene. Quaternary Science Reviews, 2014, 99: 34-41. DOI:10.1016/j.quascirev.2014.06.004 |
| [31] |
Kang X M, Hao Y B, Cui X Y, Chen H, Huang S X, Du Y G, Li W, Kardol P, Xiao X M, Cui L J. Variability and changes in climate, phenology, and gross primary production of an alpine wetland ecosystem. Remote Sensing, 2016, 8(5): 391. DOI:10.3390/rs8050391 |
| [32] |
王海波, 马明国, 王旭峰, 谭俊磊, 耿丽英, 于文凭, 家淑珍. 青藏高原东缘高寒草甸生态系统碳通量变化特征及其影响因素. 干旱区资源与环境, 2014, 28(6): 50-56. |
| [33] |
潘颜霞, 王新平, 苏延桂, 李小军, 高艳红. 荒漠人工固沙植被区浅层土壤水分动态的时间稳定性特征. 中国沙漠, 2009, 29(1): 81-86. |
| [34] |
Tong X J, Meng P, Zhang J S, Li J, Zheng N, Huang H. Ecosystem carbon exchange over a warm-temperate mixed plantation in the lithoid hilly area of the North China. Atmospheric Environment, 2012, 49: 257-267. DOI:10.1016/j.atmosenv.2011.11.049 |
| [35] |
Niu S L, Wu M Y, Han Y, Xia J Y, Li L H, Wan S Q. Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe. New Phytologist, 2008, 177(1): 209-219. DOI:10.1111/j.1469-8137.2007.02237.x |
| [36] |
李子涵, 罗维均, 杜虎, 宋同清, 彭海军, 王彦伟, 王世杰. 喀斯特常绿落叶阔叶混交林旱季CO2通量特征及其影响因子. 地球与环境, 2020, 48(5): 525-536. |
| [37] |
孙思思, 吴战平, 肖启涛, 于飞, 古书鸿, 方荻, 李浪, 赵兴炳. 云贵高原草地生态系统CO2通量变化特征. 草业学报, 2020, 29(4): 184-191. |
| [38] |
唐祥, 陈文婧, 李春义, 查天山, 吴斌, 王小平, 贾昕. 北京八达岭林场人工林净碳交换及其环境影响因子. 应用生态学报, 2013, 24(11): 3057-3064. |
| [39] |
Yang J, Zhou G S, Wang Y L, Wang Y H. Characteristics of net ecosystem flux exchanges over Stipa krylovii steppe in Inner Mongolia. Chinese Journal of Applied Ecology, 2008, 19(3): 533-538. |
| [40] |
牛晓栋, 孙鹏森, 刘晓静, 栾军伟, 刘世荣. 中国亚热带-暖温带过渡区锐齿栎林净生态系统碳交换特征. 生态学报, 2020, 40(17): 5980-5991. |
| [41] |
姜海梅, 张德广, 王若静, 何旭升, 曹乐, 王成刚. 不同生态系统呼吸模型在半干旱草原生长季碳循环研究中的比较及应用. 北京大学学报: 自然科学版, 2018, 54(3): 593-604. |
| [42] |
Lasslop G, Reichstein M, Papale D, Richardson A D, Arneth A, Barr A, Stoy P, Wohlfahrt G. Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: critical issues and global evaluation. Global Change Biology, 2010, 16(1): 187-208. DOI:10.1111/j.1365-2486.2009.02041.x |
| [43] |
Wang Y Y, Yao J M, Han H D, Liu S Y. Analysis of aerodynamic roughness of the debris-covered Keqicar Glacier. Plateau Meteorology, 2014, 33(3): 762-768. |
| [44] |
Ganjurjav H, Gao Q Z, Zhang W N, Liang Y, Li Y W, Cao X J, Wan Y F, Li Y, Danjiu L B. Effects of warming on CO2 fluxes in an alpine meadow ecosystem on the central Qinghai-Tibetan Plateau. PLoS One, 2015, 10(7): e0132044. |