 2023, Vol. 43
2023, Vol. 43文章信息
- 沈玉莹, 程俊翔, 徐力刚, 李仁英, 游海林, 杨海
- SHEN Yuying, CHENG Junxiang, XU Ligang, LI Renying, YOU Hailin, YANG Hai
- 极端水文干旱下鄱阳湖浮游动物群落结构特征及其影响因素
- Characteristics of zooplankton community structure in Poyang Lake and their influencing factors under extreme hydrological drought
- 生态学报. 2023, 43(24): 10399-10412
- Acta Ecologica Sinica. 2023, 43(24): 10399-10412
- http://dx.doi.org/10.20103/j.stxb.202306161291
-
文章历史
- 收稿日期: 2023-06-16
- 网络出版日期: 2023-11-10





2. 中国科学院南京地理与湖泊研究所, 南京 210008;
3. 中国科学院南京地理与湖泊研究所鄱阳湖湖泊湿地综合研究站, 庐山 332899;
4. 江西省生态文明研究院, 南昌 330036;
5. 中国科学院大学南京学院, 南京 211135;
6. 江西省科学院流域生态研究所, 南昌 330096;
7. 中国地质调查局南京地质调查中心, 南京 210016;
8. 自然资源部流域生态地质过程重点实验室, 南京 210016
2. Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China;
3. Poyang Lake Laboratory for Wetland Ecosystem Research, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Lushan 332899, China;
4. Jiangxi Research Academy of Ecological Civilization, Nanchang 330036, China;
5. University of Chinese Academy of Sciences, Nanjing, Nanjing 211135, China;
6. Institute of Watershed Ecology, Jiangxi Academy of Sciences, Nanchang 330096, China;
7. Nanjing Center, China Geological Survey, Nanjing 210016, China;
8. Key Laboratory of Watershed Eco-Geological Processes, Ministry of Natural Resources, Nanjing 210016, China
浮游动物作为水体食物链的基础构成部分, 是水生态系统中物质循环和能量流动的重要参与者[1]。浮游动物数量庞大、体积小、寿命短, 在营养级中有着承上启下的作用[2], 不仅通过摄食行为制约着浮游植物和微生物的生长繁殖[3], 而且是许多鱼类、虾类和水禽的重要食物来源[4]。浮游动物对环境变化比较敏感[5], 其群落结构可能随着水环境变化[6]、季节变化[7—8]而发生改变。浮游动物的自发变化能准确反映湖泊的状态[9], 因此常常被用作湖泊水生态状况的关键指示种之一。目前已有大量关于浮游动物群落结构及其影响因素的研究, 例如, Hu等[10]的研究表明鄱阳湖碟形湖浮游动物群落结构变化呈现典型的季节特征, 但不具有年际重复性, 且主要受到水位变化的影响;宋高飞等[11]通过对武汉市湖泊浮游动物进行调查研究, 认为营养盐、叶绿素a、高锰酸盐是影响浮游动物群落组成的关键因素;张建波等[12]采用典范对应分析揭示了洞庭湖后生浮游动物的分布主要受到叶绿素a、总氮、总磷和透明度的影响;巢湖浮游动物群落结构存在显著的季节和空间差异, 且水温和叶绿素a是影响枝角类丰度的重要环境因子[13]。以上研究表明湖泊浮游动物群落结构的时空异质性较大, 而且影响因素复杂多变。
气候变化可能会导致高温热浪和极端干旱的发生[14], 从而对湖泊水生态系统健康造成威胁。当湖泊面临极端干旱的威胁时, 最为明显的变化就是水位下降, 进而引起湖泊水体理化性质和浮游动物群落发生改变[15—16]。Ginatullina等[17]的研究发现干旱可以引起湖泊盐度的增加, 进而迫使浮游动物物种丰富度下降;Carrasco等[18]通过对St. Lucia河口浮游动物开展季度调查研究, 认为极端干旱使浮游动物类群的丰度和生物量均降低;然而, Primo等[19]对葡萄牙Mondego河口浮游动物的研究表明, 经历干旱后的浮游动物丰度增加, 且不再具有典型的季节差异。虽然极端干旱[20]和长期干旱[21]都会导致浮游动物群落在空间和时间上发生变化, 但是干旱对浮游动物的影响仍然具有较大的不确定性。此外, 以往研究主要关注干旱对河流浮游动物群落的影响, 亟需加强干旱胁迫下的湖泊浮游动物研究。
鄱阳湖作为中国的第一大淡水湖, 是保障长江中下游水生态安全的重要屏障[22]。2022年, 长江流域在汛期发生了非常罕见的高温干旱事件[23], 再加之降水偏少, 使得鄱阳湖面临近70年来最为严重的极端干旱[24]。极端干旱胁迫下的鄱阳湖主湖区和碟形湖严重干涸, 底栖动物大量死亡[25], 影响候鸟栖息地和食物来源[26]。为了深入探究极端干旱对鄱阳湖浮游动物群落的影响, 本研究基于2022年鄱阳湖全湖浮游动物及水环境的季度调查, 从浮游动物种类组成、丰度、生物量、多样性、优势种及其影响因素等方面进行了系统分析, 研究结果对极端干旱气候条件下的鄱阳湖水生态保护和恢复具有重要意义。
1 材料与方法 1.1 采样点布设鄱阳湖(28° 22′—29° 45′ N, 115° 47′—116° 45′ E)有着“洪水一片、枯水一线”的奇特景观, 承载着“五河”(赣江、抚河、信江、饶河、修水)来水, 经湖泊调蓄后由湖口汇入长江。浮游动物调查时间为2022年1月(冬季)、4月(春季)、7月(夏季)和10月(秋季), 将鄱阳湖分为三个湖区, 在北部湖区和南部湖区分别布设5个、中部湖区布设6个采样点, 共布设16个采样点。冬季和夏季采样点均为16个, 春季采样点为6个(PY1—PY6;受新冠疫情影响, 其余采样点无法到达), 秋季为除PY1、PY2、PY15以外的13个采样点(湖泊水位较低, 部分采样点无法到达)。其中, 采样点PY9、PY10分别对应赣江和修河入湖口, PY12、PY13和PY14分别为赣江南支入湖口、抚河入湖口和信江入湖口。星子站是鄱阳湖的代表性水文站, 本研究使用星子站水位表示湖泊水位。采样点及水文站的分布如图 1所示。

|
| 图 1 鄱阳湖浮游动物采样点及水文站分布图 Fig. 1 Distribution map of zooplankton sampling sites and hydrological station in Poyang Lake PY: 鄱阳湖 |
使用采水器进行定量采集, 当水深<2 m时, 在表层下0.5 m采集;水深为2—5 m时, 分别在表层下0.5 m、底层上0.5 m各采集一次;水深>5 m时, 则在表层下0.5 m处、中层以及底层上0.5 m处各采集一次。小型浮游动物采水样1 L, 直接装入样品瓶, 大型浮游动物采水样10—50 L, 再用25#浮游生物网过滤后装入样品瓶中, 并用5%的甲醛溶液对浮游动物进行固定。带回实验室后, 在解剖镜下全部计数鉴定。浮游动物的样品采集和分析参照《淡水浮游生物研究方法》[27]。
水温(WT)、pH、溶解氧(DO)、电导率(Cond)用多参数水质分析仪(YSI EXO2)进行现场测定, 透明度(SD)采用塞奇式圆盘法测定, 叶绿素a(Chl.a)采用热乙醇-分光光度法测定, 总氮(TN)采用过硫酸钾紫外分光光度法测定, 总磷(TP)采用连续流动-钼酸铵分光光度法测定, 化学需氧量(COD)采用酸性法测定, 硝态氮(NO3--N)采用紫外分光光度法测定, 氨氮(NH4+-N)采用水杨酸分光光度法测定。
1.3 生物多样性分析生物多样性分析采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J), 以及物种优势度指数(Y), Y>0.02视为优势种, 计算公式如下:
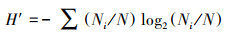
|
(1) |
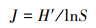
|
(2) |
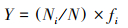
|
(3) |
式中, Ni为样品中某种生物的总个体数;N为样品中各生物的总个体数;S为所有样品中的种属总数;fi为某物种出现的频率。生物多样性指数水质评价标准[28]为:H′:0<H′<1为多污型, 1<H′<2为α—中污型, 2<H′<3为β—中污型, H′>3为清洁—寡污型;J:0<J<0.3为多污型, 0.3<J<0.5为中污型, 0.5<J<0.8寡污型, 0.8<J<1.0为清洁型。
1.4 统计分析水位、浮游动物群落结构、水体理化性质在不同季节的差异性采用单因素方差分析(One-way ANOVA), 浮游动物群落结构与环境因子的相关性分析采用Pearson相关性, 相应的分析在SPSS 26中完成, 并使用Origin 2018和ArcGIS 10.6进行制图。
对浮游动物的丰度数据进行除趋势对应分析(Detrended correspondence analysis, DCA), 在DCA结果中, 如果最大梯度长度>4, 选择典范对应分析(Canonical correspondence analysis, CCA)比较合适;如果最大梯度长度<3, 选择冗余分析(Redundancy analysis, RDA)更合适;如果介于3—4之间, 典范对应分析和冗余分析都是合适的[29], 使用R语言vegan包进行对应的分析。为提高数据的正态性, 分析之前对除pH外的数据进行log(x+1)转换。
2 结果 2.1 水位和水体理化性质 2.1.1 历史水位和2022年极端干旱水位鄱阳湖年内水位变化较大, 整体呈现先上升后下降的趋势, 并在夏季达到最高水位(图 2)。然而, 2022年的鄱阳湖平均水位仅11.38 m, 并在6月下旬出现水位下降的现象, 比历史平均水位(2000—2021年)开始下降时间提前23 d。由于2022年水位下降提前、丰水期水位过低、低枯水期延长, 造成了鄱阳湖极端水文干旱现象的出现。

|
| 图 2 2000—2022年鄱阳湖逐日和季节水位变化 Fig. 2 Daily and seasonal water level variations in Poyang Lake from 2000 to 2022 图中不同小写字母表示水位之间具有显著性差异(P<0.05) |
不同季节的鄱阳湖水位具有较大差异(图 2)。在夏季和秋季, 极端水文干旱年的水位显著低于历史平均水位, 尤其秋季水位与历史平均水位相比减少近40%, 表明2022年的鄱阳湖极端干旱主要发生在夏秋两季, 故夏秋两季被认为是干旱季节;春冬两季水位与历史平均水位相差较小, 为非干旱季节。在极端水文干旱年内夏季水位变化范围为13.40—18.26 m, 显著高于其他季节, 并出现秋季水位显著低于冬季的现象, 与多年历史平均水位情况不完全相同。
2.1.2 水体理化性质鄱阳湖不同季节的水体理化性质如表 1所示。各水环境因子都存在一定的季节性差异, SD、WT、Chl.a呈现先上升后下降的趋势, 在夏季达到最高值, SD低值出现在春、秋季, WT、Chl.a在冬季最低;DO、Cond、TN、TP、NO3--N、NH4+-N则先下降后上升, 最低值在夏季, TP最高值在秋季, 其他环境因子最高值均在冬季;COD在春、夏季显著高于秋、冬季, pH在春季显著低于其他季节。
| 指标Index | 春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter |
| SD/m | 0.30±0.06b | 0.43±0.16a | 0.30±0.07b | 0.34±0.06ab |
| WT/℃ | 16.00±0.89c | 32.45±0.97a | 19.40±0.61b | 10.89±1.64d |
| DO/(mg/L) | 12.23±1.24a | 7.11±0.73b | 7.77±0.44b | 12.66±2.57a |
| Cond/(mS/cm) | 0.13±0.05b | 0.10±0.01b | 0.19±0.03a | 0.17±0.03a |
| pH | 7.69±0.30b | 8.25±0.29a | 8.18±0.12a | 8.37±0.17a |
| COD/(mg/L) | 3.74±0.86a | 3.28±0.79a | 2.44±0.22b | 1.95±0.36b |
| Chl.a/(mg/m3) | 14.25±5.96ab | 22.53±12.78a | 13.88±3.79ab | 11.31±3.95b |
| TN/(mg/L) | 1.78±0.18c | 1.52±0.36c | 2.24±0.39b | 2.89±0.45a |
| TP/(mg/L) | 0.11±0.01b | 0.06±0.02b | 0.16±0.02a | 0.12±0.06ab |
| NO3--N/(mg/L) | 0.96±0.07c | 0.69±0.26d | 1.20±0.18b | 1.64±1.64a |
| NH4+-N/(mg/L) | 0.30±0.03b | 0.16±0.03c | 0.28±0.05b | 0.57±1.64a |
| 不同小写字母表示不同季节的理化因子之间具有显著性差异(P<0.05);SD:透明度Secchi depth;WT:水温Water temperature;DO:溶解氧Dissolved oxygen;Cond:电导率Conductivity;COD:化学需氧量Chemical oxygen demand;Chl.a:叶绿素a Chlorophyll a;TN:总氮Total nitrogen;TP:总磷Total phosphorus;NO3--N:硝态氮Nitrate nitrogen;NH4+-N:氨氮Ammonia nitrogen | ||||
本次调查共鉴定鄱阳湖浮游动物70种, 其中轮虫为优势种类, 有40种, 占浮游动物总物种数的57.14%;桡足类和枝角类分别为17种(24.29%)和13种(18.57%)。图 3表明浮游动物种类组成存在明显的季节和空间差异。从季节变化来看, 夏季浮游动物物种数为43种, 明显高于秋季(31种)、春季(26种)和冬季(24种);轮虫占夏季物种总数的65.12%, 枝角类和桡足类分别占夏季物种总数的18.61%和16.28%。从空间变化来看, 中部湖区的物种数最多(59种), 其次为南部湖区(55种), 最少的为北部湖区(46种)。轮虫的物种数在不同季节和不同湖区均占据主要优势。

|
| 图 3 不同季节和不同湖区的浮游动物物种数 Fig. 3 Number of zooplankton species in different seasons and lake zones |
极端水文干旱年的鄱阳湖浮游动物平均丰度为33.83个/L, 不同季节的浮游动物丰度组成与空间分布如图 4所示。在季节上, 夏季浮游动物丰度最高(72.21个/L), 其次是冬季(21.06个/L)和春季(20.17个/L), 秋季最低(8.63个/L);不同季节的浮游动物丰度具有极显著差异(P<0.01)。浮游动物丰度的优势种类随季节变化而变化:春季和夏季以轮虫为优势种类, 轮虫分别占春季和夏季总丰度的50.41%和62.72%;秋季以桡足类占比最大(64.51%);冬季以枝角类为优势种类(54.60%)。在空间上, 浮游动物丰度呈现由南向北减少的趋势, 南部湖区的浮游动物平均丰度最高(43.90个/L), 其次是中部湖区(34.06个/L), 北部湖区最低(24.12个/L), 浮游动物丰度不具有空间上的显著差异。

|
| 图 4 不同季节浮游动物丰度组成及空间分布 Fig. 4 Composition and spatial distribution of zooplankton abundance in different seasons |
极端水文干旱年的鄱阳湖浮游动物平均生物量为0.35 mg/L, 各季节的浮游动物生物量组成与空间分布如图 5所示。在季节上, 夏季浮游动物生物量最高(0.55 mg/L), 其次为冬季(0.41 mg/L)和春季(0.20 mg/L), 秋季最低(0.10 mg/L);除春季浮游动物生物量以枝角类为优势种类(37.65%), 夏季、秋季和冬季浮游动物生物量均为桡足类占主要优势, 分别占夏季、秋季和冬季总生物量的58.56%、49.81%和54.32%, 轮虫对各季节浮游动物生物量的贡献均很小;不同季节浮游动物生物量具有极显著差异(P<0.01)。在空间上, 浮游动物生物量呈现南部大于北部的特征, 南部湖区的浮游动物平均生物量最高(0.47 mg/L), 北部湖区次之(0.31 mg/L), 中部湖区最低(0.29 mg/L);桡足类生物量的空间变化与浮游动物生物量的空间变化特征基本一致;浮游动物生物量不具有空间上的显著差异性。

|
| 图 5 不同季节浮游动物生物量组成及空间分布 Fig. 5 Composition and spatial distribution of zooplankton biomass in different seasons |
极端水文干旱年的鄱阳湖浮游动物Shannon-Wiener指数(H′)和Pielou指数(J)均值分别为1.77±0.62和0.81±0.13, 其季节与空间变化特征如图 6所示。从季节变化来看, 夏季H′显著高于其他季节, 春季H′显著高于秋季和冬季;春季J显著高于秋季和冬季, 且秋冬两季的J变异较大。从空间变化来看, 三个湖区的H′和J均无显著差异。根据H′进行水质评价, 表明极端水文干旱年的鄱阳湖水体为α—中污型, 而根据J标准则为清洁型。

|
| 图 6 不同季节和不同湖区的浮游动物Shannon-Wiener指数和Pielou指数 Fig. 6 Shannon-Wiener index and Pielou index of zooplankton in different seasons and lake zones 不同小写字母表示不同季节、不同湖区的多样性指数之间具有显著性差异(P<0.05) |
鄱阳湖浮游动物优势种的组成在不同季节差异较大(表 2), 浮游动物优势种共16种, 其中轮虫9种、桡足类5种、枝角类2种。夏季的优势种最多, 春季次之, 秋冬两季最少;无节幼体在四个季节均为优势种。夏季优势种有10种, 以无节幼体、镰状臂尾轮虫为主要优势种, 两者丰度分别占夏季总丰度的15.58%和16.39%;春季主要优势种(7种)为简弧象鼻溞, 占春季总丰度的24.79%;秋季和冬季优势种较少, 优势度较大的分别为无节幼体和简弧象鼻溞。
| 优势物种 Dominant species |
代码 Code |
优势度Degree of dominance | |||
| 春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter | ||
| 长额象鼻溞Bosmina longirostris | sp1 | 0.027 | |||
| 简弧象鼻溞 Bosmina coregoni | sp2 | 0.248 | 0.115 | 0.424 | |
| 汤匙华哲水蚤Sinocalanus dorrii | sp3 | 0.030 | |||
| 球状许水蚤Schmackeria forbesi | sp4 | 0.026 | |||
| 无节幼体Nauplius | sp5 | 0.033 | 0.156 | 0.501 | 0.040 |
| 桡足幼体Cyclopoida larvae | sp6 | 0.031 | |||
| 广布中剑水蚤Mesocyclops leuckarti | sp7 | 0.029 | 0.061 | 0.056 | |
| 曲腿龟甲轮虫Keratella valga | sp8 | 0.045 | 0.039 | ||
| 镰状臂尾轮虫Brachionus falcatus | sp9 | 0.154 | |||
| 角突臂尾轮虫Brachionus angularis | sp10 | 0.095 | |||
| 剪形臂尾轮虫Brachionus forficula | sp11 | 0.049 | |||
| 裂足臂尾轮虫Brachionus diversicornis | sp12 | 0.032 | |||
| 等棘异尾轮虫Trichocerca similis | sp13 | 0.036 | |||
| 对棘异尾轮虫Trichocerca stylata | sp14 | 0.066 | |||
| 晶囊轮虫Asplanchna sp. | sp15 | 0.066 | |||
| 针簇多肢轮虫Polyarthra trigla | sp16 | 0.048 | |||
按照干旱季节和非干旱季节将鄱阳湖浮游动物物种数、丰度等群落结构特征与水环境和水位进行相关性分析, 结果表明浮游动物群落结构受水环境和水位的影响较大(图 7)。在干旱季节, 浮游动物物种数、丰度、生物量、Shannon-Wiener指数(H′)与WT、WL、Chl.a等呈显著正相关, 与NO3--N、NH4+-N、Cond等呈显著负相关;Pielou指数(J)与Cond呈显著负相关。在非干旱季节, 浮游动物物种数、Shannon-Wiener指数(H′)与WT、WL等呈显著正相关, Shannon-Wiener指数(H′)与NO3--N、NH4+-N等呈显著负相关;Pielou指数(J)与TN、NO3--N等呈显著负相关。

|
| 图 7 干旱季节和非干旱季节浮游动物群落结构与环境因子的相关性 Fig. 7 The correlation between community structure of zooplankton and environmental factors in drought season and non-drought season “*”表示显著相关(P<0.05), “**”表示极显著相关(P<0.01);SD:透明度;WT:水温;DO:溶解氧;Cond:电导率;COD:化学需氧量;Chl.a:叶绿素a;TN:总氮;TP:总磷;NO3--N:硝态氮;NH4+-N:氨氮;WL:水位;H′:Shannon-Wiener指数;J:Pielou指数 |
通过对浮游动物丰度数据进行DCA分析, 发现干旱季节和非干旱季节最大梯度长度分别为3.40和3.44, 介于3—4之间, 故选取冗余分析(RDA)方法探究多种环境因子对浮游动物的影响(图 8)。干旱季节的前两轴特征值分别为1.11和0.17, 累计解释率为87.23%, 主要受WT、WL、NO3--N、NH4+-N等因子的影响;镰状臂尾轮虫与Chl.a、无节幼体与pH、剪形臂尾轮虫与WT均呈正相关。非干旱季节的前两轴特征值分别为0.42和0.24, 累计解释率为68.37%, 主要受COD、WL的影响;汤匙华哲水蚤与NH4+-N、无节幼体与DO、对棘异尾轮虫与WT、曲腿龟甲轮虫与WL均呈正相关。

|
| 图 8 干旱季节和非干旱季节浮游动物与环境因子的冗余分析 Fig. 8 Redundancy analysis of zooplankton and environmental factors in drought season and non-drought season RDA:冗余分析;sp1:长额象鼻溞;sp2:简弧象鼻溞;sp3:汤匙华哲水蚤;sp4:球状许水蚤;sp5:无节幼体;sp6:桡足幼体;sp7:广布中剑水蚤;sp8:曲腿龟甲轮虫;sp9:镰状臂尾轮虫;sp10:角突臂尾轮虫;sp11:剪形臂尾轮虫;sp12:裂足臂尾轮虫;sp13:等棘异尾轮虫;sp14:对棘异尾轮虫;sp15:晶囊轮虫;sp16:针簇多肢轮虫 |
本研究发现鄱阳湖浮游动物群落存在明显的季节变化特征, 干旱季节(夏季和秋季)的物种数大于非干旱季节(春季和冬季), 与杨潇等[30]的研究一致;在干旱季节, 浮游动物丰度、生物量均为夏季显著高于秋季, 在秋季达到一年中的最低值, 与前人研究不完全一致[10]。浮游动物Shannon-Wiener指数为夏季显著高于秋冬两季[31], 主要由于夏季气温较高, 浮游动物食物来源丰富, 适宜于生长繁殖。Pielou指数呈现春季显著高于秋季的特征, 而陈业等[32]通过对洪泽湖的研究发现, 浮游动物的Pielou指数秋季最高, 与水位的变化有着密切关系。浮游动物优势种随季节变化较为明显[31], 春季、秋季和冬季均表现为单一物种占绝对优势;优势种的数目越多群落结构越复杂、稳定[33], 夏季优势种最多。简弧象鼻溞为非干旱季节的主要优势种, 是春季的标志性浮游动物类群[34];干旱季节的主要优势种为无节幼体, 丰度分别占夏季、秋季总丰度的15.58%、50.09%, 同时夏季水中较低的氨氮含量可能导致大型浮游动物如长额象鼻溞、广布中剑水蚤等的出现, 符合湖库的物种组成[35]。综上所述, 浮游动物群落结构的稳定性呈现夏季高于其他季节的特征。
从空间分布来看, 鄱阳湖浮游动物的物种数、丰度、生物量均表现为南部湖区大于北部湖区的格局。陈佳琪等[36]的研究表明鄱阳湖浮游动物丰度呈现由南向北逐渐减少的趋势, 与本研究结果一致。Shannon-Wiener指数、Pielou指数表现为南部湖区大于北部湖区的特征, 这是由于鄱阳湖南部湖区分布着赣江南支入湖口、抚河入湖口及信江入湖口, 三条支流汇入湖泊后水体流速变缓, 为浮游动物提供了良好的生存环境。鄱阳湖不同区域的水体流速存在较大差异, 通常表现为北部湖区的水体流速大于南部湖区[37], 稳定的水文状态有利于浮游动物的生长繁殖。因此, 南部湖区的浮游动物群落结构特征优于北部湖区。
3.2 水文干旱年份浮游动物群落结构的主要影响因素鄱阳湖与长江及五河形成了复杂的江-河-湖系统, 浮游动物群落受各种水文、水环境因子的综合影响较大[22]。经过Pearson相关性分析和RDA分析, 表明干旱季节的浮游动物主要受水温、水位、硝态氮、氨氮等环境因子的共同作用, 而非干旱季节受水位、化学需氧量的影响较大。
水温是影响浮游动物群落结构的重要因子。Liu等[8]发现水温的季节波动影响鄱阳湖浮游动物(枝角类和桡足类)的群落结构, 很好的解释了本研究发现的浮游动物丰度随季节发生由枝角类到轮虫再到桡足类的更替规律。水温过高或过低, 都会对浮游动物休眠卵的孵化、生长和繁殖产生不利的影响[38—39], 有研究表明, 当温度为30 ℃时, 轮虫的净繁殖率最高[40], 鄱阳湖夏季适宜的水温使得轮虫的丰度占比最大。同时适宜的水温有利于浮游植物的生长[41], 春季浮游植物生物量随水温升高而增加[42], 为浮游动物提供了更丰富的食物来源。因此, 水温通过直接和间接的方式影响浮游动物的季节性演替[43]。
硝态氮和氨氮是影响浮游动物群落结构的重要因素[44]。Calbet等[45]发现浮游动物在贫营养盐的水域摄食率较高, 与本研究浮游动物群落结构与硝态氮、氨氮呈负相关的结果相一致。有研究表明, 较高浓度的氨氮对浮游动物的部分物种具有毒害作用, 会影响其生长和繁殖[46], 因此在夏季营养盐浓度较低的情况下, 浮游动物丰度和生物量最大。此外, 浮游动物主要以浮游植物为食物来源, 营养盐是浮游植物生长繁殖的物质基础[47]。随浮游植物数量增多而对营养盐的需求增大, 为浮游动物提供充足食物的同时, 导致夏季营养盐浓度低于其他季节, 硝态氮和氨氮通过促进浮游植物的生长, 进而促进浮游动物的繁殖[48]。
对于过水性的鄱阳湖而言, 平均换水周期约为30天[49], 年内十余米的水位波动意味着湖泊状态不稳定, 而在湖泊状态稳定、湖水流速缓慢的情况下才能为浮游动物提供有利的生存条件。相对于正常水文年, 极端水文干旱年的鄱阳湖夏季水位变化更大, 秋季水位更低, 导致湖面减小和流速加快, 严重影响了浮游动物的正常生命活动和生存空间。浮游动物群落结构与水位呈正相关关系, 低水位对其生长繁殖起到了一定的限制作用, 同时水位波动也对湖泊水质产生较大的影响[50], 进而影响浮游动物的群落结构。因此, 水位是浮游动物群落结构发生季节变化的主要原因之一[51], 也是导致极端水文干旱年浮游动物主要影响因素与其他年份不同的重要原因。
化学需氧量的大小可以表征水体中有机碎屑的含量[52], 而有机碎屑是浮游动物的主要食物来源。同时, 丰富的有机质能促进浮游植物生物量的增长, 以浮游植物为食的浮游动物生物量也会相应升高。靳萍等[53]的研究表明浮游动物丰度与化学需氧量呈极显著正相关, 本研究发现浮游动物Shannon-Wiener指数与化学需氧量亦呈显著正相关。此外, 化学需氧量还通过影响叶绿素a含量进而对浮游动物造成影响[54]。化学需氧量通过直接和间接的方式促进了浮游动物的生长, 亦是浮游动物群落的重要影响因素。
3.3 干旱与常规年份的鄱阳湖浮游动物群落对比分析通过对鄱阳湖浮游动物群落的相关研究进行汇总对比(表 3), 进而探讨鄱阳湖浮游动物群落结构特征在极端水文干旱年与常规水文年的异同。本研究表明, 极端水文干旱年的鄱阳湖浮游动物群落季节差异非常大, 夏季浮游动物丰度、生物量分别是秋季的8.37、5.5倍, 秋季的丰度、生物量亦小于冬春季节, 与陈佳琪等[36]在鄱阳湖的研究不一致。相对于历史常规年份[30, 36], 极端水文干旱年的鄱阳湖浮游动物物种数、丰度、生物量均明显偏低, 特别是夏季的物种数仅为历史同期数据的57.33%—63.24%, 丰度仅为历史同期数据的1.19%—2.29%, 生物量仅为历史同期数据的9.58%。极端水文干旱年的鄱阳湖浮游动物年平均丰度和生物量分别为33.83个/L和0.35 mg/L, 仅为鄱阳湖都昌保护区常规监测年份的15.82%、23.18%[31];与鄱阳湖碟形湖的常规监测年份数据相比[10, 55], 极端水文干旱年的浮游动物丰度、生物量亦显著减少。极端水文干旱年的鄱阳湖浮游动物多样性指数与常规年份差异不明显, 优势种在干旱季节主要为无节幼体, 并具有阶段性变化的特点, 与非干旱季节和历史数据不完全一致。由此可见, 极端水文干旱下鄱阳湖浮游动物群落呈现出比历史常规年份更不稳定的结构特征。
| 研究区域 Study areas |
采样时间 Sample time |
物种数 Number of species |
丰度 Abundance/(个/L) |
生物量 Biomass/(mg/L) |
Shannon-Wiener指数 Shannon-Wiener index |
Pielou指数 Pielou index |
优势种 Dominant species |
参考文献 References |
| 鄱阳湖 | 干旱年春季 | 26 | 20.17 | 0.20 | 1.89 | 0.91 | 简弧象鼻溞、对棘异尾轮虫等 | 本研究 |
| 鄱阳湖 | 干旱年夏季 | 43 | 72.21 | 0.55 | 2.47 | 0.85 | 镰状臂尾轮虫、无节幼体等 | 本研究 |
| 鄱阳湖 | 干旱年秋季 | 31 | 8.63 | 0.10 | 1.51 | 0.76 | 无节幼体、简弧象鼻溞等 | 本研究 |
| 鄱阳湖 | 干旱年冬季 | 24 | 21.06 | 0.41 | 1.17 | 0.77 | 简弧象鼻溞、无节幼体等 | 本研究 |
| 鄱阳湖 | 常规年夏季 | 75 | 6049.82 | 5.74 | 2.75 | 0.57 | 简弧象鼻溞、广布中剑水蚤等 | [30] |
| 鄱阳湖 | 常规年秋季 | 53 | 4489.35 | 1.45 | 2.69 | 0.69 | 右突新镖水蚤、简弧象鼻溞等 | [30] |
| 鄱阳湖 | 常规年夏季 | 68 | 3150.37 | / | 2.21 | 0.73 | 广生多肢轮虫、暗小异尾轮虫等 | [36] |
| 鄱阳湖 | 常规年秋季 | 60 | 835.92 | / | 1.76 | 0.79 | 螺形龟甲轮虫、广生多肢轮虫等 | [36] |
| 鄱阳湖 | 常规年冬季 | 38 | 145.86 | / | 1.24 | 0.84 | 广生多肢轮虫、尖尾疣毛轮虫等 | [36] |
| 鄱阳湖 | 常规年春季 | 71 | 484.76 | / | 1.68 | 0.75 | 螺形龟甲轮虫、广生多肢轮虫等 | [36] |
| 鄱阳湖都昌保护区 | 常规年 | 31 | 213.81 | 1.51 | 1.65 | 0.77 | 简弧象鼻溞、颈沟基合溞等 | [31] |
| 鄱阳湖碟形湖 | 常规年春季 | 28 | 612.08 | 2.67 | 1.64 | 0.52 | / | [10] |
| 鄱阳湖碟形湖 | 常规年夏季 | 88 | 580.38 | 1.74 | 2.85 | 0.76 | / | [10] |
| 鄱阳湖碟形湖 | 常规年秋季 | 72 | 524.53 | 2.51 | 2.71 | 0.74 | / | [10] |
| 鄱阳湖碟形湖 | 常规年冬季 | 65 | 77.74 | 0.18 | 2.54 | 0.71 | / | [10] |
| 鄱阳湖碟形湖 | 常规年夏季 | 65 | 519.77 | 0.92 | / | / | 盖氏晶囊轮虫、广布多肢轮虫等 | [55] |
| 其中“/”为文献中未涉及 | ||||||||
不同水文年鄱阳湖浮游动物群落的主要影响因素存在差异。在历史常规年份, 于新平等[31]对鄱阳湖都昌自然保护区浮游动物进行调查研究, 认为浮游动物群落结构主要受总氮、叶绿素a、水位和水温等因素的影响, 陈佳琪等[36]的研究表明水温、叶绿素a、电导率是造成鄱阳湖浮游动物群落季节差异的主要环境因子。其中, 水位、水温亦是影响极端水文干旱年浮游动物群落的主要因素, 由于极端水文干旱下秋季水位过低, 硝态氮、化学需氧量呈现与往年不同的变化趋势[31, 36], 浮游动物的生存环境和食物来源受到影响, 造成秋季的丰度和生物量均小于其他季节, 呈现出极端水文干旱年不同于历史常规年份的季节特征。同时, 极端水文干旱年的鄱阳湖在干旱季节降水偏少、高温持续时间较长[56], 这种不同于往年的水文气象干旱现象导致鄱阳湖枯水期提前, 丰水期水位远低于正常年份, 引起水体流速、水环境因子及浮游动物生存空间的急剧变化, 进而使得浮游动物的物种数、丰度、生物量均呈现低于历史常规年份的特征, 也是浮游动物群落结构的稳定性较往年减弱的重要原因。
4 结论通过对极端水文干旱年的鄱阳湖浮游动物展开季度调查, 共鉴定浮游动物70种, 全年物种数均以轮虫占优。鄱阳湖浮游动物丰度和生物量范围分别为0—152.67个/L和0—1.52 mg/L, 季节上呈现夏季最高、秋季最低的特征, 空间上表现为南部湖区高于北部湖区的格局。鄱阳湖浮游动物多样性和优势种具有典型的季节特征, 夏季Shannon-Wiener指数最高, 春季Pielou指数显著高于秋季和冬季, 但在空间上无显著差异;优势种的数量夏季最多、秋季最少。鄱阳湖浮游动物在干旱季节主要受水温、水位、硝态氮、氨氮等因素的共同影响, 而在非干旱季节受化学需氧量和水位的影响较大。相对于常规水文年, 极端水文干旱年的鄱阳湖浮游动物物种数、丰度和生物量均明显降低。
| [1] |
Qin H M, Cao X Y, Cui L Y, Lv Q, Chen T T. The influence of human interference on zooplankton and fungal diversity in Poyang Lake watershed in China. Diversity, 2020, 12(8): 296. DOI:10.3390/d12080296 |
| [2] |
Yuliana, Mutmainnah. Assessment of coastal water productivity of Fitu Village, Ternate Island based on its zooplankton community structure. IOP Conference Series: Earth and Environmental Science, 2021, 890(1): 012011. DOI:10.1088/1755-1315/890/1/012011 |
| [3] |
Kim H W, Hwang S J, Joo G J. Zooplankton grazing on bacteria and phytoplankton in a regulated large river (Nakdong River, Korea). Journal of Plankton Research, 2000, 22(8): 1559-1577. DOI:10.1093/plankt/22.8.1559 |
| [4] |
Wei W Z, Chen R M, Wang L F, Fu L X. Spatial distribution of crustacean zooplankton in a large river-connected lake related to trophic status and fish. Journal of Limnology, 2017, 76(3): 546-554. |
| [5] |
Gomes L F, Pereira H R, Gomes A C A M, Vieira M C, Martins P R, Roitman I, Vieira L C G. Zooplankton functional-approach studies in continental aquatic environments: a systematic review. Aquatic Ecology, 2019, 53(2): 191-203. DOI:10.1007/s10452-019-09682-8 |
| [6] |
王西锋, 万军芳, 王永平, 孙长顺, 李明, 胡恩. 基于浮游动物群落特征的渭河橡胶坝景观河流水质生物学评价. 河南师范大学学报: 自然科学版, 2022, 50(4): 117-122. |
| [7] |
宋全健, 丰美萍, 边文华, 程晓鹏, 汪振华, 章守宇, 刘洪生. 大陈岛海域浮游动物群落的季节变化及其影响因素. 生态学报, 2023, 43(17): 1-12. DOI:10.20103/j.stxb.202207091960 |
| [8] |
Liu B G, Wu J Y, Hu Y, Wang G X, Chen Y W. Seven years study of the seasonal dynamics of zooplankton communities in a large subtropical floodplain ecosystem: a test of the PEG model. International Journal of Environmental Research and Public Health, 2022, 19(2): 956. DOI:10.3390/ijerph19020956 |
| [9] |
Krylov A V. Interannual changes in the summer zooplankton in the lakes of the Khopyor River flood plain. Biology Bulletin, 2015, 42(10): 891-898. DOI:10.1134/S1062359015100076 |
| [10] |
Hu B J, Hu X R, Nie X, Zhang X K, Wu N C, Hong Y J, Qin H M. Seasonal and inter-annual community structure characteristics of zooplankton driven by water environment factors in a sub-lake of Lake Poyang, China. PeerJ, 2019, 7: e7590. DOI:10.7717/peerj.7590 |
| [11] |
宋高飞, 朱宇轩, 米武娟, Anila P. Ajayan, 毕永红. 武汉市湖泊浮游动物群落特征及其影响因素. 河南师范大学学报: 自然科学版, 2022, 50(3): 135-142. |
| [12] |
张建波, 王丑明, 黄代中, 张屹. 洞庭湖后生浮游动物群落结构及水质生物学评价. 长江流域资源与环境, 2023, 32(2): 394-402. |
| [13] |
李静, 梁阳阳, 唐晓先, 韩琦, 尹峰, 郭匿春, 卢文轩. 巢湖后生浮游动物群落及不同湖区营养状态评价. 水生态学杂志, 2023, 44(1): 73-81. |
| [14] |
Marques S C, Primo A L, Martinho F, Azeiteiro U M, Pardal M Â. Shifts in estuarine zooplankton variability following extreme climate events: a comparison between drought and regular years. Marine Ecology Progress Series, 2014, 499: 65-76. DOI:10.3354/meps10635 |
| [15] |
聂雪, 胡旭仁, 刘观华, 金斌松, 秦海明. 鄱阳湖子湖"堑秋湖"过程中水位变化对浮游动物群落结构的影响. 水生生物学报, 2019, 43(2): 402-414. |
| [16] |
Chen G J, Dalton C, Taylor D. Cladocera as indicators of trophic state in Irish Lakes. Journal of Paleolimnology, 2010, 44(2): 465-481. DOI:10.1007/s10933-010-9428-2 |
| [17] |
Ginatullina E, Atwell L, Saito L. Resilience and resistance of zooplankton communities to drought-induced salinity in freshwater and saline lakes of Central Asia. Journal of Arid Environments, 2017, 144: 1-11. DOI:10.1016/j.jaridenv.2017.04.010 |
| [18] |
Carrasco N K, Perissinotto R, Pillay D. Zooplankton of the St. Lucia Estuary during the current drought cycle: a comparison between open- and closed-mouth conditions. Marine Ecology Progress Series, 2010, 399: 157-171. DOI:10.3354/meps08355 |
| [19] |
Primo A L, Azeiteiro U M, Marques S C, Martinho F, Pardal M Â. Changes in zooplankton diversity and distribution pattern under varying precipitation regimes in a southern temperate estuary. Estuarine, Coastal and Shelf Science, 2009, 82(2): 341-347. DOI:10.1016/j.ecss.2009.01.019 |
| [20] |
Marques S C, Azeiteiro U M, Martinho F, Pardal M Â. Climate variability and planktonic communities: the effect of an extreme event (severe drought) in a southern European Estuary. Estuarine, Coastal and Shelf Science, 2007, 73(3/4): 725-734. |
| [21] |
Simões N R, Lansac-Tôha F A, Bonecker C C. Drought disturbances increase temporal variability of zooplankton community structure in floodplains. International Review of Hydrobiology, 2013, 98(1): 24-33. DOI:10.1002/iroh.201201473 |
| [22] |
万荣荣, 杨桂山, 王晓龙, 秦年秀, 戴雪. 长江中游通江湖泊江湖关系研究进展. 湖泊科学, 2014, 26(1): 1-8. |
| [23] |
夏军, 陈进, 佘敦先. 2022年长江流域极端干旱事件及其影响与对策. 水利学报, 2022, 53(10): 1143-1153. |
| [24] |
雷声, 石莎, 屈艳萍, 刘业伟, 廖金源, 张阳, 张瑛, 李斯颖, 董帮兴. 2022年鄱阳湖流域特大干旱特征及未来应对启示. 水利学报, 2023, 54(3): 333-346. |
| [25] |
邹亮华, 邹伟, 张庆吉, 李颖, 龚志军, 张艳杰, 鲁顺保, 蔡永久. 鄱阳湖大型底栖动物时空演变特征及驱动因素. 中国环境科学, 2021, 41(6): 2881-2892. |
| [26] |
李安梅, 李言阔, 邵瑞清, 申锦, 钱磊, 张超, 王贤芳, 方彭军. 鄱阳湖鹤类越冬种群数量动态和空间分布变化. 生态与农村环境学报, 2022, 38(6): 777-786. |
| [27] |
章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社, 1991.
|
| [28] |
陈昕, 李喆, 唐富江, 齐妍, 高文燕, 鲁万桥. 连环湖两水体浮游动物群落结构特征及水质评价. 水生态学杂志, 2020, 41(6): 89-97. |
| [29] |
von Wehrden H, Hanspach J, Bruelheide H, Wesche K. Pluralism and diversity: trends in the use and application of ordination methods 1990-2007. Journal of Vegetation Science, 2009, 20(4): 695-705. DOI:10.1111/j.1654-1103.2009.01063.x |
| [30] |
杨潇, 马吉顺, 张欢, 周琼. 鄱阳湖不同水文期浮游生物群落结构特征和影响因素及水质评价. 水生生物学报, 2021, 45(5): 1093-1103. |
| [31] |
于新平, 陈宇炜, 王俊颉, 李瀚成, 邓枫, 徐光锋, 夏雨, 刘金福. 鄱阳湖都昌自然保护区浮游甲壳动物群落时空分布特征及其驱动因子. 湖泊科学, 2023, 35(3): 998-1008. |
| [32] |
陈业, 彭凯, 张庆吉, 蔡永久, 张永志, 龚志军, 项贤领. 洪泽湖浮游动物时空分布特征及其驱动因素. 环境科学, 2021, 42(8): 3753-3762. |
| [33] |
代培, 刘凯, 周彦锋, 周游, 徐跑. 太湖五里湖湖滨带浮游动物群落结构特征. 水生态学杂志, 2019, 40(1): 55-63. |
| [34] |
张群, 许旭明, 陈倩. 潮白河流域水库浮游动物群落季节变化特征及其驱动因素. 北京大学学报: 自然科学版, 2023, 59(2): 290-300. |
| [35] |
白雪兰, 范泽宇, 徐聚臣, 林深, 马帅兵, 黄路全, 李金能, 侯杰, 何绪刚. 洈水水库浮游生物群落结构特征及水质评价. 华中农业大学学报, 2023, 42(1): 118-127. |
| [36] |
陈佳琪, 赵坤, 曹玥, 吴波, 庞婉婷, 尤庆敏, 王全喜. 鄱阳湖浮游动物群落结构及其与环境因子的关系. 生态学报, 2020, 40(18): 6644-6658. |
| [37] |
欧阳千林, 王婧. 鄱阳湖河湖两相下湖流和水质变化特征研究. 江西水利科技, 2018, 44(5): 356-362. |
| [38] |
Lenz P H, Hower A E, Hartline D K. Temperature compensation in the escape response of a marine copepod, Calanus finmarchicus (Crustacea). The Biological Bulletin, 2005, 209(1): 75-85. DOI:10.2307/3593143 |
| [39] |
Devreker D, Souissi S, Seuront L. Effects of chlorophyll concentration and temperature variation on the reproduction and survival of Temora longicornis (Copepoda, Calanoida) in the Eastern English Channel. Journal of Experimental Marine Biology and Ecology, 2005, 318(2): 145-162. DOI:10.1016/j.jembe.2004.12.011 |
| [40] |
宁乐锋, 席贻龙, 孙强, 周澳. 温度和食物密度对裂足臂尾轮虫(Brachionus diversicornis)生命表统计学参数的影响. 湖泊科学, 2013, 25(2): 295-301. DOI:10.3969/j.issn.1003-5427.2013.02.018 |
| [41] |
Wu Z S, Cai Y J, Liu X, Xu C P, Chen Y W, Zhang L. Temporal and spatial variability of phytoplankton in Lake Poyang: the largest freshwater lake in China. Journal of Great Lakes Research, 2013, 39(3): 476-483. DOI:10.1016/j.jglr.2013.06.008 |
| [42] |
周慜, 何亮, 黄建美, 张萌, 王启沛, 万禀颢, 熊名日, 刘足根. 鄱阳湖-珠湖浮游植物群落结构时空变化特征与环境因子的关系. 生态科学, 2021, 40(4): 36-46. |
| [43] |
朱旭宇, 黄伟, 曾江宁, 江志兵, 刘晶晶, 徐晓群, 陈全震. 氮磷比对冬季浮游植物群落结构的影响. 应用与环境生物学报, 2013, 19(2): 293-299. |
| [44] |
Li C C, Feng W Y, Chen H Y, Li X F, Song F H, Guo W J, Giesy J P, Sun F H. Temporal variation in zooplankton and phytoplankton community species composition and the affecting factors in Lake Taihu: a large freshwater lake in China. Environmental Pollution, 2019, 245: 1050-1057. DOI:10.1016/j.envpol.2018.11.007 |
| [45] |
Calbet A, Landry M R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnology and Oceanography, 2004, 49(1): 51-57. DOI:10.4319/lo.2004.49.1.0051 |
| [46] |
Xiang F H, Yang W, Chen Y F, Yang Z. Acute toxicity of nitrite and ammonia to Daphnia similoides of different developmental stages: using the modified Gaussian model to describe. Bulletin of Environmental Contamination and Toxicology, 2010, 84(6): 708-711. DOI:10.1007/s00128-010-0017-x |
| [47] |
Tian W, Zhang H Y, Zhao L, Xiong Y W, Huang H. Effects of environmental factors on the temporal stability of phytoplankton biomass in a eutrophic man-made lake. Water, 2016, 8(12): 582. DOI:10.3390/w8120582 |
| [48] |
彭颖, 崔键, 李金凤, 杜易, 邱寒, 崔建伟, 陈建, 程铮, 葛正顺, 张晗. 连云港市徐圩新区张圩湖和云湖浮游动物和浮游植物群落组成研究. 湿地科学, 2023, 21(1): 93-103. |
| [49] |
李云良, 姚静, 李梦凡, 张奇. 鄱阳湖换水周期与示踪剂传输时间特征的数值模拟. 湖泊科学, 2017, 29(1): 32-42. |
| [50] |
杜冰雪, 徐力刚, 张杰, 范宏翔, 程俊翔, 黄涛, 姜加虎. 鄱阳湖富营养化时空变化特征及其与水位的关系. 环境科学研究, 2019, 32(5): 795-801. |
| [51] |
刘宝贵, 刘霞, 吴瑶, 钟正, 陈宇炜. 鄱阳湖浮游甲壳动物群落结构特征. 生态学报, 2016, 36(24): 8205-8213. DOI:10.5846/stxb201505070941 |
| [52] |
刘倩纯, 胡维, 葛刚, 熊勇, 赖劲虎, 吴兰. 鄱阳湖枯水期水体营养浓度及重金属含量分布研究. 长江流域资源与环境, 2012, 21(10): 1230-1235. |
| [53] |
靳萍, 代克岩, 杨程, 郭萌, 徐婷婷, 蔺庆伟, 马剑敏. 牧野湖浮游动物群落结构及其与环境因子的相关性研究. 水生态学杂志, 2013, 34(2): 53-61. |
| [54] |
邱小琮, 赵红雪, 孙晓雪. 沙湖浮游动物与水环境因子关系的多元分析. 生态学杂志, 2012, 31(4): 896-901. |
| [55] |
吕乾, 胡旭仁, 聂雪, 欧阳珊, 王超, 秦海明. 鄱阳湖丰水期水位波动对浮游动物群落演替的影响. 生态学报, 2020, 40(4): 1486-1495. |
| [56] |
贾建伟, 王栋, 徐伟峰, 刘昕. 2022年鄱阳湖流域干旱综合评估及成因分析. 人民长江, 2023, 54(2): 36-42. |