 2023, Vol. 43
2023, Vol. 43文章信息
- 董琦, 王一媚, 管平婷, 宋传涛, 吴东辉, 王平
- DONG Qi, WANG Yimei, GUAN Pingting, SONG Chuantao, WU Donghui, WANG Ping
- 土壤微食物网生物和土壤微生物对小星穗薹草生长和土壤养分的影响差异
- Effects of soil micro-food web organisms and soil microorganisms on the growth of Carex angustior and soil nutrients
- 生态学报. 2023, 43(23): 9784-9793
- Acta Ecologica Sinica. 2023, 43(23): 9784-9793
- http://dx.doi.org/10.20103/j.stxb.202210012797
-
文章历史
- 收稿日期: 2022-10-01
- 网络出版日期: 2023-08-15

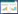




随着地下生态学的蓬勃发展, 越来越多研究揭示了地下生物对地上植物的重要性[1]。其中, 普遍认为土壤微生物能够促进植物生长[2—4]。作为分解者, 微生物参与调节土壤的多个关键生态过程[5], 如改变土壤养分的有效性[6]或促进根系生长[7]等, 从而影响植物的生物量[8]、养分含量[9], 或提高其抗逆性[6, 10]。一些植物还通过积累土壤有益微生物来促进生长, 以提高其在群落中的相对丰度[11]。但地下生物是由功能不同且关系复杂的众多土壤生物群体组成[12—13]。因此, 微生物对植物的影响可能因其他土壤生物的存在而被掩盖或放大, 如捕食微生物的土壤线虫[14]。
土壤线虫与土壤微生物之间存在极为复杂的食物链关系, 并由此形成土壤微食物网[15]。土壤微食物网是土壤生态功能的基础, 从土壤养分被分解释放到被植物吸收利用的过程中, 土壤线虫等微食物网成员发挥着重要作用, 其重要性随着地下生态学的迅速发展和研究手段的改进而逐渐呈现出来[16—17]。例如, 一些研究发现, 土壤线虫在捕食微生物过程中, 一方面持续排出超过其代谢所需要的养分物质, 使土壤有效养分含量增加[18]。另一方面线虫的捕食作用促进了微生物增殖[19], 增强了土壤呼吸强度和土壤酶活性[20]。由于线虫及其与微生物的捕食关系是土壤微食物网的构成主体[21], 因此, 土壤微食物网生物在土壤养分元素的矿化和释放[22—23]、有效养分含量提升[17]等方面的作用可能远大于土壤微生物。此外, 有研究发现土壤线虫还是植物与微生物养分竞争平衡的关键机制[14]。当土壤微生物与植物之间存在养分竞争关系时[24], 线虫对微生物的捕食作用可能有利于植物的竞争地位[25]。营养级水平较高的线虫(杂食和捕食线虫)捕食几乎所有较小的生物包括对植物有害的植食性线虫[26], 这对植物生长也会产生积极的促进作用[27]。
由于土壤微食物网生物在养分元素的转化、储存和释放, 调节植物-微生物关系等方面均产生积极影响[28], 土壤线虫及微食物网生物对植物的促进作用逐渐受到关注[29], 被认为是联系地下和地上生态过程的关键环节[14, 30], 亟待更多研究揭示其影响过程及作用机制。基于以上分析, 本研究通过设置土壤微食物网生物和土壤微生物添加处理的室内控制实验, 探究土壤微食物网生物对植物生长的促进作用是否大于土壤微生物, 以及两者对植物形态、生理特征和土壤养分含量等方面的影响是否不同。希望通过本研究, 在一定程度上推动土壤微食物网生物对植物的影响研究, 为揭示不同营养级土壤生物对植物生长和土壤养分循环的作用过程和影响机制, 以及从植物-土壤生物互作角度深入认识植物生长策略等理论研究提供新思路。
1 材料与方法 1.1 实验材料及来源本研究所用植物和土壤实验材料均来源于吉林省的金川泥炭沼泽, 位于吉林龙湾国家级自然保护区。该保护区是中国最大的火口湖群分布区, 具有特殊性和典型性。金川泥炭沼泽是在火山喷发形成火口湖的基础上, 经过漫长沼泽化过程而形成, 在我国湿地分类体系中具有独特地位[31]。金川泥炭沼泽的优势植物为薹草属植物。其中小星穗薹草(Carex angustior)在沼泽中分布广泛, 该物种也是我国北方泥炭沼泽中的常见种[32]。因此, 本研究以小星穗薹草为植物材料。植物种子于2020年7月采集, 带回东北师范大学环境学院实验室, 置于通风处阴干备用。8月以赤霉素和浓硫酸处理小星穗薹草种子促其萌发[33], 并用0.3%的高锰酸钾溶液进行消毒处理。将消毒后的种子放在带盖培养皿中, 在浸以无菌水的滤纸上萌发。3周左右后, 将萌发种子陆续移栽至生长室的育苗盘中培养, 育苗盘中土壤为高温蒸汽灭菌土壤(121℃灭菌2h, 间隔3d后再次灭菌), 采用无菌水浇灌。生长室光照12h/12h, 温度30℃/22℃, 后续植物培育均在此环境条件下进行。
2020年10月, 在采集小星穗薹草种子的相同区域采集新鲜沼泽土壤, 随即4℃保存运输至东北师范大学环境学院实验室。关于土壤线虫和微生物群落的物种组成及多样性指标详见文献[34]。按照通用方法分别提取土壤微食物网生物和土壤微生物[35]。即首先将土壤浸提液(1∶1水土比)过45μm无菌微孔滤膜, 得到土壤微食物网生物滤液(同时含有土壤微生物和线虫); 然后从该滤液中分别随机取两份, 一份过22μm无菌微孔滤膜获得土壤微生物滤液(仅含土壤微生物), 另一份灭菌(方法同土壤灭菌)作为对照处理(图 1)。

|
| 图 1 实验设计示意图 Fig. 1 Generalized diagram of experimental design |
本研究共设置土壤微食物网生物添加、微生物添加和灭菌土壤三个实验处理, 每种处理10个重复种植容器。所有处理使用相同土壤基质, 即高温蒸汽灭菌泥炭沼泽土壤。将等量灭菌土壤(约570g鲜重)装入已灭菌种植容器中(直径8cm, 高16cm的底部无孔塑料瓶), 浇入齐地表的蒸馏水静置1天待土壤稳定。随后挑选育苗盘中长势良好且个体大小基本一致的小星穗薹草幼苗移栽到容器瓶中。每瓶移栽2—3株。待1—2周移栽苗生长稳定后每瓶定株1株植物。
11月初, 分别将10mL微食物网生物浸提液、微生物浸提液或灭菌浸提液对应实验处理添加至种植容器中。在整个实验过程中均采用无菌水浇灌, 种植瓶水位始终维持在齐地表状态, 以模拟小星穗薹草的野外湿生环境。
1.3 样品采集与处理2021年3月, 即实验处理设置4个月后收获植物和土壤样品。测量每株植物的绝对高度后齐地面剪下小星穗薹草的地上部分。将植物根系与土壤仔细分离后清洗, 然后置于根盘中扫描, 获得每株植物的完整根系图片。将扫描后根系与植物地上部分放置在65℃烘箱烘干至恒重后分别称重, 计算获得总生物量和根茎比。利用WinRHIZO软件分析根系图片, 获得总根长、根系平均直径和细根长(直径<1mm)占总根长的比例等形态指标。基于总根长和根系干重计算获比根长。
使用球磨仪将烘干植物粉碎过筛后, 分别测量地上部分和根系的总氮(凯氏定氮法, 连续流动分析仪Skalar SAN ++)和总磷浓度(钼锑抗分光光度法, 连续流动分析仪Skalar SAN ++)。基于植物地上和地下生物量及总氮、总磷浓度计算获得整株植物所吸收固定的总氮量和总磷量。
种植瓶土壤风干后粉碎过筛测定土壤的全效养分和有效养分含量。以0.8mol/L重铬酸钾滴定法测定土壤有机质含量, 凯氏定氮法测定土壤总氮含量, 总有机碳分析仪法测定土壤可溶性有机碳, 碱性扩散滴定法测定土壤有效氮含量, 通过钼锑抗分光光度法测定总磷和有效磷浓度。
1.4 数据分析采用单因素方差分析法确定各个植物和土壤指标是否存在显著的处理间差异, 并以Duncan检验法进行各指标的多重比较。统计分析前检查所有数据的正态性, 对不符合正态的变量进行适当的数据转换。所有统计分析基于IBM SPSS 23.0软件, 使用Origin 2017软件制图, 图中数据均为平均值±标准误。
2 结果与分析 2.1 植物株高、生物量及根茎比不同处理之间植物株高(F=0.370, P=0.694)无显著性差异(图 2)。土壤微生物对植物生物量的促进作用未达到显著水平, 但微食物网生物显著提高了植物生物量(图 2)。与对照组相比, 土壤微食物网处理组的植物生物量增加了74.32% ±4.95%。在添加微食物网生物和微生物后植物根茎比有所下降, 但下降程度未达到显著水平(F=2.175, P=0.133, 图 2)。

|
| 图 2 三种土壤生物处理条件下植物的株高、生物量和根茎比 Fig. 2 Plant height, total biomass, and root-shoot ratio under three treatments of soil organisms 不同小写字母表示不同处理间差异显著(P<0.05),ns表示不同处理间无显著性差异 |
上述结果表明, 土壤微食物网生物对植物生物量的促进作用显著大于微生物的作用, 土壤微生物对植物生长的促进作用未达到显著性水平。在本研究中, 土壤生物未明显改变植物的株高和地上地下分配策略。
2.2 植物根系形态特征添加土壤微食物网生物后, 植物的根系总长度显著增加(图 3), 但植物的比根长、根系平均直径和细根长占比, 均与对照组植物相似(图 3)。土壤微生物对植物总根长的促进作用未达到显著水平(图 3), 却显著提高了比根长(图 3)和细根长占比(图 3), 并且显著降低了根系的平均直径(图 3)。

|
| 图 3 三种土壤生物处理条件下植物总根长、比根长、根平均直径和细根长占比 Fig. 3 Total root length, specific root length, average root diameter, and length percentage of fine roots under three treatments of soil organisms |
上述结果表明, 土壤微食物网生物和微生物对植物根系形态特征产生的影响不同。土壤微食物网生物使植物总根长显著增加, 不改变根系的形态特征, 而土壤微生物并未显著增加植物总根长, 却改变了植物的根系形态, 使根系明显纤细化。
2.3 植物氮磷浓度和总量不同处理之间无论是植物叶片(F=0.061, P=0.941)和根系(F=2.968, P=0.127)的氮浓度(图 4), 还是根系的磷浓度(F=0.395, P=0.690, 图 4)均未出现明显变化。但土壤微食物网生物显著降低了植物叶片的磷浓度(图 4)。

|
| 图 4 三种土壤生物处理条件下植物叶片和根系的总氮和总磷浓度、植物的总氮量和总磷量 Fig. 4 Total nitrogen and phosphorus concentration of plant leaves and roots, and total amount of nitrogen and phosphorus of plant under three treatments of soil organisms |
当添加土壤微食物网生物或微生物时, 植物吸收固定的总氮量增加(图 4)。但与对照组相比, 土壤微生物组的促进作用并未达到显著水平。在土壤微食物网生物处理条件下, 植物固定的总氮量显著高于对照组的(86.89±14.72)%, 并且与微生物组的差异也达到显著水平(图 4)。在本实验中, 无论添加土壤微食物网生物还是微生物, 均未明显改变植物吸收固定的总磷量(F=0.237, P=0.796, 图 4)。
上述结果表明, 土壤微食物网生物和微生物对植物氮磷浓度和氮磷元素总量的影响不同。土壤微食物网生物虽未明显改变植物的氮浓度, 却帮助小星穗薹草吸收了更多的土壤氮。土壤微生物对小星穗薹草叶片和根系的氮磷浓度, 以及吸收固定的氮磷总量影响较小。
2.4 土壤养分含量土壤微食物网生物和微生物均未显著影响土壤的有机质(F=0.379, P=0.700)、总氮(F=2.014, P=0.214)和总磷(F=1.041, P=0.409)含量(图 5)。

|
| 图 5 三种土壤生物处理条件下土壤有机质、总氮、总磷、可溶性有机碳、有效氮和有效磷含量 Fig. 5 The contents of soil organic matter, total nitrogen, total phosphorus, soluble organic carbon, available nitrogen and phosphorus under three treatments of soil organisms |
添加土壤生物后, 土壤可溶性有机碳含量增加, 并且微生物处理条件下的可溶性有机碳含量显著高于对照组(图 5)。土壤微食物网生物和微生物未显著改变土壤有效氮含量(F=0.787, P=0.497, 图 5), 但均显著降低了土壤有效磷含量(图 5)。并且微食物网生物对土壤有效磷的降低作用显著大于土壤微生物的作用。
上述结果表明, 在本实验条件下, 添加土壤食物网微生物和微生物均未改变土壤的碳氮磷全效养分含量, 但提高了土壤可溶性碳含量、降低了土壤有效磷含量。虽然双方对土壤有效养分含量的作用方向相同, 但作用程度存在差异。基本表现为, 与对照相比, 添加微食物网生物更明显降低了土壤有效磷含量, 而添加土壤微生物则明显提高了土壤可溶性碳含量。
3 讨论 3.1 土壤微食物网生物对植物生长的促进作用显著大于土壤微生物在本研究中, 土壤微生物对小星穗薹草生物量的促进作用未达到显著水平, 土壤微食物网处理则显著提高了植物生物量(图 2)。本研究及前人的一些研究[17, 36—37], 均观察到添加土壤线虫对植物生长产生的促进作用十分明显。通常情况下, 土壤微生物分解释放的养分仅有一部分能够被植物根系吸收利用, 而70%—80%的土壤碳、氮等元素则被微生物自身所固持[38]。线虫等土壤动物通过捕食微生物把这些养分释放出来, 使植物能够得以获取和利用[14]。以往一些研究中设置的微生物添加处理, 例如向灭菌土壤中添加未灭菌土壤或土壤滤液, 实质上添加的可能是包含土壤线虫在内的微食物网生物[39]。以往研究表明, 土壤线虫一方面通过捕食作用提高了微生物的代谢活性, 另一方面线虫的分泌物及排泄物也为微生物提供了养分[17]。因此, 由相互作用的线虫和微生物构成的土壤微食物网, 可加快土壤碳氮元素周转[40], 从而提高植物对土壤养分的利用率[41]。在本研究的土壤微食物网生物添加处理中, 小星穗薹草的植物总氮量显著高于其他两个处理(图 4)。这证明土壤微食物网生物帮助植物吸收了更多的土壤可利用养分资源[18, 42], 最终促进植物生长[43—44]。
此外, 土壤线虫及其他原生动物能够通过激素效应刺激植物生长, 如促进植物根尖和根系生长[45]、增加根系分枝等[40], 这也是微食物网生物促进植物生长的原因之一。添加土壤微食物网生物后, 小星穗薹草的总根长明显大于对照组植物(图 3), 植物占有和吸收土壤养分的能力显著增加。除促进根系发育外, 土壤线虫还帮助有益微生物(如硝化细菌)到达植物的新生根尖处[46], 进一步增强植物对土壤养分的利用能力。
与土壤微食物网生物对植物根系产生的作用不同, 土壤微生物并未显著提高小星穗薹草的总根长, 却改变了根系的生长策略和形态。与其他处理相比, 土壤微生物使小星穗薹草倾向于采取消耗等量干物质形成更长根系的生长策略(比根长增大, 图 3), 从而使整个根系的外部形态更显纤细(根系平均直径减小, 细根所占长度比例增加, 图 3)。植物具有改变生长策略, 调整其形态和结构以提高环境适合度的能力[47—48]。小星穗薹草根系发生的上述生长策略和形态变化与植物在低养分环境中的表现相似[49]。这或许是因为在本研究的湿生环境中, 土壤微生物对土壤养分的矿化能力受到限制, 其分解释放的养分无法充分满足微生物和植物的共同需求, 从而出现某种程度的养分竞争, 使得小星穗薹草对根系的投资策略发生了适应性改变。前人研究发现线虫对植物生长的促进作用通常发生在微生物与植物竞争养分的环境中[36], 未来应开展相关研究以提供更多实验证据。
3.2 土壤微食物网生物和微生物对土壤有效养分含量的影响不同植物根系吸收和获取的营养资源与土壤微食物网生物活动密切相关[16]。小星穗薹草需不断从土壤中获取可利用营养以满足生长所需。以往研究表明, 土壤生物影响土壤养分的可利用性, 继而影响植物对营养元素的吸收[16, 18, 43]。一般而言, 土壤的全效养分含量反映土壤养分的储量, 而有效养分含量反映土壤矿化与植物吸收之间的动态平衡[50]。在本次四个月的实验周期中, 短期的实验处理并未明显改变土壤的碳、氮、磷全效养分含量(图 5)。但在土壤生物和小星穗薹草的共同作用下, 土壤有效养分含量发生差异性变化, 表明有无、以及不同土壤生物对土壤矿化与植物吸收的动态平衡产生了不同影响。
土壤微生物的代谢活动促进了土壤可溶性有机碳的释放[51], 使土壤可溶性有机碳含量明显升高(图 5)。因土壤微生物直接参与有机物分解, 土壤碳循环易受到微生物的影响[52]。土壤线虫也可促进土壤可溶性有机碳的释放[53], 特别是食微线虫(食细菌线虫和食真菌线虫)。但线虫从微生物中获取得的大部分碳被各营养级线虫(包括高营养级捕食性线虫和杂食性线虫)呼吸代谢消耗[54]。这可能是导致微食物网处理的土壤可溶性有机碳含量略低于微生物处理的原因之一(图 5)。
土壤有效磷含量在不同处理间也差异明显。添加土壤生物后, 土壤的有效磷含量显著下降, 而且微食物网生物对土壤有效磷的降低作用大于土壤微生物(图 5)。土壤生物有利于土壤养分的矿化[55], 但矿化后的养分元素可能很快又被植物或地下生物吸收转化而固定, 土壤的有效养分含量未必提高, 甚至可能因土壤生物的固持反而降低。如以往研究发现, 细菌重新固持已分解释放的磷素[56], 并且当土壤线虫存在时这种磷固持现象更明显[17, 57]。这是因为线虫的碳磷比(C/P)值与微生物接近, 线虫取食微生物后很少释放有效磷, 从而导致土壤有效磷含量较低[58]。
4 结论土壤生物与植物之间相互依存, 但不同土壤生物类群对植物生长产生的影响各不相同。本研究发现, 土壤微食物网生物对植物生长的促进作用大于土壤微生物的作用。土壤微食物网生物未改变植物的氮浓度和土壤有效氮含量, 却促进了植物对土壤氮元素的吸收, 使植物的总氮量显著增加。土壤微食物网生物和土壤微生物在植物形态、生理特征和土壤养分含量等方面存在的差异性影响结果, 可能是因为土壤微食物网生物比土壤微生物更加快了土壤养分的释放和周转, 使植物能够从土壤中吸收更多可利用养分, 相关生态过程和作用机制研究亟待深入开展。此外, 应构建野外环境条件下合理可行的实验方法和途径, 以确定室内实验的研究结论是否与野外环境中的一致。未来在开展相关研究工作时, 还应考虑到土壤线虫与土壤微生物、以及不同营养级线虫和不同种类微生物之间的复杂关系。其中的一些重要关系, 以及这些关系与土壤养分的反馈作用, 可能是揭示土壤微食物网生物对植物生长的影响过程和作用机制的关键枢纽。
| [1] |
van Geem M, Gols R, van Dam N M, van der Putten W H, Fortuna T, Harvey J A. The importance of aboveground-belowground interactions on the evolution and maintenance of variation in plant defense traits. Frontiers in Plant Science, 2013, 4: 431. |
| [2] |
Wei Z, Gu Y A, Friman V P, Kowalchuk G A, Xu Y C, Shen Q R, Jousset A. Initial soil microbiome composition and functioning predetermine future plant health. Science Advances, 2019, 5(9): eaaw0759. DOI:10.1126/sciadv.aaw0759 |
| [3] |
Trivedi P, Leach J E, Tringe S G, Sa T M, Singh B K. Plant-microbiome interactions: from community assembly to plant health. Nature Reviews Microbiology, 2020, 18(11): 607-621. DOI:10.1038/s41579-020-0412-1 |
| [4] |
Li E Q, de Jonge R, Liu C, Jiang H N, Friman V P, Pieterse C M J, Bakker P A H M, Jousset A. Rapid evolution of bacterial mutualism in the plant rhizosphere. Nature Communications, 2021, 12(1): 1-13. DOI:10.1038/s41467-020-20314-w |
| [5] |
van der Putten W H, Bardgett R D, Bever J D, Bezemer T M, Casper B B, Fukami T, Kardol P, Klironomos J N, Kulmatiski A, Schweitzer J A, Suding K N, Van de Voorde T F J, Wardle D A. Plant-soil feedbacks: the past, the present and future challenges. Journal of Ecology, 2013, 101(2): 265-276. DOI:10.1111/1365-2745.12054 |
| [6] |
金媛媛, SAMAN Bowatte, 田沛, 彭泽晨, 侯扶江, 李春杰. 禾草-内生真菌共生对土壤理化性质及其微生物影响的研究进展. 草业科学, 2019, 36(5): 1292-1307. |
| [7] |
Verbon E H, Liberman L M. Beneficial microbes affect endogenous mechanisms controlling root development. Trends in Plant Science, 2016, 21(3): 218-229. DOI:10.1016/j.tplants.2016.01.013 |
| [8] |
Wang X Y, Ge Y, Wang J. Positive effects of plant diversity on soil microbial biomass and activity are associated with more root biomass production. Journal of Plant Interactions, 2017, 12(1): 533-541. DOI:10.1080/17429145.2017.1400123 |
| [9] |
Oldroyd G E D, Leyser O. A plant's diet, surviving in a variable nutrient environment. Science, 2020, 368(6486): eaba0196. DOI:10.1126/science.aba0196 |
| [10] |
de Vries F T, Griffiths R I, Knight C G, Nicolitch O, Williams A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science, 2020, 368(6488): 270-274. DOI:10.1126/science.aaz5192 |
| [11] |
Oliverio A M, Geisen S, Delgado-Baquerizo M, Maestre F T, Turner B L, Fierer N. The global-scale distributions of soil protists and their contributions to belowground systems. Science Advances, 2020, 6(4): eaax8787. DOI:10.1126/sciadv.aax8787 |
| [12] |
Mickael H, Bartz Marie L C, Andrey Z. The effects of soil fauna on soil functioning through the prism of traits. Geoderma, 2023, 429: 116244. DOI:10.1016/j.geoderma.2022.116244 |
| [13] |
Du X F, Liu H W, Li Y B, Li B, Han X, Li Y H, Mahamood M, Li Q. Soil community richness and composition jointly influence the multifunctionality of soil along the forest-steppe ecotone. Ecological Indicators, 2022, 139: 108900. DOI:10.1016/j.ecolind.2022.108900 |
| [14] |
Wilschut R A, Geisen S. Nematodes as drivers of plant performance in natural systems. Trends in Plant Science, 2021, 26(3): 237-247. DOI:10.1016/j.tplants.2020.10.006 |
| [15] |
Bonkowski M, Villenave C, Griffiths B. Rhizosphere fauna: the functional and structural diversity of intimate interactions of soil fauna with plant roots. Plant and Soil, 2009, 321(1): 213-233. |
| [16] |
Chen D M, Xing W, Lan Z C, Saleem M, Wu Y, Hu S J, Bai Y F. Direct and indirect effects of nitrogen enrichment on soil organisms and carbon and nitrogen mineralization in a semi-arid grassland. Functional Ecology, 2019, 33(1): 175-187. DOI:10.1111/1365-2435.13226 |
| [17] |
陈云峰, 曹志平. 土壤食物网: 结构、能流及稳定性. 生态学报, 2008, 28(10): 5055-5064. |
| [18] |
Zhang S X, Chang L, McLaughlin N, Cui S Y, Wu H T, Wu D H, Liang W J, Liang A Z. Complex soil food web enhances the association between N mineralization and soybean yield-a model study from long-term application of a conservation tillage system in a black soil of Northeast China. SOIL, 2021, 7(1): 71-82. DOI:10.5194/soil-7-71-2021 |
| [19] |
Jiang Y J, Liu M Q, Zhang J B, Chen Y, Chen X Y, Chen L J, Li H X, Zhang X X, Sun B. Nematode grazing promotes bacterial community dynamics in soil at the aggregate level. The ISME Journal, 2017, 11(12): 2705-2717. DOI:10.1038/ismej.2017.120 |
| [20] |
毛小芳, 李辉信, 龙梅, 胡锋. 不同食细菌线虫取食密度下线虫对细菌数量、活性及土壤氮素矿化的影响. 应用生态学报, 2005, 16(6): 1112-1116. |
| [21] |
杜晓芳, 李英滨, 刘芳, 宿晓琳, 李琪. 土壤微食物网结构与生态功能. 应用生态学报, 2018, 29(2): 403-411. |
| [22] |
Osler G H R, Sommerkorn M. Toward a complete soil c and n cycle: incorporating the soil fauna. Ecology, 2007, 88(7): 1611-1621. DOI:10.1890/06-1357.1 |
| [23] |
Zhang P, Zhang W J, Hu S J. Fungivorous nematode Aphelenchus avenae and Collembola Hypogastrura perplexa alleviate damping-off disease caused by Pythium ultimum in tomato. Plant and Soil, 2023, 482(1): 175-189. |
| [24] |
Kaštovská E, Šantrčková H. Comparison of uptake of different N forms by soil microorganisms and two wet-grassland plants: a pot study. Soil Biology and Biochemistry, 2011, 43(6): 1285-1291. DOI:10.1016/j.soilbio.2011.02.021 |
| [25] |
De Deyn G B, Raaijmakers C E, Van Ruijven J, Berendse F, Van Der Putten W H. Plant species identity and diversity effects on different trophic levels of nematodes in the soil food web. Oikos, 2004, 106(3): 576-586. DOI:10.1111/j.0030-1299.2004.13265.x |
| [26] |
Khan Z, Kim Y H. A review on the role of predatory soil nematodes in the biological control of plant parasitic nematodes. Applied Soil Ecology, 2007, 35(2): 370-379. DOI:10.1016/j.apsoil.2006.07.007 |
| [27] |
Dietrich P, Roeder A, Cesarz S, Eisenhauer N, Ebeling A, Schmid B, Schulze E D, Wagg C, Weigelt A, Roscher C. Nematode communities, plant nutrient economy and life-cycle characteristics jointly determine plant monoculture performance over 12 years. Oikos, 2020, 129(4): 466-479. DOI:10.1111/oik.06989 |
| [28] |
Shankar B S, Karolina F. Soil-plant-microbe interactions determine soil biological fertility by altering rhizospheric nutrient cycling and biocrust formation. Sustainability, 2022, 15(1): 625. DOI:10.3390/su15010625 |
| [29] |
Sackett T E, Classen A T, Sanders N J. Linking soil food web structure to above- and belowground ecosystem processes: a meta-analysis. Oikos, 2010, 119(12): 1984-1992. DOI:10.1111/j.1600-0706.2010.18728.x |
| [30] |
Bardgett R D, van der Putten W H. Belowground biodiversity and ecosystem functioning. Nature, 2014, 515(7528): 505-511. DOI:10.1038/nature13855 |
| [31] |
盛连喜. 吉林龙湾国家级自然保护区综合科学考察报告. 长春: 吉林科学技术出版社, 2013.
|
| [32] |
吴征镒. 中国植物志(第十二卷). 北京: 科学出版社, 2000: 514.
|
| [33] |
袁继红, 王银柳, 林友兴, 盛连喜, 王平. 酸和赤霉素处理对3种薹草种子萌发特征的影响. 东北师大学报: 自然科学版, 2016, 48(2): 112-117. |
| [34] |
王一媚. 土壤微食物网对湿地植物种间竞争的影响研究[D]. 长春: 东北师范大学, 2022.
|
| [35] |
Li Y B, Veen G F, Gera Hol W H, Vandenbrande S, Hannula S E, ten Hooven F C, Li Q, Liang W J, Bezemer T M. 'Home' and 'away' litter decomposition depends on the size fractions of the soil biotic community. Soil Biology and Biochemistry, 2020, 144: 107783. DOI:10.1016/j.soilbio.2020.107783 |
| [36] |
Matus-Acuña V, Caballero-Flores G, Reyes-Hernandez B J, Martínez-Romero E. Bacterial preys and commensals condition the effects of bacteriovorus nematodes on Zea mays and Arabidopsis thaliana. Applied Soil Ecology, 2018, 132: 99-106. DOI:10.1016/j.apsoil.2018.08.012 |
| [37] |
Kou X C, Su T Q, Ma N N, Li Q, Wang P, Wu Z F, Liang W J, Cheng W X. Soil micro-food web interactions and rhizosphere priming effect. Plant and Soil, 2018, 432(1): 129-142. |
| [38] |
Garcia A B, Locke H, Crawford K M. Soil microbes weaken the positive effect of an aquatic-terrestrial subsidy on plant performance. Journal of Plant Ecology, 2021, 14(1): 108-116. DOI:10.1093/jpe/rtaa079 |
| [39] |
Petipas R H, Wruck A C, Geber M A. Microbe-mediated local adaptation to limestone barrens is context dependent. Ecology, 2020, 101(8): e03092. DOI:10.1002/ecy.3092 |
| [40] |
Trap J, Bonkowski M, Plassard C, Villenave C, Blanchart E. Ecological importance of soil bacterivores for ecosystem functions. Plant and Soil, 2016, 398(1): 1-24. |
| [41] |
Djigal D, Brauman A, Diop T A, Chotte J L, Villenave C. Influence of bacterial-feeding nematodes (Cephalobidae) on soil microbial communities during maize growth. Soil Biology and Biochemistry, 2004, 36(2): 323-331. DOI:10.1016/j.soilbio.2003.10.007 |
| [42] |
Haase S, Ruess L, Neumann G, Marhan S, Kandeler E. Low-level herbivory by root-knot nematodes (Meloidogyne incognita) modifies root hair morphology and rhizodeposition in host plants (Hordeum vulgare). Plant and Soil, 2007, 301(1): 151-164. |
| [43] |
Jiang Y, Wang Z H, Liu Y, Han Y L, Wang Y, Wang Q, Liu T. Nematodes and their bacterial prey improve phosphorus acquisition by wheat. New Phytologist, 2023, 237(3): 974-986. DOI:10.1111/nph.18569 |
| [44] |
Jiang Y, Wu Y, Hu N, Li H X, Jiao J G. Interactions of bacterial-feeding nematodes and indole-3-acetic acid (IAA)-producing bacteria promotes growth of Arabidopsis thaliana by regulating soil auxin status. Applied Soil Ecology, 2020, 147: 103447. DOI:10.1016/j.apsoil.2019.103447 |
| [45] |
Mao X F, Hu F, Griffiths B, Li H X. Bacterial-feeding nematodes enhance root growth of tomato seedlings. Soil Biology and Biochemistry, 2006, 38(7): 1615-1622. |
| [46] |
Knox O G G, Killham K, Artz R R E, Mullins C, Wilson M. Effect of nematodes on rhizosphere colonization by seed-applied bacteria. Applied and Environmental Microbiology, 2004, 70(8): 4666-4671. DOI:10.1128/AEM.70.8.4666-4671.2004 |
| [47] |
Postma J A, Schurr U, Fiorani F. Dynamic root growth and architecture responses to limiting nutrient availability: linking physiological models and experimentation. Biotechnology Advances, 2014, 32(1): 53-65. |
| [48] |
de Lima C F F, Kleine-Vehn J, De Smet I, Feraru E. Getting to the root of belowground high temperature responses in plants. Journal of Experimental Botany, 2021, 72(21): 7404-7413. |
| [49] |
Gruber B D, Giehl R F H, Friedel S, von Wirén N. Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiology, 2013, 163(1): 161-179. |
| [50] |
Wang L, Yang F, Yaoyao E, Yuan J, Raza W, Huang Q W, Shen Q R. Long-term application of bioorganic fertilizers improved soil biochemical properties and microbial communities of an apple orchard soil. Frontiers in Microbiology, 2016, 7: 1893. |
| [51] |
Shi L, Dech J P, Yao H X, Zhao P W, Shu Y, Zhou M. The effects of nitrogen addition on dissolved carbon in boreal forest soils of northeastern China. Scientific Reports, 2019, 9(1): 1-9. |
| [52] |
Maron P A, Sarr A, Kaisermann A, Lévêque J, Mathieu O, Guigue J, Karimi B, Bernard L, Dequiedt S, Terrat S, Chabbi A, Ranjard L. High microbial diversity promotes soil ecosystem functioning. Applied and Environmental Microbiology, 2018, 84(9): e02738-e02717. |
| [53] |
罗天相, 李辉信, 王同, 胡锋. 线虫和蚯蚓对土壤微量气体排放的影响. 生态学报, 2008, 28(3): 993-999. |
| [54] |
Griffiths B S. Microbial-feeding nematodes and protozoa in soil: their effectson microbial activity and nitrogen mineralization in decomposition hotspots and the rhizosphere. Plant and Soil, 1994, 164(1): 25-33. |
| [55] |
Li Z L, Tian D S, Wang B X, Wang J S, Wang S, Chen H Y H, Xu X F, Wang C H, He N P, Niu S L. Microbes drive global soil nitrogen mineralization and availability. Global Change Biology, 2019, 25(3): 1078-1088. |
| [56] |
Savin M C, Görres J H, Neher D A, Amador J A. Uncoupling of carbon and nitrogen mineralization: role of microbivorous nematodes. Soil Biology and Biochemistry, 2001, 33(11): 1463-1472. |
| [57] |
Zheng J, Dini-Andreote F, Luan L, Geisen S, Xue J R, Li H X, Sun B, Jiang Y J. Nematode predation and competitive interactions affect microbe-mediated phosphorus dynamics. mBio, 2022, 13(3): e0329321. |
| [58] |
胡锋, 李辉信, 谢涟琪, 吴珊眉. 土壤食细菌线虫与细菌的相互作用及其对N、P矿化生物固定的影响及机理. 生态学报, 1999, 19(6): 914-920. |