 2023, Vol. 43
2023, Vol. 43文章信息
- 刘燕, 张凌楠, 刘晓宏, 曾小敏, 贾瑞萱
- LIU Yan, ZHANG Lingnan, LIU Xiaohong, ZENG Xiaomin, JIA Ruixuan
- 干旱胁迫植物个体生理响应及其生态模型预测研究进展
- Research progress from individual plant physiological response to ecological model prediction under drought stress
- 生态学报. 2023, 43(24): 10042-10053
- Acta Ecologica Sinica. 2023, 43(24): 10042-10053
- http://dx.doi.org/10.20103/j.stxb.202304070703
-
文章历史
- 收稿日期: 2023-04-07
- 采用日期: 2023-11-13

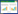




随着全球的气温上升, 严重或持续的干旱事件频发, 许多植物的生存面临着严重的威胁。干旱事件导致植物水分蒸发加剧, 降低了土壤水分的可利用性, 强烈影响了植物的水分运输过程, 这一系列过程导致冠层叶片枯萎, 枝条枯死甚至整个植物死亡的现象引起学者的广泛关注。干旱引发植物死亡的背后生理机制以及未来气候变化下植物的响应模式逐渐成为热点研究领域。
植物对干旱的响应是一个涉及基因表达、代谢调节、生理适应以及生态互作的复杂过程。首先与抗旱有关的基因表达增强, 如参与渗透代谢、次级代谢产物合成和激素合成的基因, 有助于快速诱导反应机制和串扰, 进而通过影响气孔来减少水分损失。其次植物通过适当调整结构来提高水分利用效率, 如适当提高叶脉与气孔密度, 增加木质部输水结构储水、保水能力, 这些都是植物应对干旱的策略。植物结构与功能对气候和环境的适应性会成为影响区域甚至全球森林生态系统组成和分布的重要因素。
本文主要通过综合植物抵御干旱胁迫的重要机理过程, 探讨植物水力特性与解剖结构之间的联系, 并进一步研究水力特性与光合作用的关系。理清植物在发生水分胁迫时, 怎样调整解剖结构与生理机制相适应来响应环境气候变化, 期望基于植物水力、解剖和光合之间的联系改善植物水力模型, 推动其与生理生态等多角度模型结合, 为今后评估气候巨变带来植物死亡风险提供依据。
1 干旱胁迫下植物的生理响应 1.1 干旱胁迫下植物渗透物质调节干旱胁迫下, 植物会通过渗透调节来适应水分不足的环境[1]。渗透调节是植物适应环境水分胁迫的一种生理反应, 植物通过渗透调节物质积累, 增加细胞渗透压, 降低水分散失速率, 提高抗旱能力。其中渗透调节物质分为有机渗透剂(脯氨酸、甜菜碱等)[2]和无机离子(Ca2+、K+等)两大类。前者通过对酶、蛋白质和生物膜起保护作用来维持细胞内代谢正常进行, 后者主要改变离子通道和蛋白质活性来调节细胞渗透势稳定细胞形态。
植物体中, 脯氨酸可由谷氨酸和鸟氨酸分别经过吡咯啉-5-羧酸合成酶(Pyrroline-5-carboxylate Synthetase, P5CS)和吡咯啉-5-羧酸还原酶(Pyrroline-5-carboxylate Reductase, P5CR)在细胞质和叶绿体中转化而成[3—4], 而过多的脯氨酸又会在线粒体中被脯氨酸脱氢酶转化为谷氨酸。此代谢平衡会在植物受到干旱胁迫时被破坏, 如P5CS基因水平表达过高或线粒体功能受损导致脯氨酸降解受阻等因素, 都会导致脯氨酸含量增加。因此很多研究将植物体内脯氨酸含量作为指示植物是否受到水分胁迫的指标。但更多研究结果表明, 脯氨酸含量只是在水分胁迫过程中迅速积累, 但在解除胁迫后又会恢复正常水平, 所以脯氨酸的含量变化无法反应植物抗旱性。
有机渗透剂的合成通常是一个缓慢的过程, 需要消耗大量底物和能量, 这一过程可能只在植物发生严重脱水时才会启动[5]。相比之下, 通过离子通道或转运蛋白从环境中吸收无机离子来增加细胞离子浓度, 是一种更高效的渗透调节方式[6]。例如, 植物在接收到干旱信号时会增加体内Ca2+的含量, 其可以与钙调蛋白结合, 进一步促进植物体内多种抗氧化酶的激活来抵御水分亏缺[7]。同时Ca2+作为胁迫信号传递的第二信使, Ca2+的含量增加, 使钙依赖蛋白激酶磷酸化下游靶蛋白[8], 与脱落酸一起介导气孔运动来减少水分散失, 有利于植物抗旱。K+是植物抵御干旱胁迫的重要保护剂, 植物细胞恢复膨压需要根系细胞质膜上的K+转运蛋白介导吸收K+、Cl-和Na+来参与渗透调节[9]。K+含量增加会诱导溶质积累, 从而降低渗透电位, 并有助于在渗透胁迫下维持植物细胞的膨压。K+浓度除了直接影响渗透压的控制外, 还参与了植物细胞膜完整性维持以及酶活性控制的间接过程。K+通过提供适当的离子强度或与酶的特定结合位点结合参与催化来激活酶, 从而发挥辅因子的作用, 使各种酶正常进行代谢和生理机制[10]。此外, 充足的钾可以增加干旱胁迫下植物的总干物质积累[11], 且木质部汁液中钾浓度增加有利于水力传导率的提高[12], 使植物逐渐适应干旱环境。
1.2 干旱胁迫下植物激素脱落酸调节脱落酸(Abscisic Acid, ABA)是一种植物响应干旱胁迫的关键激素。在干旱胁迫下, 它由植物根部产生, 作为一种信号被运输到叶片, 通过诱导气孔关闭, 从而防止植物水分流失, 提高植物抗旱性[13]。也有研究证实, 缺水期间, ABA也能在叶片中合成, 然后转运到其他器官中[14], 而且保卫细胞也可以合成ABA响应水分亏缺[15], 这种方式有利于植物快速感知空气中水分变化, 避免因根部传递土壤缺水信号到地上部分的延迟。
为了应对干旱, ABA的快速积累会引发相关信号成分的串扰和调节, 触发细胞质膜阴离子通道和钾离子外向通道开放, 导致保卫细胞的膨压降低, 气孔关闭[16]。ABA进入细胞内, 与对应受体蛋白(PYR/PYL/RCAR)结合[17], 形成的ABA与PYR/PYL/RCAR结合体进而与2Cs类蛋白磷酸酶(Protein Phosphatase 2Cs, PP2Cs)形成复合物, 促使PP2Cs解除对蔗糖非发酵相关蛋白激酶2(Sucrose non-fermenting 1-related Protein Kinases 2, SnRK2)的抑制作用[18], 使SnRK2被激活, 并磷酸化下游靶点调节质膜阴离子通道和钾离子外向通道[19], 使细胞质内Cl-和K+外流, 最终导致植物气孔开度受抑或气孔关闭(图 1)。研究发现裸子植物气孔导度在适度干旱下由叶片水势直接作用, 而在长期或严重干旱时, 气孔调控变为ABA介导的主动调节, 这分别反应了针叶树种中对ABA变化敏感的辐射松和水杉的等水调节策略, 以及柏科植物在严重干旱下的非等水调节策略。Mcadam等人[20]认为被子植物叶片中ABA水平增加的幅度取决于叶片膨压, 且不同物种叶片ABA水平显著升高与失水点膨压值有关。

|
| 图 1 植物体内水分运输示意图 Fig. 1 Diagram of water transportation in plants ABA: 脱落酸 |
植物的水分运输分为质外体运输和共质体运输(图 1)。将水分在相邻细胞的细胞壁和细胞间隙中运输称为质外体运输, 此过程无需跨膜, 运输速度较快;而水分通过各个细胞经胞间连丝形成的连续原生质体的运输称为共质体运输, 此时水分子需进行跨膜运输, 运输速度较慢。水通道蛋白(Aquaporins, AQPs)广泛存在于植物细胞膜上, 可以介导水分跨膜运输调节根部导水率增加水分吸收, 提高植物整体水分运输效率[21—22]。干旱胁迫下植物木质部薄壁组织细胞会产生大量AQPs, 加强质外体和共质体间的水分交换来响应水分胁迫[23]。AQPs在干旱胁迫下不仅能调节膜的传导率并加强水分渗透性, 也可作为快速响应水分胁迫信号的调节蛋白, 主动参与植株的干旱胁迫响应[24], 但AQPs的表达与活性也会受干旱的调节。
AQPs在植物根、茎、叶中均有表达, 具有透水能力的AQPs多集中在质膜内在蛋白(Plasma membrane Intrinsic Proteins, PIPs)和液泡膜内在蛋白(Tonoplast Intrinsic Proteins, TIPs)家族, 它们分别位于植物原生质膜和液泡膜内, 在促进水分运输和调节细胞膨压方面发挥重要作用[25—26]。AQPs通过一系列调控发挥作用, 使植物细胞和器官能够对周围环境的水分变化做出反应, 调控包括转录调控、翻译后修饰、门控机制和异聚化[27]。AQPs磷酸化调节可以提高或降低AQPs的活性, 从而影响植物水分运输, 是AQPs应对水分亏缺中的重要一环。植物中AQPs的磷酸化能够被ABA和钙调蛋白激酶等多种信号途径调节。植物体内ABA水平上升对AQPs的磷酸化会使AQPs减少对水分子的渗透, 从而减少水分流失。而当Ca2+浓度升高, 钙调蛋白激酶被激活并对AQPs进行磷酸化调节, 此方式会使AQPs增加对水分子的渗透, 达到促进水分吸收的目的。
研究发现AQPs有调节植物叶片导水率的功能。例如汞添加会阻断某些AQPs活性来降低水分在叶片中的运输, 从而降低叶片水力导度[28—29]。与此同时, 研究者经过实验发现TIPs主要与植物叶片干旱后的复水过程有关, 并且证实了编码TIPs的基因表达增强, 这表明TIPs有助于逆转干旱诱导的叶片水力导度减少[30]。
2 干旱胁迫下植物水力特性与其他生理指标关系植物的水力特性是评估植物在干旱条件下生存和适应能力的重要指标, 主要指植物在水分吸收、运输和蒸腾等方面的特性。我们熟知的水力特性指标基于压力—容积曲线计算的有膨压丧失点(ψtlp)、叶片相对含水量、饱和渗透势、叶片水容、细胞壁弹性模量等, 以及叶片导水率(Kleaf)、枝条导水率(Kbranch)。当植物遇到干旱胁迫时, Kleaf和Kbranch都会下降, Kleaf损失50% 对应的水势值为Pleaf-50, Kbranch损失50% 和80% 对应的水势值分别为P50和P80。P50和P80已被证实能够有效代表植物木质部水力脆弱性。植物枝叶的水力特性指标主要测定方法、水力特性间权衡与协调关系等方面研究相对成熟[31]。故本文将研究重点集中在植物水力特性与枝叶解剖结构及光合作用之间的关系上。
2.1 植物叶片水力特性与解剖结构关系叶片作为土壤-植物-大气连续体(Soil-Plant-Atmosphere Continuum, SPAC)的重要环节, 对环境变化十分敏感[32], 当植物生长受到水分限制, 其叶片更倾向于朝有利于提高水分利用效率的方向在结构上发生改变。例如改变叶脉密度、气孔密度和大小。通常, 高叶脉密度能减少水分运输距离, 极大提高水分运输效率。但有研究发现主脉密度与Kleaf没有明显关系[33], 反而水分运输效率取决于细脉密度[34]。究其原因, 在干旱环境下, 主脉导管更容易产生栓塞[35], 而叶片末端细脉是水分流向气孔的最终限速部分[36]。同时叶脉中较多的纤维细胞使叶组织机械功能增强, 植物采取提高叶脉密度的方式来抵御外界干扰, 从而延长叶片寿命来抵御干旱[37]。然而, 提高叶脉密度虽然是植物抵抗干旱胁迫的有利方式, 但其需要更多的碳投入, 因此叶脉系统建设受环境影响以及碳投入与遗传学的共同调控[38]。
气孔作为植物体与外界进行水分和气体交换的通道, 其解剖特征可以作为植物应对水分胁迫能力的一种指示。气孔的密度和大小多受环境水分情况影响, 在干旱环境中小而密的气孔具有更高的灵活性, 更利于植物在保证呼吸作用的同时保持水分, 但当干旱程度加剧, 气孔密度反而降低[39]。研究者普遍认为是水分胁迫限制叶面积扩张而导致气孔密度增加, 但随着干旱程度加剧, 气孔数目减少, 最终表现为气孔密度下降。
2.2 植物枝条水力特性与解剖结构关系茎作为连接植物根与叶的重要器官, 一方面承担着植物的水分和营养物质的运输作用, 另一方面通过机械支撑作用以保证植物健康地向上生长。研究茎的解剖结构与水力特性间的关系, 可以掌握植物水分运输情况以及抗旱机制, 为改良或增强植物抗旱性提供基本依据。有研究发现植物茎可以通过增大表皮细胞提高细胞的储水能力来应对干旱[40]。同时, 皮层中包含的薄壁组织和厚角组织也可以吸收和输送水分, 两者表面变硬程度也均与细胞失水情况有关。茎木质部的导管(或管胞)是植物水分运输的主要途径, 最内部的髓虽不能进行水分运输, 但排列紧密的髓细胞中含有大量含晶细胞与粘液细胞可以增大细胞内渗透压, 从而提高髓细胞保水能力, 因此髓越发达越有利于提高植物水分贮藏能力[41]。
木质部管道(导管或管胞)结构是植物水力特性研究的重点。管道作为植物水分运输的主要通道, 其直径越大、数量越多, 木质部的输水效率就越高[42], 但在水分亏缺时, 管道发生栓塞的风险也会增加[43]。而植物的栓塞抗性与抵御干旱胁迫能力间通常为正相关关系[44], 植物栓塞抗性越强, 其抗旱性就越强。于是, 研究管道结构特征与栓塞抗性间的关系将是对比植物抗旱性强弱的新切入口。李泽东等人对不同抗旱树种的木质部解剖特征间差异进行对比, 发现耐旱树种拥有较厚的导管壁和较大的最大导管直径, 同时两者都与潜在最大导水率呈显著正相关, 非耐旱树种则没有表现出此关系[45]。在一些干旱地区, 植物木质部导管直径小, 数量多, 但小径导管的导管壁更厚, 降低了植物导管发生栓塞的几率, 在一定程度上补偿了水分运输的有效性[46]。
纹孔是由初生细胞壁未经次生加厚留下凹陷形成的空隙, 它连接着相邻导管, 其上的纹孔膜主要控制水分在其间径向流动。植物进行水分运输不仅要克服重力和摩擦力, 同时也要克服水分经过穿过纹孔膜的水力阻力, 这部分阻力占木质部总水力阻力的50% 甚至更多[47—49], 成为影响植物整体水力效率的重要因素。大量研究发现, 纹孔膜边缘有无环面结构是裸子植物和被子植物木质部栓塞抗性存在差异的主要原因, 同时, 纹孔膜厚度[50], 纹孔膜上微孔的大小[51]会在一定程度上影响植物的栓塞抗性。最新研究纹孔膜与栓塞修复有关, 但这方面的研究还未得到强有力的证明。
2.3 植物叶片水力特性与光合作用关系气孔在光合作用中起到平衡水分损失与CO2捕获的重要作用[52], 同时其对植物叶水势的变化十分敏感。已有实验证实植物的水分运输严重丧失是气孔关闭的一个主要驱动因素[53]。因此, 植物体内水分运输的速度和效率不仅影响Kleaf, 也会直接影响光合速率。同时, Kleaf和光合作用之间存在耦合关系[54], 且二者间的协调被认为是由气孔导度(gs)决定的[55]。而gs与Kleaf之间也存在良好的相关性[56—57]。当植物面临水力风险时, 供应气孔的水力驱动不足致使gs下降, 最终影响光合碳同化[58]。gs不仅受土壤水分、CO2浓度等环境因素的影响, 也受到气孔密度、气孔长度等气孔特征的调控。Sack和Frole研究发现阳生植物的叶片导水率与气孔指数有关[59], 高气孔密度可以满足它们对水分的高需求, 小气孔和高密度的组合可以产生较高的气孔导度和光合生产力[60]。研究发现植物气孔密度与气孔长度间具有明显权衡关系[61], 但两者谁对gs起决定性作用尚有争议[62], 也有观点认为gs取决于两者共同协调结果。
叶脉的结构与功能会直接或间接影响植物水分调节和光合作用。其中叶脉密度会影响水分通过叶肉间隙的距离, 例如高叶脉密度会缩短水分的运输距离, 使植物的进行光合作用效率更高[63]。叶脉密度与气孔特征往往共同协调水分供给与需求的平衡。某些物种中高气孔密度通常会伴随高叶脉密度同时出现[64]。起初, Sack等人研究发现叶脉密度与单位叶面积上的气孔面积呈正相关, 也证明叶脉密度越高, 叶片的气孔密度和气孔导度也越高[65]。但有研究发现在高海拔或高饱和水汽压差(VPD)环境下, 植物气孔密度和叶脉密度间关系微弱甚至解耦[66—67]。这也说明植物叶脉密度与气孔特征间关系受环境因素影响, 两者通过不断协调以维持水力平衡, 提高光合作用。
3 植物水力特性完善生态系统中植物对气候响应 3.1 植物水力指标发展的生态意义在全球范围内, 植物的水力特性在描述生态群落和生态系统的功能方面具有重要意义[68—69]。植物水力性状的多样性可以调节生态系统在干旱胁迫趋势下的稳定性与恢复力, 可能对各生态系统结构功能产生重大影响[70]。有研究基于ψtlp与群落干旱指数关系发现ψtlp可以比较群落水平的抗旱性[71]。然而以ψtlp的差异来预测物种的耐旱性受很多因素的影响。因此需要综合植物其他水力特性才能更准确地阐明植物应对干旱的响应。
有研究发现物种在干旱中的表现与P50密切相关[72]。干旱下树木能否存活取决于其水势是否低于其死亡阈值(即致死水势), 裸子植物的致死水势接近其P50, 而被子植物的致死水势均低于其P50而更接近P88, 由于裸子植物和被子植物致死水势均与木材密度呈负相关, 将易于测得的木材密度作为干旱期间树木存活率预测指标可信度更强[73]。通过对亚热带常绿阔叶林中不同树种对极端干旱事件的响应与恢复研究发现, P50越大的树种存活较久, 其光合作用和蒸腾作用对干旱的抵抗力和干旱过后的恢复力也越强[74]。
现在针对水分胁迫下植物耐受阈值的水力特性指标研究越来越多, 但以单一指标, 例如ψtlp和P50的大小作为预测物种抗旱临界水平的指标还缺乏有力支撑。研究发现植物水力安全裕度(Hydraulic Safety Margin, HSM)即某一物种在自然条件下经历的最小叶片水势(ψmin)与P50差值被认为是评估植物的干旱死亡风险指标, 值越大, 表示植物历经栓塞可能性越低, 死亡风险越低。HSM的变化与森林群落的脆弱性相关[68], 可以预测物种死亡率[75], 并且相比于P50和木材密度等树木性状指标, HSM是森林生态系统对干旱响应指标中解释力最高的预测因子[70]。由于在不同程度的干旱下ψmin很难做到测量准确[68]。有学者提出用气孔关闭时对应的叶水势(ψclose)代替ψmin所得到的气孔安全阈值(Stomatal Safety Margin, SSM)来衡量植物抗旱的能力, 可以更好反应气孔和水力性状之间的协调性[76]。Chen等人在对温带阔叶树种死亡率预测的研究中, 发现相比P50和HSM, SSM能更准确地预测干旱胁迫下植物的死亡风险[77]。但目前该指标使用并不广泛, 需要更多的研究加以验证。
3.2 植物水力模型发展的生态意义在全球范围内, 气候变化导致的干旱强度增加, 可能使未来几十年内树木死亡更加频繁[78]。鉴于森林对生物化学循环和生态系统服务的重要性[79], 评估干旱期间树木死亡风险越来越值得被关注。最初的评估是基于气候指标、土壤水分等, 并未考虑干旱下树木死亡的机制。随着对树木死亡机制探索的深入进行, 研究发现树木在干旱胁迫下死亡的两个重要机制是: 水力失衡和碳饥饿[80]。虽然长期森林清查和遥感技术是检测树木死亡的主要手段, 但长期清查容易忽略短期干扰事件对植物死亡的影响[81], 而遥感监测对小尺度上个体树木死亡引起的森林冠层变化不敏感[82]。为了更准确判定树木死亡, 就需要结合植物水力学等研究对树木死亡风险进行全面的评估。要想实现从单一物种到区域或全球尺度上树木死亡风险评估, 通过模型手段十分重要。
近年来发展的植物水力学模型旨在建立植物水分吸收和散失对土壤和大气干旱的直接水力响应关系。基于木质部水分传输与土壤水分运动的相似性, Sperry等人将描述土壤水分运动的数学物理方程扩展至土壤-植物系统[83]。之后在此基础上, Sperry and Love[84]通过引入气孔对植物水分供需关系的调节, 建立了植物供需水力模型(即Sperry模型), 其可通过植物根区土壤水分分布和VPD直接计算SPAC系统中的水流通量、冠层导度以及系统内水势和水流阻力的分布, 为研究根区土壤水分垂向异质性对SPAC水分传输安全性的影响提供了重要工具。此后, Sperry基于植物水力特征和气孔优化理论再次对模型进行升级, 即考虑到为避免木质部管道过度栓塞而进行的气孔调节并兼顾光合作用最大化, 称其为光合增益与水力成本优化模型。该优化模型已被证明在树木、林分和生态系统尺度上改善了对干旱情况下蒸腾和碳同化速率的预测, 且已被用于预测干旱控制实验中白杨(Populus tremuloides)幼苗的死亡率[85]。与此同时, Eller等人基于不同的优化目标, 将优化模型改编为基于木质部水力学的气孔优化模型(SOX)[86], SOX相对简单、只需少量参数, 因此其很有力成为热带森林应对未来极端气候变化的大规模建模的首选者。例如JULES地表模型与SOX结合, 不仅克服了经验模型的局限性, 使模型模拟结果更接近观测结果, 而且提高植被对土壤干旱和气候变化的响应过程, 改善欧洲森林对干旱反应的预测[87]。在Sperry系列模型启发下, 研究者开发了一种便于应用的、简化的离散时间土壤-植物水力模型——SurEau模型。为了便于应用, 该模型只考虑根际和植株内的水分运输阻力, 对栓塞抗性值范围内植物水力失调的时间进行模拟[88]。基于SurEau模型预测法国中部欧洲山毛榉森林死亡情况, 发现即使是表面上安全的、湿度适中的法国中部森林, 预计在本世纪末之前也会出现严重的森林死亡[89]。SurEau-Ecos模型是在SurEau模型基础上, 阐明植物叶和茎的共质体及质外体水分运输情况, 并增加从林分到区域尺度树木干燥和死亡率预测的植物水力学模型。SurEau-Ecos模型能够对干旱引起的植物水力破坏进行区域尺度的预测, 确定最易受干旱条件影响的地区和生态系统, 还能评估森林可燃性的动态情况[90]。
植物水分运输的水力学机制与模型的不断完善可以提高干旱条件下植物蒸散发模拟的准确性, 对生态系统的生产力下降做出合理解释, 并在模拟植物抗旱性与死亡风险预测方面更具说服力。起初地表模型由于一直采用土壤水分胁迫方案(Soil Moisture Stress, SMS), 导致模型对生态系统的蒸散发估计值整体偏高。随后植物水分胁迫方案(Plant Hydraulic Stress, PHS)的提出, 避免SMS依赖经验公式的局限性, PHS以物理过程为基准, 在SMS的基础上优化了水力再分配模块, 提高干旱胁迫下植物对水分的利用率。利用PHS的地表模型可以提高预测植物水分利用的准确性, 更好地模拟植物的生长, 为气候变化下生态系统提供保护措施。据悉, CLM5是第一个结合PHS的地表模型, 结合后的模型可以更好地模拟植被的水分利用策略变化如何影响生态系统生产力对干旱事件的响应, 并提高热带森林对干旱响应的预测准确性[91]。而PHS与Noah-MP地表模型耦合(简称为Noah-MP-PHS模型)考虑了整个植物的水力策略, 综合根茎叶的水力特性, 强调水容量在植物应对干旱策略方面的重要性。有研究针对成熟混合阔叶林地块级尺度观测结果进行评估, 发现Noah-MP-PHS模型更真实地反应植物的水分胁迫, 强调植物蓄水能缓解干旱土壤条件下木质部的水分胁迫, 影响植物水分利用效率, 并为今后在更大尺度下模拟植被对气候变化的响应提供新的方案[92]。有研究基于供需水力模型, 对生物群落-生物地球化学模型中的SPAC水分运输过程进行了修正, 使改进后的模型有效地捕捉了黄土高原刺槐林冠层蒸腾、土壤水分、叶片水势和叶面积指数的动态变化, 并预测未来气候变化对黄土高原刺槐生长和干旱致死风险影响[93]。Tai等人[94]将供需水力模型并入陆地区域生态系统交换模拟器(TREES)中, 发现利用植物水力特征能极大提高模型解释白杨死亡模式的能力。随后将融入植物水力模型的TREES模型与可变饱和地下水模型相结合, 形成综合模型, 以量化干旱导致的加拿大西南部河岸棉白杨的死亡风险[95]。未来多模型耦合可能为森林生态系统功能变化和风险的评估提供了很好的基础(表 1)。水力模型模拟的水分运输过程的重要性日渐凸显, 但依然需要对其进行不断完善;水力模型与其他模型合理联用可能会极大提高森林生态系统对极端气候响应预测的准确性。
| 模型 Model |
水力模型部分类型 Types of hydraulic model parts |
水力模型部分原理 Principles of hydraulic model |
模型尺度 Model scale |
模型应用 Model application |
参考文献 References |
| Sperry 模型 Spery model |
电类比模型 | 达西定律 | 树木个体尺度 | 预测干旱引起的植物导水性、冠层、碳同化和生产力的损失 | [84] |
| Sperry 优化模型 Sperry optimization model |
/ | / | 叶片尺度、树木个体尺度、生态系统尺度 | 预测生长季节内植物死亡风险的水力阈值 | [96] |
| 结合 Sperry 模型的 TREES 模型 TREES integrates Sperry model |
电类比模型 | 达西定律 | 生态系统尺度 | 结合植物水力学与地形预测森林生态系统对气候变化 | [94] |
| SOX 模型 SOX model |
电类比模型 | 达西定律 | 生态系统尺度 | 模拟热带森林对干旱的反应 | [86] |
| JULES-SOX 模型 JULES-SOX model |
电类比模型 | 达西定律 | 全球尺度 | 模拟木本植物对气候变化下气孔导度响应、以及冠层水势和土壤水分动态的真实模式 | [87] |
| SurEau 模型 SurEau model |
电类比模型 | 达西定律 | 树木个体尺度 | 整合植物多个可测量的抗旱性状到一个框架中, 模拟干旱引起的死亡率 | [88] |
| SurEau-Ecos 模型 SurEau-Ecos model |
电类比模型 | 达西定律 | 生态系统尺度 | 预测从林分到区域的植物水分状况和干旱导致的死亡率 | [90] |
| CLM5-PHS 模型 CLM5-PHS model |
电类比模型 | 达西定律 | 全球尺度 | 提高热带森林对干旱的响应预测准确性 | [91] |
| Noah-MP-PHS 模型 Noah-MP-PHS model |
电类比模型 | 达西定律 | 树木个体尺度、地块级尺度 | 结合整个植物的水力策略及茎和冠层蓄水量, 评估干旱条件下植物的水分利用效率 | [92] |
| TRIPLEX-Mortality 模型 TRIPLEX-Mortality model |
/ | / | 林分尺度 | 定量模拟树木干旱死亡率 | [97] |
| Sperry: 人名;TREES: 陆地区域生态系统交换模拟器 Terrestrial regional ecosystem exchange simulator;SOX: 基于木质部水力学的气孔优化模型 Stomatal optimization model based on xylem hydraulics;JULES: 联合英国陆地环境模拟器 Joint UK land environment simulator;CLM5:社区土地模型第5版 Version 5 of the community land model;PHS: 植物水分胁迫方案 Plant hydraulic stress;Noah-MP: 具有多参数选择的社区Noah地表模型 The community Noah land surface model with multiparameterization options | |||||
明晰干旱胁迫下植物生理机制如何受环境因素调控, 可以清楚地知道植物水力特性所传递的信息。将水力特性指标与更多表征植物生理活动的结构和功能性状指标相结合, 能为与植物水力学有关的建模过程提供更多可参数化的指标。这样的综合研究使植物水力学由微观走向宏观的统一理论框架得到完善, 同时为评估和预测全球生态系统健康状况及风险提供科学依据。
未来的研究重点倾向于将植物的木质部解剖性状与光合碳获取、蒸腾水散失过程串联起来, 并融入到当下合适的模型中以简化模型参数。未来期望在类似TRIPLEX-Mortality模型的基础上实现我们的设想(表 1), 希望通过实测数据与模型模拟相结合的手段, 达到可以预测不同生态系统因干旱引起的树木死亡率对碳汇的影响程度。
| [1] |
Seleiman M F, Al-Suhaibani N, Ali N, Akmal M, Alotaibi M, Refay Y, Dindaroglu T, Abdul-Wajid H H, Battaglia M L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants, 2021, 10(2): 259. DOI:10.3390/plants10020259 |
| [2] |
Ozturk M, Turkyilmaz B, Gucel S, Guvensen A. Proline accumulation in some coastal zone plants of the Agean region of Turkey. The European Journal of Plant Science and Biotechnology, 2011, 5(SPEC.2): 54-56. |
| [3] |
Szabados L, Savouré A. Proline: a multifunctional amino acid. Trends in Plant Science, 2010, 15(2): 89-97. DOI:10.1016/j.tplants.2009.11.009 |
| [4] |
Roosens N H C J, Thu T T, Iskandar H M, Jacobs M. Isolation of the ornithine-δ-aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis thaliana. Plant Physiology, 1998, 117(1): 263-271. DOI:10.1104/pp.117.1.263 |
| [5] |
Serraj R, Sinclair T R. Osmolyte accumulation: can it really help increase crop yield under drought conditions?. Plant, Cell and Environment, 2002, 25(2): 333-341. DOI:10.1046/j.1365-3040.2002.00754.x |
| [6] |
Shabala S, Demidchik V, Shabala L, Cuin T A, Smith S J, Miller A J, Davies J M, Newman I A. Extracellular Ca2+ ameliorates NaCl-induced K+ loss from Arabidopsis root and leaf cells by controlling plasma membrane K+-permeable channels. Plant Physiology, 2006, 141(4): 1653-1665. DOI:10.1104/pp.106.082388 |
| [7] |
李雪, 王胜男, 程江珂. 干旱胁迫对植物体内钙离子含量和分布的影响研究进展. 种业导刊, 2019(12): 3-7. DOI:10.3969/j.issn.1003-4749.2019.12.001 |
| [8] |
Atif R M, Shahid L, Waqas M, Ali B, Rashid M A R, Azeem F, Nawaz M A, Wani S H, Chung G. Insights on calcium-dependent protein kinases (CPKs) signaling for abiotic stress tolerance in plants. International Journal of Molecular Sciences, 2019, 20(21): E5298. DOI:10.3390/ijms20215298 |
| [9] |
Shabala S N, Lew R R. Turgor regulation in osmotically stressed Arabidopsis epidermal root cells. direct support for the role of inorganic ion uptake as revealed by concurrent flux and cell turgor measurements. Plant Physiology, 2002, 129(1): 290-299. DOI:10.1104/pp.020005 |
| [10] |
Cui J, Tcherkez G. Potassium dependency of enzymes in plant primary metabolism. Plant Physiology and Biochemistry, 2021, 166: 522-530. DOI:10.1016/j.plaphy.2021.06.017 |
| [11] |
Egilla J N, Davies F T Jr, Drew M C. Effect of potassium on drought resistance of Hibiscus rosa-sinensis cv. Leprechaun: plant growth, leaf macro-and micronutrient content and root longevity. Plant and Soil, 2001, 229(2): 213-224. DOI:10.1023/A:1004883032383 |
| [12] |
Oddo E, Inzerillo S, Grisafi F, Sajeva M, Salleo S, Nardini A. Does short-term potassium fertilization improve recovery from drought stress in laurel?. Tree Physiology, 2014, 34(8): 906-913. DOI:10.1093/treephys/tpt120 |
| [13] |
Jia W S, Zhang J H. Stomatal movements and long-distance signaling in plants. Plant Signaling and Behavior, 2008, 3(10): 772-777. DOI:10.4161/psb.3.10.6294 |
| [14] |
Zhang F P, Sussmilch F, Nichols D S, Cardoso A A, Brodribb T J, McAdam S A M. Leaves, not roots or floral tissue, are the main site of rapid, external pressure-induced ABA biosynthesis in angiosperms. Journal of Experimental Botany, 2018, 69(5): 1261-1267. DOI:10.1093/jxb/erx480 |
| [15] |
Bauer H, Ache P, Lautner S, Fromm J, Hartung W, Al-Rasheid K A S, Sonnewald S, Sonnewald U, Kneitz S, Lachmann N, Mendel R R, Bittner F, Hetherington A M, Hedrich R. The stomatal response to reduced relative humidity requires guard cell-autonomous ABA synthesis. Current Biology, 2013, 23(1): 53-57. DOI:10.1016/j.cub.2012.11.022 |
| [16] |
Munemasa S, Hauser F, Park J, Waadt R, Brandt B, Schroeder J I. Mechanisms of abscisic acid-mediated control of stomatal aperture. Current Opinion in Plant Biology, 2015, 28: 154-162. DOI:10.1016/j.pbi.2015.10.010 |
| [17] |
Park S Y, Fung P, Nishimura N, Jensen D R, Fujii H, Zhao Y, Lumba S, Santiago J, Rodrigues A, Chow T F F, Alfred S E, Bonetta D, Finkelstein R, Provart N J, Desveaux D, Rodriguez P L, McCourt P, Zhu J K, Schroeder J I, Volkman B F, Cutler S R. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science, 2009, 324(5930): 1068-1071. DOI:10.1126/science.1173041 |
| [18] |
Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, Christmann A, Grill E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science, 2009, 324(5930): 1064-1068. DOI:10.1126/science.1172408 |
| [19] |
Wang P C, Xue L, Batelli G, Lee S, Hou Y J, Van Oosten M J, Zhang H M, Tao W A, Zhu J K. Quantitative phosphoproteomics identifies SnRK2 protein kinase substrates and reveals the effectors of abscisic acid action. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(27): 11205-11210. |
| [20] |
McAdam S A M, Brodribb T J. Linking turgor with ABA biosynthesis: implications for stomatal responses to vapor pressure deficit across land plants. Plant Physiology, 2016, 171(3): 2008-2016. DOI:10.1104/pp.16.00380 |
| [21] |
Veselov D S, Sharipova G V, Veselov S Y, Dodd I C, Ivanov I, Kudoyarova G R. Rapid changes in root HvPIP2;2 aquaporins abundance and ABA concentration are required to enhance root hydraulic conductivity and maintain leaf water potential in response to increased evaporative demand. Functional Plant Biology, 2016, 45(2): 143-149. |
| [22] |
Yaaran A, Moshelion M. Role of aquaporins in a composite model of water transport in the leaf. International Journal of Molecular Sciences, 2016, 17(7): 1045. DOI:10.3390/ijms17071045 |
| [23] |
Secchi F, Pagliarani C, Zwieniecki M A. The functional role of xylem parenchyma cells and aquaporins during recovery from severe water stress. Plant, Cell and Environment, 2017, 40(6): 858-871. DOI:10.1111/pce.12831 |
| [24] |
Zhang S, Feng M, Chen W, Zhou X F, Lu J Y, Wang Y R, Li Y H, Jiang C Z, Gan S S, Ma N, Gao J P. In rose, transcription factor PTM balances growth and drought survival via PIP2;1 aquaporin. Nature Plants, 2019, 5(3): 290-299. DOI:10.1038/s41477-019-0376-1 |
| [25] |
Daniels M J, Mirkov T E, Chrispeels M J. The plasma membrane of Arabidopsis thaliana contains a mercury-insensitive aquaporin that is a homolog of the tonoplast water channel protein TIP. Plant Physiology, 1994, 106(4): 1325-1333. DOI:10.1104/pp.106.4.1325 |
| [26] |
Postaire O, Tournaire-Roux C, Grondin A, Boursiac Y, Morillon R, Schäffner A R, Maurel C. A PIP1 aquaporin contributes to hydrostatic pressure-induced water transport in both the root and rosette of Arabidopsis. Plant Physiology, 2010, 152(3): 1418-1430. DOI:10.1104/pp.109.145326 |
| [27] |
Chaumont F, Tyerman S D. Aquaporins: highly regulated channels controlling plant water relations. Plant Physiology, 2014, 164(4): 1600-1618. DOI:10.1104/pp.113.233791 |
| [28] |
Aasamaa K, Sõber A. Seasonal courses of maximum hydraulic conductance in shoots of six temperate deciduous tree species. Functional Plant Biology, 2005, 32(12): 1077-1087. DOI:10.1071/FP05088 |
| [29] |
Savage D F, Stroud R M. Structural basis of aquaporin inhibition by mercury. Journal of Molecular Biology, 2007, 368(3): 607-617. DOI:10.1016/j.jmb.2007.02.070 |
| [30] |
Laur J, Hacke U G. The role of water channel proteins in facilitating recovery of leaf hydraulic conductance from water stress in Populus trichocarpa. PLoS One, 2014, 9(11): e111751. DOI:10.1371/journal.pone.0111751 |
| [31] |
金鹰, 王传宽. 植物叶片水力与经济性状权衡关系的研究进展. 植物生态学报, 2015, 39(10): 1021-1032. DOI:10.17521/cjpe.2015.0099 |
| [32] |
Valladares F, Wright S J, Lasso E, Kitajima K, Pearcy R W. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology, 2000, 81(7): 1925-1936. DOI:10.1890/0012-9658(2000)081[1925:PPRTLO]2.0.CO;2 |
| [33] |
Sack L, Dietrich E M, Streeter C M, Sánchez-Gómez D, Holbrook N M. Leaf palmate venation and vascular redundancy confer tolerance of hydraulic disruption. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(5): 1567-1572. |
| [34] |
Sack L, Scoffoni C. Leaf venation: structure, function, development, evolution, ecology and applications in the past, present and future. New Phytologist, 2013, 198(4): 983-1000. DOI:10.1111/nph.12253 |
| [35] |
Scoffoni C, Rawls M, McKown A, Cochard H, Sack L. Decline of leaf hydraulic conductance with dehydration: relationship to leaf size and venation architecture. Plant Physiology, 2011, 156(2): 832-843. DOI:10.1104/pp.111.173856 |
| [36] |
Brodribb T J, Feild T S. Leaf hydraulic evolution led a surge in leaf photosynthetic capacity during early angiosperm diversification. Ecology Letters, 2010, 13(2): 175-183. DOI:10.1111/j.1461-0248.2009.01410.x |
| [37] |
Salleo S, Nardini A. Sclerophylly: evolutionary advantage or mere epiphenomenon?. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology, 2000, 134(3): 247-259. DOI:10.1080/11263500012331350435 |
| [38] |
孙素静, 李芳兰, 包维楷. 叶脉网络系统的构建和系统学意义研究进展. 热带亚热带植物学报, 2015, 23(3): 353-360. |
| [39] |
刘世鹏, 刘济明, 曹娟云, 白重炎, 师荣. 干旱胁迫下枣树叶片表皮气孔分布及特征分析. 安徽农业科学, 2006, 34(7): 1315-1318. DOI:10.3969/j.issn.0517-6611.2006.07.009 |
| [40] |
孟庆辉, 潘青华, 鲁韧强, 金洪. 4个品种扶芳藤茎叶解剖结构及其与抗旱性的关系. 中国农学通报, 2006, 22(4): 138-142. DOI:10.3969/j.issn.1000-6850.2006.04.036 |
| [41] |
阮成江, 李代琼. 黄土丘陵区沙棘林几个水分生理生态特征研究. 林业科学研究, 2002, 15(1): 47-53. DOI:10.3321/j.issn:1001-1498.2002.01.008 |
| [42] |
Jordan G J, Brodribb T J, Blackman C J, Weston P H. Climate drives vein anatomy in Proteaceae. American Journal of Botany, 2013, 100(8): 1483-1493. DOI:10.3732/ajb.1200471 |
| [43] |
Berlyn G P. Plant structures: xylem structure and the ascent of sap. Science, 1983, 222(4623): 500-501. DOI:10.1126/science.222.4623.500.a |
| [44] |
李荣, 党维, 蔡靖, 张硕新, 姜在民. 6个耐旱树种木质部结构与栓塞脆弱性的关系. 植物生态学报, 2016, 40(3): 255-263. |
| [45] |
李泽东, 陈志成, 曹振, 车路平, 刘舒文, 张永涛. 华北低山丘陵区常用树种木质部解剖特征及水其力学抗旱性. 生态学报, 2021, 41(1): 69-78. DOI:10.3969/j.issn.1673-1182.2021.01.013 |
| [46] |
Skelton R P, Dawson T E, Thompson S E, Shen Y Z, Weitz A P, Ackerly D. Low vulnerability to xylem embolism in leaves and stems of North American oaks. Plant Physiology, 2018, 177(3): 1066-1077. DOI:10.1104/pp.18.00103 |
| [47] |
Choat B, Brodie T W, Cobb A R, Zwieniecki M A, Holbrook N M. Direct measurements of intervessel pit membrane hydraulic resistance in two angiosperm tree species. American Journal of Botany, 2006, 93(7): 993-1000. DOI:10.3732/ajb.93.7.993 |
| [48] |
Pittermann J, Sperry J S, Hacke U G, Wheeler J K, Sikkema E H. Torus-Margo pits help conifers compete with angiosperms. Science, 2005, 310(5756): 1924. DOI:10.1126/science.1120479 |
| [49] |
Sperry J S, Hacke U G, Wheeler J K. Comparative analysis of end wall resistivity in xylem conduits. Plant, Cell and Environment, 2005, 28(4): 456-465. DOI:10.1111/j.1365-3040.2005.01287.x |
| [50] |
Li S, Lens F, Espino S, Karimi Z, Klepsch M, Schenk H, Schmitt M, Schuldt B, Jansen S. Intervessel pit membrane thickness as a key determinant of embolism resistance in angiosperm xylem. Iawa Journal, 2016, 37(2): 152-171. DOI:10.1163/22941932-20160128 |
| [51] |
Shane M W, McCully M E, Canny M J. Architecture of branch-root junctions in maize: structure of the connecting xylem and the porosity of pit membranes. Annals of Botany, 2000, 85(5): 613-624. DOI:10.1006/anbo.2000.1113 |
| [52] |
范嘉智, 王丹, 胡亚林, 景盼盼, 王朋朋, 陈吉泉. 最优气孔行为理论和气孔导度模拟. 植物生态学报, 2016, 40(6): 631-642. |
| [53] |
Torres-Ruiz J M, Diaz-Espejo A, Perez-Martin A, Hernandez-Santana V. Role of hydraulic and chemical signals in leaves, stems and roots in the stomatal behaviour of olive trees under water stress and recovery conditions. Tree Physiology, 2015, 35(4): 415-424. DOI:10.1093/treephys/tpu055 |
| [54] |
Scoffoni C, Sack L. The causes and consequences of leaf hydraulic decline with dehydration. Journal of Experimental Botany, 2017, 68(16): 4479-4496. DOI:10.1093/jxb/erx252 |
| [55] |
Scoffoni C, Chatelet D S, Pasquet-kok J, Rawls M, Donoghue M J, Edwards E J, Sack L. Hydraulic basis for the evolution of photosynthetic productivity. Nature Plants, 2016, 2(6): 16072. DOI:10.1038/nplants.2016.72 |
| [56] |
Brodribb T J, Jordan G J. Internal coordination between hydraulics and stomatal control in leaves. Plant, Cell and Environment, 2008, 31(11): 1557-1564. DOI:10.1111/j.1365-3040.2008.01865.x |
| [57] |
Males J, Griffiths H. Economic and hydraulic divergences underpin ecological differentiation in the Bromeliaceae. Plant, Cell and Environment, 2018, 41(1): 64-78. DOI:10.1111/pce.12954 |
| [58] |
Flexas J, Diaz-Espejo A, Gago J, Gallé A, Galmés J, Gulías J, Medrano H. Photosynthetic limitations in Mediterranean plants: a review. Environmental and Experimental Botany, 2014, 103: 12-23. DOI:10.1016/j.envexpbot.2013.09.002 |
| [59] |
Sack L, Frole K. Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology, 2006, 87(2): 483-491. DOI:10.1890/05-0710 |
| [60] |
Fischer R A, Rees D, Sayre K D, Lu Z M, Condon A G, Saavedra A L. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Science, 1998, 38(6): 1467-1475. DOI:10.2135/cropsci1998.0011183X003800060011x |
| [61] |
Franks P J, Beerling D J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(25): 10343-10347. |
| [62] |
姜星星, 邹安龙, 王媛媛, 周序力, 吉成均. 我国东部典型森林木本植物的气孔特征及其对氮添加的响应. 北京大学学报: 自然科学版, 2018, 54(4): 839-847. |
| [63] |
Nardini A, Salleo S. Water stress-induced modifications of leaf hydraulic architecture in sunflower: co-ordination with gas exchange. Journal of Experimental Botany, 2005, 56(422): 3093-3101. DOI:10.1093/jxb/eri306 |
| [64] |
Brodribb T J, Jordan G J, Carpenter R J. Unified changes in cell size permit coordinated leaf evolution. New Phytologist, 2013, 199(2): 559-570. DOI:10.1111/nph.12300 |
| [65] |
Sack L, Tyree M T, Holbrook N M. Leaf hydraulic architecture correlates with regeneration irradiance in tropical rainforest trees. New Phytologist, 2005, 167(2): 403-413. DOI:10.1111/j.1469-8137.2005.01432.x |
| [66] |
Zhao W L, Chen Y J, Brodribb T J, Cao K F. Weak co-ordination between vein and stomatal densities in 105 angiosperm tree species along altitudinal gradients in Southwest China. Functional Plant Biology, 2016, 43(12): 1126-1133. DOI:10.1071/FP16012 |
| [67] |
Du Q J, Liu T, Jiao X C, Song X M, Zhang J Y, Li J M. Leaf anatomical adaptations have central roles in photosynthetic acclimation to humidity. Journal of Experimental Botany, 2019, 70(18): 4949-4962. DOI:10.1093/jxb/erz238 |
| [68] |
Choat B, Jansen S, Brodribb T J, Cochard H, Delzon S, Bhaskar R, Bucci S J, Feild T S, Gleason S M, Hacke U G, Jacobsen A L, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, Mencuccini M, Mitchell P J, Nardini A, Pittermann J, Pratt R B, Sperry J S, Westoby M, Wright I J, Zanne A E. Global convergence in the vulnerability of forests to drought. Nature, 2012, 491(7426): 752-755. DOI:10.1038/nature11688 |
| [69] |
Gleason S M, Westoby M, Jansen S, Choat B, Hacke U G, Pratt R B, Bhaskar R, Brodribb T J, Bucci S J, Cao K F, Cochard H, Delzon S, Domec J C, Fan Z X, Feild T S, Jacobsen A L, Johnson D M, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, McCulloh K A, Mencuccini M, Mitchell P J, Morris H, Nardini A, Pittermann J, Plavcová L, Schreiber S G, Sperry J S, Wright I J, Zanne A E. Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world's woody plant species. New Phytologist, 2016, 209(1): 123-136. DOI:10.1111/nph.13646 |
| [70] |
Anderegg W R L, Konings A G, Trugman A T, Yu K L, Bowling D R, Gabbitas R, Karp D S, Pacala S, Sperry J S, Sulman B N, Zenes N. Hydraulic diversity of forests regulates ecosystem resilience during drought. Nature, 2018, 561(7724): 538-541. DOI:10.1038/s41586-018-0539-7 |
| [71] |
Blackman C J. Leaf turgor loss as a predictor of plant drought response strategies. Tree Physiology, 2018, 38(5): 655-657. DOI:10.1093/treephys/tpy047 |
| [72] |
Ogasa M, Miki N H, Murakami Y, Yoshikawa K. Recovery performance in xylem hydraulic conductivity is correlated with cavitation resistance for temperate deciduous tree species. Tree Physiology, 2013, 33(4): 335-344. DOI:10.1093/treephys/tpt010 |
| [73] |
Liang X Y, Ye Q, Liu H, Brodribb T J. Wood density predicts mortality threshold for diverse trees. New Phytologist, 2021, 229(6): 3053-3057. DOI:10.1111/nph.17117 |
| [74] |
Shao J J, Zhou X H, Zhang P P, Zhai D P, Yuan T F, Li Z, He Y H, McDowell N G. Embolism resistance explains mortality and recovery of five subtropical evergreen broadleaf trees to persistent drought. Ecology, 2023, 104(2): e3877. DOI:10.1002/ecy.3877 |
| [75] |
Powers J S, Vargas G G, Brodribb T J, Schwartz N B, Pérez-Aviles D, Smith-Martin C M, Becknell J M, Aureli F, Blanco R, Calderón-Morales E, Calvo-Alvarado J C, Calvo-Obando A J, Chavarría M M, Carvajal-Vanegas D, Jiménez-Rodríguez C D, Chacon E M, Schaffner C M, Werden L K, Xu X T, Medvigy D. A catastrophic tropical drought kills hydraulically vulnerable tree species. Global Change Biology, 2020, 26(5): 3122-3133. DOI:10.1111/gcb.15037 |
| [76] |
罗丹丹, 王传宽, 金鹰. 木本植物水力系统对干旱胁迫的响应机制. 植物生态学报, 2021, 45(9): 925-941. |
| [77] |
Chen Z C, Li S, Luan J W, Zhang Y T, Zhu S D, Wan X C, Liu S R. Prediction of temperate broadleaf tree species mortality in arid limestone habitats with stomatal safety margins. Tree Physiology, 2019, 39(8): 1428-1437. DOI:10.1093/treephys/tpz045 |
| [78] |
Allen Craig D, Breshears David D, McDowell N G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere, 2015, 6(8): 1-55. |
| [79] |
Seidl R, Schelhaas M J, Rammer W, Verkerk P J. Increasing forest disturbances in Europe and their impact on carbon storage. Nature Climate Change, 2014, 4(9): 806-810. DOI:10.1038/nclimate2318 |
| [80] |
McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams D G, Yepez E A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. New Phytologist, 2008, 178(4): 719-739. DOI:10.1111/j.1469-8137.2008.02436.x |
| [81] |
Biondi F. Comparing tree-ring chronologies and repeated timber inventories as forest monitoring tools. Ecological Applications, 1999, 9(1): 216-227. DOI:10.1890/1051-0761(1999)009[0216:CTRCAR]2.0.CO;2 |
| [82] |
Marvin D C, Asner G P. Branchfall dominates annual carbon flux across lowland Amazonian forests. Environmental Research Letters, 2016, 11(9): 094027. DOI:10.1088/1748-9326/11/9/094027 |
| [83] |
Sperry J S, Adler F R, Campbell G S, Comstock J P. Limitation of plant water use by rhizosphere and xylem conductance: results from a model. Plant, Cell and Environment, 1998, 21(4): 347-359. DOI:10.1046/j.1365-3040.1998.00287.x |
| [84] |
Sperry J S, Love D M. What plant hydraulics can tell us about responses to climate-change droughts. New Phytologist, 2015, 207(1): 14-27. DOI:10.1111/nph.13354 |
| [85] |
Venturas M D, Sperry J S, Love D M, Frehner E H, Allred M G, Wang Y J, Anderegg W R L. A stomatal control model based on optimization of carbon gain versus hydraulic risk predicts aspen sapling responses to drought. New Phytologist, 2018, 220(3): 836-850. DOI:10.1111/nph.15333 |
| [86] |
Eller C B, Rowland L, Oliveira R S, Bittencourt P R L, Barros F V, Lola da Costa A C, Meir P, Friend A D, Mencuccini M, Sitch S, Cox P. Modelling tropical forest responses to drought and El Niño with a stomatal optimization model based on xylem hydraulics. Philosophical Transactions of the Royal Society B: Biological Sciences, 2018, 373(1760): 20170315. DOI:10.1098/rstb.2017.0315 |
| [87] |
Eller C B, Rowland L, Mencuccini M, Rosas T, Williams K, Harper A, Medlyn B, Wagner Y, Klein T, Teodoro G, Oliveira R, Matos I, Rosado B H P, Fuchs K, Wohlfahrt G, Montagnani L, Meir P, Sitch S, Cox P. Stomatal optimization based on xylem hydraulics (SOX) improves land surface model simulation of vegetation responses to climate. New Phytologist, 2020, 226(6): 1622-1637. DOI:10.1111/nph.16419 |
| [88] |
Martin-StPaul N, Delzon S, Cochard H. Plant resistance to drought depends on timely stomatal closure. Ecology Letters, 2017, 20(11): 1437-1447. DOI:10.1111/ele.12851 |
| [89] |
Brodribb T J, Cochard H, Dominguez C R. Measuring the pulse of trees; using the vascular system to predict tree mortality in the 21st century. Conservation Physiology, 2019, 7(1): coz046. DOI:10.1093/conphys/coz046 |
| [90] |
Ruffault J, Pimont F, Cochard H, Dupuy J L, Martin-StPaul N. SurEau-Ecos v2.0:a trait-based plant hydraulics model for simulations of plant water status and drought-induced mortality at the ecosystem level. Geoscientific Model Development, 2022, 15(14): 5593-5626. DOI:10.5194/gmd-15-5593-2022 |
| [91] |
Kennedy D, Swenson S, Oleson K W, Lawrence D M, Fisher R, Lola da Costa A C, Gentine P. Implementing plant hydraulics in the community land model, version 5. Journal of Advances in Modeling Earth Systems, 2019, 11(2): 485-513. DOI:10.1029/2018MS001500 |
| [92] |
Li L C, Yang Z L, Matheny A M, Zheng H, Swenson S C, Lawrence D M, Barlage M, Yan B Y, McDowell N G, Leung L R. Representation of plant hydraulics in the noah-MP land surface model: model development and multiscale evaluation. Journal of Advances in Modeling Earth Systems, 2021, 13(4): e2020MS002214. DOI:10.1029/2020MS002214 |
| [93] |
Yan X Y, Zhang Z D, Huang M B, Zhao X F, Yang F, Wu X F. The impact of climate change on growth and drought-induced mortality risk of Robinia pseudoacacia plantations along a precipitation gradient on the Chinese Loess Plateau. Agricultural and Forest Meteorology, 2022, 325: 109160. DOI:10.1016/j.agrformet.2022.109160 |
| [94] |
Tai X N, MacKay D S, Anderegg W R L, Sperry J S, Brooks P D. Plant hydraulics improves and topography mediates prediction of aspen mortality in southwestern USA. New Phytologist, 2017, 213(1): 113-127. DOI:10.1111/nph.14098 |
| [95] |
Tai X N, Mackay D S, Sperry J S, Brooks P, Anderegg W R L, Flanagan L B, Rood S B, Hopkinson C. Distributed plant hydraulic and hydrological modeling to understand the susceptibility of riparian woodland trees to drought-induced mortality. Water Resources Research, 2018, 54(7): 4901-4915. DOI:10.1029/2018WR022801 |
| [96] |
Sperry J S, Venturas M D, Anderegg W R L, Mencuccini M, MacKay D S, Wang Y J, Love D M. Predicting stomatal responses to the environment from the optimization of photosyn tic gain and hydraulic cost. Plant, Cell and Environment, 2017, 40(6): 816-830. DOI:10.1111/pce.12852 |
| [97] |
Liu Q Y, Peng C H, Schneider R, Cyr D, Liu Z L, Zhou X L, Kneeshaw D. TRIPLEX-Mortality model for simulating drought-induced tree mortality in boreal forests: model development and evaluation. Ecological Modelling, 2021, 455: 109652. DOI:10.1016/j.ecolmodel.2021.109652 |