 2023, Vol. 43
2023, Vol. 43文章信息
- 苏文华, 司宏敏, 张辉红, 张光飞, 周睿, 郭晓荣, 杨琼
- SU Wenhua, SI Hongmin, ZHANG Huihong, ZHANG Guangfei, ZHOU Rui, GUO Xiaorong, YANG Qiong
- 中国西南四种常见松树火适应性状比较
- Fire-adapted traits of four pine trees in the southwestern China
- 生态学报. 2023, 43(3): 1064-1072
- Acta Ecologica Sinica. 2023, 43(3): 1064-1072
- http://dx.doi.org/10.5846/stxb202101250256
-
文章历史
- 收稿日期: 2021-01-25





2. 云南大学生态与环境学院, 昆明 650091
2. School of Ecology and Environmental Science, Yunnan University, Kunming 650091, China
野火是陆地生态系统中一个常见且重要的生态因子, 影响着陆生植物的进化和陆地生态系统的格局与过程[1—3], 在植物性状形成、维持陆地生态系统的稳定与生物多样性方面具有重要意义[2, 4—6]。许多陆地生态系统与生境中的野火形成了特定的生态关系, 一方面野火发生是不可避免的;另一方面, 完全排除野火后可能会引发生态灾难[7—8]。人类只能学会与野火共存[8—10]。在人类活动加剧和全球气候变化背景下, 野火发生有加剧的趋势[7, 11]。野火与植物和生态系统的关系备受关注[9, 12]。
大部分松属物种生活在易发生野火的地区, 生境火状况从频繁发生林冠火或地表火, 到偶尔发生林冠火或地表火[13]。野火对松属的进化和分布有着重要的影响[14—16]。在与野火抗争中, 松属植物形成了一系列性状来提高适合度, 维持在易火生境中的生存与繁衍[12, 14]。主要适应性状有:球果延迟开放、厚树皮、针叶高可燃性、高温或烟雾促进种子萌发、草丛状幼苗期和快速高生长等[14, 17]。相关研究表明, 任何单一适应性状都不能使植物避免被烧毁, 而是以多个“火适应性状”组合在特定的火状况生境中提高松树的适合度[14]。因此, 具有某些火适应性状组合的物种占据特定的火状况生态位[18](简称火生态位)。如果生境火状况发生变化, 它们可能就失去适应意义[12]。
不同的火适应性状组合形成特殊的火适应生活史对策。根据Pausas[6]提出的理论, 松属植物存在火依赖、火耐受和火避免三类生活史对策。火耐受对策的松树适应频发地表火的生境, 一般具有较厚的基部树皮、下枝自疏等性状;火依赖对策的松属植物适应会发生林冠火的生境, 一般是具有球果延迟开放特征的物种, 适应对策的重点是火后快速更新恢复;火避免对策的松树生活在鲜少发生野火的生境中, 火适应性状不明显。绝大部分松属植物都可以被归入这三类适应对策中, 有些广泛分布的种会因生境火状况的变化在种群间表现出不同的适应对策[19—20]。对不同适应对策的物种而言, 气候变化引发的林火加剧带来的影响可能是不一样的。已有研究显示, 一旦灾难性的高强度野火在一些历史上以地表火为主的生态系统中出现, 会导致生境中一些不具备球果延迟开放的松树无法在火后成功更新, 引发森林类型的改变[21—22]。植物火适应对策的认知对林地火后管理十分重要。
中国是松属植物的分布中心之一, 有22种松树[23]。中国的大部分区域也是野火多发区[24], 但从生活史层面研究松属植物与野火生态关系的不多。中国西南地区的小部分区域全年受东南季风控制, 降水丰沛湿润, 森林火险等级低;而大部分区域冬季受西南季风控制, 少雨干旱, 春季森林火险等级高[25—26]。西南地区有3种特有种或变种松树, 云南松(Pinus yunnanensis var. yunnanensis)、思茅松(P. kesiya var. langbianensis)和高山松(P. densata)。我国东部广泛分布的马尾松(P. massoniana)在成都盆地和云贵高原东部的贵州也有少量分布。4个种空间分布不重叠, 但边界相连, 存在地理替代关系[27]。4个种中, 云南松具有典型的火适应性状, 球果延迟开放[28]。4个种的种子都表现出萌发前高温处理促进萌发的热冲击效应[29], 表明这四种松树的进化都受到了周期性过火的影响[12, 14]。但缺少系统的火适应, 以及火适应生活史对策方面的研究。
本文将通过对西南地区这四种松树的火适应相关性状以及分布区火险和生境火状况的比较, 探讨:(1)这四种松树火适应生活史对策的差异, (2)生境火状况差异在四种松树分布格局中的作用。研究结果有利于更深入地了解野火在我国南方松树进化以及松林生态系统形成中的作用, 探讨各自分布区自然野火发生规律及生态作用;为西南地区松林野火管理提供理论依据和科学指导。
1 研究方法 1.1 样地选择每种松树选7—10个典型分布地作为样点, 考虑马尾松的主要分布区范围, 一些样点选在西南地区以外。调查采样地中, 云南松10个样点分布在云南、贵州和四川, 思茅松8个样点均在云南, 高山松9个样点分布在云南、四川和西藏, 马尾松7个样点分布在四川、贵州、广西、湖南和安徽。
1.2 树皮厚度的调查和测量在树干距地面大约30 cm且外观正常处, 用树皮取样器在树干北面取样。每个样点分5个胸径级为:I级1—10 cm、II级11—20 cm、III级21—30 cm、IV级31—40 cm和V级>40 cm, 每个胸径级采至少3棵树, 总共15—30棵。用游标卡尺测量树皮的内、外皮层的厚度和总厚度[12]。相对树皮厚度等于总厚度除以木材直径, 木材直径为采样处直径减去2倍树皮厚度[30]。
1.3 球果延迟开放的调查每个样点选30棵树, 观察每一棵松树球果是否具有宿存、延迟开放特征(每棵树随机选三支由主干发出的枝条, 分别数出开放和闭合球果的数量)。这里“延迟开放度”定义为球果成熟后继续留在树上的闭合数量与该树现有的全部同龄球果间的比例[31]。

|
每个样点选30棵树, 每棵树选取1个球果。球果45℃烘至鳞片开裂释放出种子。每个球果选出30粒饱满且带完整种翅的种子, 按粒用精度为0.001 g的电子分析天平称量每颗种子的质量并记录。用扫描仪扫描种翅, 并用Image J软件计算出种翅面积。翅载力用单位种翅面积的种子总质量来表示[32]。翅载力高则种子下降快, 飞行距离短[32]。计算公式为:
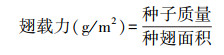
|
每个样点选30棵树, 每棵树剪一条小枝(能正常晒到太阳)。每枝取15束生长完整的针叶, 编号并带回。用直尺和测厚仪分别测量每束针叶中所有的针叶长度和直径, 取平均值作为该束针叶的长度和直径。然后计算针叶的长径比(针叶长度与针叶直径的比值)。
1.6 木质部密度的测量每个样点选15棵树, 用内径为5.15 mm的生长锥在树干距地面大约1.3 m处钻取树芯。树芯样品带回实验室后用刀片切去树皮, 并将两端切平, 测量长度(cm)后置于60℃烘箱中烘干, 测量干重(g)。木质部密度计算公式为:
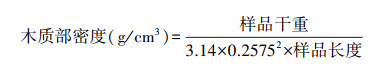
|
利用Excel软件对数据进行统计分析并作图, 用Origin 2019通过95%水平进行单因素方差分析(ANOVA), Duncan检验法进行多重比较及差异显著性检验(α=0.05)[33]。使用Origin 2019对火适应性状进行主成分分析和聚类分析, 聚类方法采用ward的方法、欧氏距离(Euclidean Distance)进行[34]。
2 结果与分析 2.1 四种松树火适应性状比较 2.1.1 树皮厚度的比较四种松树树皮厚度与胸径之间具有极显著的正相关关系(P < 0.001), 它们的回归系数分别为:云南松0.797, 高山松1.099, 思茅松0.596, 马尾松0.621(图 1)。高山松树皮厚度随胸径增长最快, 云南松次之, 思茅松最慢。云南松和思茅松的回归截距更高, 表明在幼树阶段, 云南松和思茅松的树皮比高山松和马尾松的更厚(图 1)。树皮厚度达到可抵抗地表过火的15 mm时[35], 个体胸径值分别为云南松8.1 cm, 高山松13.3 cm, 思茅松11.6 cm, 马尾松14.8 cm。

|
| 图 1 云南松、思茅松、高山松和马尾松基部树皮厚度与胸径的关系比较 Fig. 1 Relationship between basal bark thickness and diameter at breast height of four pines |
调查样地中, 云南松(37.35±33.90)%的植株有宿存1年以上的闭合球果, 高山松(2.06±2.63)%、思茅松(4.99±4.71)%、马尾松仅有(0.14±0.38)%(图 2)。云南松延迟开放度为(17.25±17.01)%, 显著大于思茅松(1.94±1.74)%、高山松(0.48±0.87)%和马尾松(0.14±0.38)%, 后三者之间差异不显著(P < 0.05)(图 2)。思茅松、高山松和马尾松在调查样地中只有极个别的个体存在延迟开放, 不是普遍现象。

|
| 图 2 云南松、思茅松、高山松和马尾松球果延迟开放程度和样点延迟开放树占比的比较 Fig. 2 Comparison of percentage of serotinous cones and percentage of serotinous individuals among four pines 不同字母标记表示物种间存在显著差异(P < 0.05) |
云南松木质部密度最大, 为(0.51±0.05)g/cm3。其次是思茅松和马尾松的木质部密度为(0.44±0.04)g/cm3和(0.45±0.04)g/cm3, 两者之间无显著差异(P>0.05)。高山松木质部密度最小, 为(0.37±0.03)g/cm3(图 3)。

|
| 图 3 云南松、思茅松、高山松和马尾松木质部密度的比较 Fig. 3 Comparison of density of xylems among four pines |
云南松针叶最长, 为(22.24±1.79)cm。其次是思茅松, 为(19.58±1.43)cm。马尾松的针叶长度为(16.87±1.02)cm, 显著大于高山松(12.99±1.36)cm(图 4)。高山松的针叶最粗, 直径为(0.89±0.06)mm。云南松次之, 为(0.71±0.06)mm。思茅松针叶直径为(0.61±0.04)mm;马尾松最细, 为(0.61±0.01)mm(图 4)。四种松树中, 高山松针叶短粗, 云南松细长。

|
| 图 4 云南松、思茅松、高山松和马尾松针叶长度和直径的比较 Fig. 4 Comparison of length and diameter of needles among four pines |
四种松树的种子质量均值分别为云南松0.0183 g、思茅松0.0192 g、高山松0.0151 g和马尾松0.0146 g。云南松和思茅松的种子质量无显著差异, 高于高山松和马尾松。高山松和马尾松之间无显著差异(P>0.05)(图 5)。云南松、思茅松和马尾松种翅面积无显著差异, 高山松的种翅面积显著小于其他三种松树(P < 0.05)(图 5)。马尾松的种子翅载力最低, 显著低于思茅松和高山松(P < 0.05), 云南松次之, 思茅松最高(图 5)。

|
| 图 5 云南松、思茅松、高山松和马尾松种子质量、种翅面积和翅载力比较 Fig. 5 Comparison of the weight of seeds, area of wings and seed wing loading among four pines |
基于13个与火适应相关性状的主成分分析显示, 四种松树位置有所分离(图 6)。两个主成分共能解释63%, 第一主成分贡献最大的性状有针叶长、球果长和种翅面积等, 第二主成分贡献大的有针叶直径和球果延迟开放度等。高山松明显与其它三个种分开, 最显著的区别是针叶较粗。云南松、思茅松与马尾松没有表现出明显分界, 尤其是云南松与思茅松, 有的种群很靠近。

|
| 图 6 云南松、思茅松、高山松和马尾松的13个火适应性状主成分分析 Fig. 6 Principal component analysis for 13 fire-adapted traits of four pines |
四种松树都具有一些典型的火适应性状, 它们的进化受到了野火选择压力的影响[36—37]。其中, 思茅松的原变种卡西亚松(P. kesiya)主要分布的中南半岛是亚洲野火发生频率最高的地区[11, 23]。思茅松生长在旱季更干燥的云南南部和中南半岛中北部, 生境野火发生频率也应该较高。该区域总体水热条件好, 思茅松每年春、秋两次抽梢, 年高生长可达0.91 m[38], 是4个种中最快的。林冠高度是在地表火中存活下来的最重要因素[14]。此外, 思茅松幼树树皮相对较厚, 大约16年生, 胸径约12 cm的植株树皮厚度可达到15 mm, 能有效抵抗地表火[35]。同时, 思茅松具有较长的松针, 过火时燃烧更强烈[39—40], 有助于消除竞争者[41]。思茅松是以快速高生长, 幼树厚树皮适应生境频繁的地表火。
云南松主要分布的滇中高原, 旱季更长, 发生野火风险更高[25]。同时热量和降水量下降, 云南松年平均高生长不足0.5 m[42], 30年的树高一般10—15 m。林地一旦起火, 容易形成林冠火。云南松进化出延迟开放的球果, 能在火后释放种子, 实现种群持续[43]。物种具备延迟开放被认为与生境经常发生林冠火有关, 且往往形成火依赖生活史对策[6]。云南松的球果延迟开放度不高[44], 低于地中海的P. halepensis [45], 表明云南松林发生林冠火的几率应该低于地中海地区。典型的火依赖对策松树往往不需要很强的抗火能力, 没有很厚的树皮[6]。云南松却具有厚树皮和较大的外皮占比, 表明生境地表火的危害较重[12]。本研究认为, 云南松林发生林冠火的风险不高, 但地表火应该较为频繁。云南松针叶长且粗, 提高了易燃性和过火烈度[6]。这是一些具有火后更新能力的物种的适应对策[46—48]。云南松是演替前期的树种, 较强的耐火能力可使其通过强烈的地表火压制、清除演替后期树种。
高山松分布于西藏高原东部边缘地带, 春季干燥易发生野火。四种松树中, 高山松木质部密度最低, 是快速生长对策。但生长季最短, 年高生长只有约0.4 m[49]。参考亚洲北方针叶林的自然野火发生周期[11], 高山松林的自然过火周期应该大于50年。较长的野火间期提供时间使其形成高大树干来躲避林冠火。有研究报道高山松表现出一定的抗火能力, 地表火还会释放其生长[50], 本研究也发现高山松有较厚的树皮。高山松分布区山高谷深, 地形破碎, 过火时常常形成地表火和林冠火镶嵌的混合火。火后残存的植株成为林冠火迹地的种子源, 高山松较轻的种子有利于传播更远[51—52]。与云南松不同[53], 高山松还具有土壤种子库[54—55], 可作为火后更新的种子源。在野外已观察到高山松火后种子更新。
马尾松是我国东部湿润区最常见的针叶树种, 分布区全年受东南季风的控制, 降水丰富, 湿润。生境过火风险要低于其他三种松树的生境[56]。马尾松的树皮较薄, 但胸径15 cm及以上的植株可以抵抗地表火[35]。这与贵州喀斯特地区马尾松林地过火, 平均胸径大于17 cm的林地个体死亡率小于10%的研究结果相符[57]。四种松树中, 马尾松针叶最细, 为快速燃烧对策[48]。提高针叶的燃烧速率, 减少过火时间, 有利于马尾松相对较薄的树皮抵抗火烧。四个种中, 马尾松的抗火能力表现最弱, 适应生境偶尔发生的林火。
3.2 四种松树生境火险与适应对策依据《全国森林火险区划等级》, 可将森林火险区划等级分为Ⅰ、Ⅱ、Ⅲ级, 分别表示森林火险高、中、低[56]。对本研究的样点以县级单位进行火险等级确认可得, 云南松、思茅松和高山松分布区森林火险为最高的Ⅰ级, 而马尾松森林火险为Ⅱ级[31]。根据鲁韦坤等[25]关于云南省森林火险气象等级区划, 云南松分布区的火险等级主要为5、4、3级, 思茅松分布区火险等级主要为3、2级, 高山松分布区火险等级为较低的2、1级。据此, 四个种分布区火险等级由高到底的顺序为, 云南松最高, 其次是思茅松、高山松, 马尾松最低。四种松树的火适应性状与其生境火险等级基本相符。
主成分分析的结果显示4种松树的火生态位空间总体上是分离的, 也表现出一些重叠(图 6), 反映出适应生境不同火状况的生态分化[3, 17]。Gallien等[58]利用8个质体基因建立了松树系统发育进化关系。其中马尾松很早与卡西亚松(思茅松原变种)和云南松分开, 云南松与高山松的亲缘关系较近。本研究用13个火适应性状进行聚类分析(图 7), 结果与Gallien的结果有些不同。尽管高山松与云南松和思茅松亲缘关系更近, 但火适应对策却有较大差异。亲缘关系较远的马尾松和思茅松表现出更相似的火适应对策。张辉红用球果松脂含量等9个性状聚类结果同样显示马尾松与思茅松有更相似的火适应策略[31]。在相似的生态环境中亲缘关系远的松属植物往往形成类似的生态适应性状[59], 表现出平行进化[36]。

|
| 图 7 四种松树火适应性状聚类图 Fig. 7 Cluster diagram obtained on fire-adapted traits of four pine trees |
根据以上结果, 本研究认为云南松火适应生活史对策是火耐受型和火依赖型的中间类型, 适应会发生林冠火的生境。高山松、思茅松和马尾松都属于火耐受型对策, 其中思茅松以快速高生长和幼树厚树皮适应频繁的地表火, 高山松以较厚的树皮适应低频率的地表火, 马尾松快速高生长适应生境偶尔发生的地表火。这四种松树所表现出的火适应性状说明野火在这一区域广泛存在, 并对植物进化和森林格局有重要影响。四种松树的地理替代现象是生境火状况和水热条件差异共同作用的结果。火在这一地区松属分布和种群维持中的生态作用应被给予足够重视。
| [1] |
Bond W J, Woodward F I, Midgley G F. The global distribution of ecosystems in a world without fire. New Phytologist, 2005, 165(2): 525-538. DOI:10.1111/j.1469-8137.2004.01252.x |
| [2] |
Bowman D M J S, Balch J K, Artaxo P, Bond W J, Carlson J M, Cochrane M A, D'Antonio C M, Defries R S, Doyle J C, Harrison S P, Johnston F H, Keeley J E, Krawchuk M A, Kull C A, Marston J B, Moritz M A, Prentice I C, Roos C I, Scott A C, Swetnam T W, van der Werf G R, Pyne S J. Fire in the earth system. Science, 2009, 324(5926): 481-484. DOI:10.1126/science.1163886 |
| [3] |
Pausas J G, Keeley J E. A burning story: the role of fire in the history of life. BioScience, 2009, 59(7): 593-601. DOI:10.1525/bio.2009.59.7.10 |
| [4] |
Pausas J G, Ribeiro E, Vallejo R. Post-fire regeneration variability of Pinus halepensis in the eastern Iberian Peninsula. Forest Ecology and Management, 2004, 203(1/2/3): 251-259. |
| [5] |
Keeley J E, Pausas J G, Rundel P W, Bond W J, Bradstock R A. Fire as an evolutionary pressure shaping plant traits. Trends in Plant Science, 2011, 16(8): 406-411. DOI:10.1016/j.tplants.2011.04.002 |
| [6] |
Pausas J G. Evolutionary fire ecology: lessons learned from pines. Trends in Plant Science, 2015, 20(5): 318-324. DOI:10.1016/j.tplants.2015.03.001 |
| [7] |
Gill A M, Stephens S L, Cary G J. The worldwide "wildfire" problem. Ecological Applications, 2013, 23(2): 438-454. DOI:10.1890/10-2213.1 |
| [8] |
Moritz M A, Batllori E, Bradstock R A, Gill A M, Handmer J, Hessburg P F, Leonard J, McCaffrey S, Odion D C, Schoennagel T, Syphard A D. Learning to coexist with wildfire. Nature, 2014, 515(7525): 58-66. DOI:10.1038/nature13946 |
| [9] |
Kelly L T, Brotons L. Using fire to promote biodiversity. Science, 2017, 355(6331): 1264-1265. DOI:10.1126/science.aam7672 |
| [10] |
North M P, Stephens S L, Collins B M, Agee J K, Aplet G, Franklin J F, Fulé P Z. Reform forest fire management. Science, 2015, 349(6254): 1280-1281. DOI:10.1126/science.aab2356 |
| [11] |
van der Werf G R, Randerson J T, Giglio L, van Leeuwen T T, Chen Y, Rogers B M, Mu M Q, van Marle M J E, Morton D C, Collatz G J, Yokelson R J, Kasibhatla P S. Global fire emissions estimates during 1997-2016. Earth System Science Data, 2017, 9(2): 697-720. DOI:10.5194/essd-9-697-2017 |
| [12] |
Pausas J G. Bark thickness and fire regime. Functional Ecology, 2015, 29(3): 315-327. DOI:10.1111/1365-2435.12372 |
| [13] |
Schwilk D W, Ackerly D D. Flammability and serotiny as strategies: correlated evolution in pines. Oikos, 2001, 94(2): 326-336. DOI:10.1034/j.1600-0706.2001.940213.x |
| [14] |
Keeley J E. Ecology and evolution of pine life histories. Annals of Forest Science, 2012, 69(4): 445-453. DOI:10.1007/s13595-012-0201-8 |
| [15] |
Tapias R, Climent J, Pardos J A, Gil L. Life histories of Mediterranean pines. Plant Ecology: Formerly Vegetatio, 2004, 171(1/2): 53-68. |
| [16] |
Ledig F T, Hom J L, Smouse P E. The evolution of the new jersey pine Plains. American Journal of Botany, 2013, 100(4): 778-791. DOI:10.3732/ajb.1200581 |
| [17] |
Bellingham P J, Sparrow A D. Resprouting as a life history strategy in woody plant communities. Oikos, 2000, 89(2): 409-416. DOI:10.1034/j.1600-0706.2000.890224.x |
| [18] |
Murphy B P, Bradstock R A, Boer M M, Carter J, Cary G J, Cochrane M A, Fensham R J, Russell-Smith J, Williamson G J, Bowman D M J S. Fire regimes of Australia: a pyrogeographic model system. Journal of Biogeography, 2013, 40(6): 1048-1058. DOI:10.1111/jbi.12065 |
| [19] |
Tapias R, Climent J, Pardos J A, Gil L. Life histories of Mediterranean pines. Plant Ecology: Formerly Vegetatio, 2004, 171(1/2): 53-68. |
| [20] |
Ledig F T, Hom J L, Smouse P E. The evolution of the new jersey pine Plains. American Journal of Botany, 2013, 100(4): 778-791. DOI:10.3732/ajb.1200581 |
| [21] |
Retana J, Maria Espelta J, Habrouk A, Luis OrdoÑEz J, de Solà-Morales F. Regeneration patterns of three Mediterranean pines and forest changes after a large wildfire in northeastern Spain. Écoscience, 2002, 9(1): 89-97. DOI:10.1080/11956860.2002.11682694 |
| [22] |
Kemp K B, Higuera P E, Morgan P. Fire legacies impact conifer regeneration across environmental gradients in the US northern Rockies. Landscape Ecology, 2016, 31(3): 619-636. DOI:10.1007/s10980-015-0268-3 |
| [23] |
中国科学院中国植物志编辑委员会. 中国植物志-第七卷. 北京: 科学出版社, 1978.
|
| [24] |
田晓瑞, 舒立福, 赵凤君, 王明玉. 中国主要生态地理区的林火动态特征分析. 林业科学, 2015, 51(9): 71-77. |
| [25] |
鲁韦坤, 李湘, 余凌翔, 戴丛蕊, 徐虹. 云南省森林火险气象等级区划研究. 大气科学学报, 2011, 34(3): 322-328. DOI:10.3969/j.issn.1674-7097.2011.03.008 |
| [26] |
田晓瑞, 舒立福, 赵凤君, 王明玉. 未来情景下西南地区森林火险变化. 林业科学, 2012, 48(1): 121-125, 192. |
| [27] |
吴兆录. 云南松属植物地理分布和叶形变化. 云南林业科技, 1990, 19(4): 19-22. |
| [28] |
苏文华, 崔凤涛, 赵元蛟, 周睿, 张光飞, 曹建新. 云南松球果延迟开放及其植冠种子库. 生态学报, 2017, 37(2): 541-548. |
| [29] |
张辉红, 孟丽媛, 余婷, 余娇娥, 司洪敏, 吴雪涛, 马筱, 苏文华, 张光飞. 短时高温处理对4种松树种子萌发的影响. 种子, 2018, 37(1): 32-37. |
| [30] |
Midgley J J, Lawes M J. Relative bark thickness: towards standardised measurement and analysis. Plant Ecology, 2016, 217(6): 677-681. DOI:10.1007/s11258-016-0587-8 |
| [31] |
张辉红. 中国西南地区四种松树火适应性状与生态适应意义[D]. 昆明: 云南大学, 2018.
|
| [32] |
Matlack G R. Diaspore size, shape, and fall behavior in wind-dispersed plant species. American Journal of Botany, 1987, 74(8): 1150-1160. DOI:10.1002/j.1537-2197.1987.tb08729.x |
| [33] |
Duncan D B. Multiple range and multiple F tests. Biometrics, 1955, 11(1): 1. DOI:10.2307/3001478 |
| [34] |
Hartigan J A, Wong M A. Algorithm AS 136: a K-means clustering algorithm. Applied Statistics, 1979, 28(1): 100. DOI:10.2307/2346830 |
| [35] |
He T H, Pausas J G, Belcher C M, Schwilk D W, Lamont B B. Fire-adapted traits of Pinus arose in the fiery Cretaceous. New Phytologist, 2012, 194(3): 751-759. DOI:10.1111/j.1469-8137.2012.04079.x |
| [36] |
Badik K J, Jahner J P, Wilson J S. A biogeographic perspective on the evolution of fire syndromes in pine trees (Pinus: Pinaceae). Royal Society Open Science, 2018, 5(3): 172412. DOI:10.1098/rsos.172412 |
| [37] |
Lamont B B, He T H. Fire-proneness as a prerequisite for the evolution of fire-adapted traits. Trends in Plant Science, 2017, 22(4): 278-288. DOI:10.1016/j.tplants.2016.11.004 |
| [38] |
杨利华, 徐玉梅, 杨德军, 邱琼, 袁莲珍, 陈勇, 钟萍. 不同造林密度对思茅松中龄林生长量的影响. 江苏林业科技, 2013, 40(6): 43-46. DOI:10.3969/j.issn.1001-7380.2013.06.011 |
| [39] |
Fonda R, Bélanger L, Burley L. Burning characteristics of western conifer needles. Northwest Science, 1998, 72: 1-3. |
| [40] |
王秋华. 森林火灾燃烧过程中的火行为研究[D]. 北京: 中国林业科学研究院, 2010.
|
| [41] |
Williamson G B, Black E M. High temperature of forest fires under pines as a selective advantage over oak. Nature, 1981, 293(5834): 643-644. DOI:10.1038/293643a0 |
| [42] |
杨晏平, 周娟, 赵江萍, 李碧林, 李归林. 滇西地区湿地松人工林生长过程分析. 西部林业科学, 2020, 49(1): 128-132. |
| [43] |
司宏敏, 余娇娥, 吴雪涛, 马筱, 曹建新, 张光飞, 苏文华. 两种过火类型对云南松林损伤及火后自然更新的研究. 西部林业科学, 2019, 48(1): 7-11, 17. |
| [44] |
Su W H, Shi Z, Zhou R, Zhao Y J, Zhang G F. The role of fire in the Central Yunnan Plateau ecosystem, southwestern China. Forest Ecology and Management, 2015, 356: 22-30. DOI:10.1016/j.foreco.2015.05.015 |
| [45] |
Tapias R, Gil L, Fuentes-Utrilla P, Pardos J A. Canopy seed banks in Mediterranean pines of south-eastern Spain: a comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L. Journal of Ecology, 2001, 89(4): 629-638. DOI:10.1046/j.1365-2745.2001.00575.x |
| [46] |
Bond W J, Midgley J J. Kill thy neighbour: an individualistic argument for the evolution of flammability. Oikos, 1995, 73(1): 79. DOI:10.2307/3545728 |
| [47] |
Pausas J G, Moreira B. Flammability as a biological concept. The New Phytologist, 2012, 194(3): 610-613. DOI:10.1111/j.1469-8137.2012.04132.x |
| [48] |
Pausas J G, Keeley J E, Schwilk D W. Flammability as an ecological and evolutionary driver. Journal of Ecology, 2017, 105(2): 289-297. DOI:10.1111/1365-2745.12691 |
| [49] |
张昆林, 钟蓉军. 地形条件对高山松天然更新和生长的影响. 林业科技, 2000, 25(1): 22-23. |
| [50] |
李宝, 程雪寒, 吕利新. 西藏朗县地区不同龄级高山松林木径向生长对火干扰的响应. 植物生态学报, 2016, 40(5): 436-446. |
| [51] |
潘燕, 王帅, 王崇云, 朱晓媛, 任鹏, 胡明江, 艾培顺, 段啟纲. 云南松与云南油杉种子风力传播特征比较. 植物分类与资源学报, 2014, 36(3): 403-410. |
| [52] |
Andersen M C. Diaspore morphology and seed dispersal in several wind-dispersed Asteraceae. American Journal of Botany, 1993, 80(5): 487-492. |
| [53] |
Su W H, Yu J E, Zhang G F, Shi Z, Wang L L, Zhao G H, Zhou R. Comparison of the canopy and soil seed banks of Pinus yunnanensis in central Yunnan, China. Forest Ecology and Management, 2019, 437: 41-48. |
| [54] |
李照青, 高郯, 张铎, 卢杰. 西藏林芝高山松土壤种子库空间分布特征研究. 西北林学院学报, 2019, 34(4): 43-48. |
| [55] |
Lu J, Zhao Q L, Tang X Q, Xue J L. Characteristics of Soil Seed Bank and Natural Regenerations in Pinus densata Natural Forest of Southeast Tibet. Agricultural Science & Technology, 2017, 18(7): 1248-1254. |
| [56] |
王栋. 中国森林火险调查与区划. 北京: 中国林业出版社, 2000.
|
| [57] |
张喜, 崔迎春, 朱军, 潘德权, 杨春华, 陈骏, 吴鹏. 火烧对黔中喀斯特山地马尾松林分的影响. 生态学报, 2011, 31(21): 6442-6450. |
| [58] |
Gallien L, Saladin B, Boucher F C, Richardson D M, Zimmermann N E. Does the legacy of historical biogeography shape current invasiveness in pines?. New Phytologist, 2016, 209(3): 1096-1105. |
| [59] |
McCune B. Ecological diversity in North American pines. American Journal of Botany, 1988, 75(3): 353-368. |