 2023, Vol. 43
2023, Vol. 43文章信息
- 黄松, 李家湘, 谢勇, 刘文倩, 龚群龙, 邓梦达, 黄明增, 赵茉涵
- HUANG Song, LI Jiaxiang, XIE Yong, LIU Wenqian, GONG Qunlong, DENG Mengda, Huang Mingzeng, ZHAO Mohan
- 世界古老果树榅桲在中国的归化及其种群动态——以湖南省褒忠山种群为例
- Naturalization of the ancient fruit tree Cydonia oblonga and its population dynamics in China: A case study of Baozhong Mountain population in Hunan Province
- 生态学报. 2023, 43(21): 8840-8851
- Acta Ecologica Sinica. 2023, 43(21): 8840-8851
- http://dx.doi.org/10.20103/j.stxb.202210122899
-
文章历史
- 收稿日期: 2022-10-12
- 网络出版日期: 2023-06-26





2. 中南林业科技大学家具与艺术设计学院, 长沙 410004;
3. 国家林业和草原局中南调查规划院, 长沙 410014;
4. 湘潭市林业局, 湘潭 411199
2. College of Furniture and Art Design, Central South University of Forestry & Technology, Changsha 410004, China;
3. Central South Inventory and Planning Institute of National Forestry and Grassland Administration, Changsha 410014, China;
4. Forestry Administration of Xiangtan Municipality, Xiangtan 411199, China
归化植物是指在自然状态下可自行繁衍, 且能够长期维持种群(一般为10年以上)的本土之外的外来植物[1—2], 当归化植物对本地的生态和经济造成损害时则构成了入侵。外来植物的归化过程是其形成入侵的前提, 详细清晰的归化植物本底资料是防治外来植物入侵的重要基础, 21世纪以来越来越多的国家或地区制定了本土归化植物清单, 如印度、巴西和加纳等分别报道了本土共有787种[3]、525种[4]、291[5]种归化植物。中国的归化植物于不同的名录中分别记载有420种[6]、861种[7]、933种[8]和1099种[9], 呈现出物种数随着时间的推移和研究的深入而不断增加的趋势。但在部分名录中也存在着将引进栽培物种和国产物种视为归化物种、已经归化的物种未能收录等问题[10], 导致名录缺乏足够的科学性与说服力, 其原因在于对外来物种是否归化的判断缺乏科学规范的调查研究方法。
目前, 中国对于归化植物的报道主要基于外来新记录种的植物分类依据, 对于其群落学调查内容多限于简要概述[11—13], 因而其数量规模、生境特征、自我繁衍和种群维持能力等是否完全符合归化植物的定性标准仍有待商榷[14]。国际上关于如何判断外来引种植物的归化状态的研究多局限于草本植物[15—17], 对外来木本植物多基于自我繁殖能力、物候规律以及生境适应性等特性来判断其归化状态[18—20], 鲜有从群落状况和种群动态的角度对逸生木本植物种群的发展状况进行分析。种群动态是种群生态学研究的重要内容, 能反映现有种群个体数量的分布和配置现状, 体现种群的数量动态及未来演变趋势[21]。较草本植物而言, 外来木本植物生命周期长、更新世代慢, 从引种、逸生再到归化需要更长的时间和更广的生境, 对其归化状态进行确定更为艰难, 易出现偏差。因此对外来木本植物的种群动态进行研究将有助于对其归化状态及潜在入侵性进行客观、科学的判断。
榅桲(Cydonia oblonga)为蔷薇科(Rosaceae)榅桲属(Cydonia)木本植物, 起源于高加索地区, 以伊朗北部、土耳其、阿塞拜疆、俄罗斯西南部等地区为多样性中心, 在药用、食用加工和果树栽植等方面均具有重要经济价值, 已被引种于美国、墨西哥、印度、日本、突尼斯等多个国家[22]。榅桲在中国的引种可追溯至两汉至南北朝时期, 距今已有2500年[23], 现主要生长于新疆、陕西、山东、福建等省份, 且均处于人工栽植状态下[24], 未见榅桲在中国逸生或归化的报道和研究, 在已有的中国归化植物名录中也未将榅桲列入[8—9]。本研究首次记录了逸生于湖南省褒忠山的榅桲种群, 基于严靖等[25]提出的归化植物广义的定性标准, 通过群落物种组成、区系特征和垂直结构对榅桲种群优势地位和生境适应性进行分析, 结合静态生命表、生存分析和时间序列预测等方法对其种群动态进行描述, 从而对榅桲的归化状态进行判断, 结果可为中国归化植物名录的完善和外来木本植物归化状况的判断方法提供参考。
1 方法 1.1 研究区概况研究区位于湖南省湘潭市褒忠山国有林场(27°48′24″—27°52′35″N, 112°12′30″—112°12′52″E), 为湘中丘陵区孤山地貌, 坡度在30—45°之间, 最高海拔802 m, 最低海拔185 m, 最大高差617 m。成土母岩主要为板页岩和砂岩, 土壤垂直分异明显: 400 m以下为红壤, 400—500 m为黄红壤, 600 m以上为黄壤, 土层厚度平均为40—80 cm, 土壤呈酸性至微酸性。气候属中亚热带湿润气候, 年均温15℃, 年降水量1240—1360 mm, 无霜期262—275 d。在植被区划上为中亚热带常绿阔叶林北部植被亚地带, 属于湘中、湘东植被区下的长、潭、株丘陵植被小区[26], 地带性植被为中亚热带常绿阔叶林, 因长期人为干扰, 原生植被破坏严重, 现存植被以樟树(Cinnamomum camphora)林、杉木(Cunninghamia lanceolata)林、湿地松(Pinus elliottii)林和毛竹(Phyllostachys edulis)林为主, 仅在名胜古迹附近保留有小面积老龄林, 建群种以甜槠(Castanopsis eyrei)、苦槠(Castanopsis sclerophylla)、小叶白辛树(Pterostyrax corymbosus)等为优势。榅桲群落分布于褒忠山顶峰, 海拔750 m以上, 平均坡度大于30°, 伴生小乔木有冬青(Ilex chinensis)、白背叶(Mallotus apelta)等, 灌木有中华绣线菊(Spiraea chinensis)、杜鹃(Rhododendron simsii)、山莓(Rubus corchorifolius)等, 林下草本主要有褐果薹草(Carex brunnea)、阔鳞鳞毛蕨(Dryopteris championii)、三脉紫菀(Aster ageratoides)、芒(Miscanthus sinensis)等。
1.2 调查方法于2021年7月对榅桲群落进行样地调查, 设置20个5 m × 5 m的灌木层监测样方, 每个灌木层样方沿斜对角线在两角和中心共设置3个1 m × 1 m的草本层样方[27]。以灌木层样方为单位, 将离地5 cm以下分枝的萌条视作独立个体[28], 测记所有灌木个体的种类、基径(Basal diameter, BD, 离地5 cm处树干直径)、树高(Height, H)、冠幅[29]。同时记录草本层样方内的种类、多度、盖度、平均高度等[30], 对调查范围内草本层样方外的植物仅记录种类。
1.3 数据分析 1.3.1 群落数据分析(1) 群落植物区系。参照吴征镒[31]划分的中国种子植物属的分布区类型, 对样方内种子植物属的区系成分进行统计分析。
(2) 物种重要值。参考方精云[30]计算灌丛样方各层物种重要值的方法: 灌木层植物重要值=(相对密度+相对显著度+相对盖度)/3×100%, 其中相对显著度=(该物种的基面积/所有灌木的基面积)×100%;草本层植物重要值=(相对盖度+相对多度+相对频度)/3×100%。
(3) 高度级划分。仅对灌木层进行统计分析, 高度级以0.5 m为递增单位, 共划分成H<0.5 m, 0.5 m≤H<1 m, …, 3.5 m≤H<4 m共8个级别。
1.3.2 种群龄级划分采用时空替代的方法, 以径级代替龄级的划分标准来对种群动态变化规律进行分析已得到许多学者的认可[32]。根据样地调查数据和种群状况, 将其划分为8个龄级: Ⅰ级(BD<2 cm)、Ⅱ级(2 cm≤BD<4 cm)、Ⅲ级(4 cm≤BD<6 cm)、......、Ⅷ级(14 cm≤BD<16 cm)。根据榅桲生长发育特点, 8个龄级可以归为5个生长发育阶段: 幼苗(Ⅰ龄级)、小树(Ⅱ—Ⅲ龄级)、中树(Ⅳ—Ⅴ龄级)、大树(Ⅵ—Ⅶ龄级)、老树(Ⅷ龄级)。
1.3.3 种群年龄结构及动态参考陈晓德[33]分析种群结构动态量化的方法, 具体公式如下:
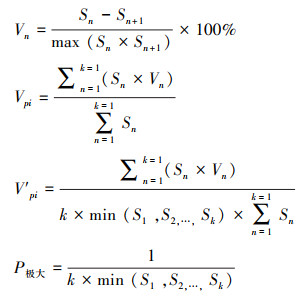
|
式中, Vn为种群从n到n+1级的个体数量变化; Vpi为忽略外部干扰情况下种群结构数量变化动态指数; V′pi为在考虑外部干扰情况下种群结构数量变化动态指数; Sn为第n级种群个体数; Sn+1为第n+1级种群个体数; k为种群年龄级数量。Vn、Vpi、V′pi取正、零、负值时分别反映种群个体数量的增长、稳定和衰退的结构动态关系。当P为最大值时, 才会对种群动态V′pi构成最大影响。
1.3.4 种群静态生命表和存活曲线种群静态生命表是利用特定时期内所有个体数据编制而成, 能够反映出种群年龄动态过程中的特定时间, 是解释种群变化的前提[6]。静态生命表编制公式如下[34]: lx=(Ax/A0)×1000; dx=lx-lx+1; qx=Ax/lx×100%; Lx=(lx+lx+1)/2;Tx=Lx+Lx+1+Lx+2+…+Lx+n; ex=Tx/lx; Kx=lnlx-lx+1; Sx=lx+1/lx。式中, Ax为x龄级内的每公顷存活个体数; lx为第x龄级开始时标准化存活个体数; dx为从x到x+1龄级期间的标准化死亡数; qx为第x到x+1龄级间隔期间死亡率; Lx为第x到x+1龄级间隔期间仍存活的个体数; Tx为大于或等于x径级的存活个体总数; ex为进入第x径级个体的生命期望或平均期望寿命; Kx为消失率; Sx为存活率。
存活曲线与种群的更新能力密切相关, 常被用于描述特定年龄的死亡率[6]。依据Helt和Loucks提出指数函数Nx=N0e-bx和幂函数Nx=N0x-b对标准化存活对数lnx和龄级关系进行拟合, 依据决定系数R2和F检验值确定最优模型, 根据两个函数的拟合效果来判断榅桲种群存活状况符合哪种类型曲线(Deevey-Ⅱ型和Deevey-Ⅲ型)[35]。
1.3.5 种群生存分析种群生存曲线基于生存分析中的4个函数(生存函数S(i)、累计死亡率函数F(i)、死亡密度函数f(ti)和危险率函数λ(ti)), 对榅桲种群动态变化情况进行分析, 4个函数的表达式如下[32]:
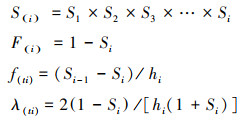
|
式中, Si为存活率; hi为径级宽度。
1.3.6 种群数量动态预测时间序列分析是研究种群未来年龄结构发展趋势的重要手段。采用一次移动平均法预测榅桲种群年龄结构, 其计算公式为[21]:
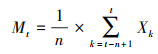
|
式中, n为预测时间, t为龄级, Xk为k龄级内的种群个体数量, Mt为n个龄级后t龄级的种群个体数量。
2 结果 2.1 群落结构 2.1.1 物种组成与区系分析榅桲群落共记录维管束植物20科28属31种, 包含蕨类植物3科3属3种, 种子植物17科25属28种。属、种最具优势的科为蔷薇科(Rosaceae)(3属6种)、菊科(Compositae)(4属4种)。木本植物16种, 占总种数的51.61%, 其中落叶木本植物13种, 占木本植物总数的81.25%, 常绿木本植物仅3种, 分别为冬青(Ilex chinensis)、小果蔷薇(Rosa cymosa)、菝葜(Smilax china)。乔木树种共4种, 分别为榅桲、冬青、白背叶和茅栗(Castanea seguinii), 但皆成灌木状。草本植物15种, 占总种数的48.39%。该群落区系地理成分复杂, 25个种子植物属可划分为9个分布区类型(表 1)。热带成分(2, 6, 7分布型)共5属, 占总属数(未包括世界分布属)的25%。温带成分(8, 9, 10, 12, 14分布型)共15属, 占总属数的75%, 其中以北温带分布(8分布型)为主, 占总属数的28%。
| 分布区类型Areal-type | 属数No. of genera | 比例/% |
| 1世界分布Cosmopolitan | 5 | 20 |
| 2泛热带分布Pantropic | 3 | 12 |
| 6热带亚洲至热带非洲分布Tropical Asia to Tropical Africa | 1 | 4 |
| 7热带亚洲分布Tropical Asia | 1 | 4 |
| 8北温带分布North Temperate | 7 | 28 |
| 9东亚和北美洲间断分布East Asia and North America disjuncted | 1 | 4 |
| 10旧世界温带分布Old World Temperate | 3 | 12 |
| 12地中海区、西亚至中亚分布Mediterranean, West Asia to Central Asia | 1 | 4 |
| 14东亚分布East Asia | 3 | 12 |
| 总计Total | 25 | 100 |
榅桲群落可以明显地分为灌木层和草本层, 灌木层平均高0.94 m, 草本层平均高0.26 m, 各层优势种明显(图 1)。灌木层盖度约85%, 以榅桲为优势种(平均高1.73 m, 重要值达67.12%), 其次为中华绣线菊、杜鹃、白马骨(Serissa serissoides)、山莓等(表 2)。草本层盖度约为30%, 以褐果薹草为主要优势种, 重要值达49.49%, 其次为阔鳞鳞毛蕨、三脉紫菀等(表 3)。

|
| 图 1 褒忠山榅桲群落 Fig. 1 Cydonia oblonga community in Baozhong mountain |
| 物种 Species |
个体数 No. of individuals |
平均高 Mean H/m |
相对显著度 RS/% |
相对密度 RD/% |
相对盖度 RC/% |
重要值 IV/% |
| 榅桲Cydonia oblonga Mill. | 385 | 1.73 | 96.26 | 28.00 | 77.11 | 67.12 |
| 中华绣线菊Spiraea chinensis Maxim. | 265 | 0.92 | 1.15 | 19.27 | 9.49 | 9.97 |
| 杜鹃Rhododendron simsii Planch. | 115 | 0.99 | 1.05 | 8.36 | 5.49 | 4.97 |
| 白马骨Serissa serissoides (DC.) Druce | 155 | 0.35 | 0.25 | 11.27 | 0.78 | 4.10 |
| 山莓Rubus corchorifolius L.f. | 110 | 0.93 | 0.32 | 8.00 | 1.59 | 3.30 |
| 金丝桃Hypericum monogynum L. | 95 | 0.40 | 0.13 | 6.91 | 0.44 | 2.49 |
| 胡枝子Lespedeza bicolor Turcz. | 80 | 0.30 | 0.08 | 5.82 | 0.50 | 2.13 |
| 茅莓Rubus parvifolius L. | 40 | 0.90 | 0.05 | 2.91 | 0.75 | 1.24 |
| 小果蔷薇Rosa cymosa Tratt. | 35 | 1.06 | 0.15 | 2.55 | 0.87 | 1.19 |
| 冬青Ilex chinensis Sims | 30 | 0.75 | 0.09 | 2.18 | 0.64 | 0.97 |
| 白背叶Mallotus apelta (Lour.) Muell. Arg. | 5 | 1.50 | 0.35 | 0.36 | 1.37 | 0.70 |
| 茅栗Castanea seguinii Dode | 15 | 1.00 | 0.07 | 1.09 | 0.50 | 0.55 |
| 蜡子树Ligustrum leucanthum(S.Moore) P.S.Green | 15 | 0.47 | 0.02 | 1.09 | 0.29 | 0.47 |
| 忍冬Lonicera japonica Thunb. | 10 | 0.30 | 0.02 | 0.73 | 0.02 | 0.26 |
| 菝葜Smilax china L. | 10 | 0.60 | 0.01 | 0.73 | 0.02 | 0.25 |
| 茶荚蒾Viburnum setigerum Hance | 5 | 0.30 | 0.00 | 0.36 | 0.08 | 0.15 |
| 灰白毛莓Rubus tephrodes Hance | 5 | 0.30 | 0.01 | 0.36 | 0.04 | 0.14 |
| 合计Total | 1375 | — | 100 | 100 | 100 | 100 |
| 物种 Species |
平均高 Mean H/m |
平均盖度 Mean C/% |
相对多度 RA/% |
相对盖度 RC/% |
重要值 IV/% |
| 褐果薹草Carex brunnea Thunb. | 0.21 | 18.92 | 50.55 | 62.62 | 49.49 |
| 阔鳞鳞毛蕨Dryopteris championii (Benth.) C. Chr. | 0.25 | 2.75 | 12.09 | 9.10 | 11.97 |
| 三脉紫菀Aster ageratoides Turcz. | 0.41 | 2.21 | 13.19 | 7.31 | 11.73 |
| 芒Miscanthus sinensis Anderss. | 0.73 | 4.17 | 6.59 | 13.79 | 9.74 |
| 海金沙Lygodium japonicum (Thunb.) Sw. | 0.13 | 0.33 | 4.40 | 1.10 | 3.79 |
| 凹叶景天Sedum emarginatum Migo | 0.05 | 0.08 | 4.40 | 0.28 | 3.52 |
| 狗脊Woodwardia japonica (L. F.) Sm. | 0.20 | 0.75 | 2.20 | 2.48 | 2.54 |
| 山麦冬Liriope spicata (Thunb.) Lour. | 0.30 | 0.42 | 2.20 | 1.38 | 2.17 |
| 长萼堇菜Viola inconspicua Blume | 0.05 | 0.08 | 2.20 | 0.28 | 1.80 |
| 千里光Senecio scandens Buch.-Ham. ex D. Don | 0.15 | 0.25 | 1.10 | 0.83 | 1.62 |
| 小蓬草Conyza canadensis(L.) Cronq. | 0.25 | 0.25 | 1.10 | 0.83 | 1.62 |
| 合计Total | — | — | 100 | 100 | 100 |
灌木层高度级集中于1.5 m以下, 共有1125株, 占灌木层总个体数的81.82%, 其中以1—1.5 m高度级个体数最多, 达440株, 占总个体数的32%(图 2); 1.5 m以上的植株有250株, 其中榅桲占个体数比为98%, 其余皆为白背叶。群落优势种榅桲的高度级整体缺乏规律性, 最高为3.8 m, 以1.5—2 m高度级个体数最多(90株), 其次为0-0.5 m(70株)、1—1.5 m(65株)和2.5—3 m(65株), 0.5—1 m高度级仅有5株。

|
| 图 2 褒忠山榅桲灌丛灌木层的高度级结构及榅桲种群的高度级结构和龄级结构 Fig. 2 The height structure of all shrubs, the height structure and the age structure of Cydonia oblonga population in the Cydonia oblonga shrubland of Baozhong Mountain |
榅桲种群龄级分布整体呈近倒“J”型[36], 种群年龄结构完整, 为增长型种群(图 2)。各龄级中, Ⅰ龄级个体数量最多, 占总数的22.08%, 其次为第Ⅱ、Ⅲ龄级, 均占总数的20.78%。各发育阶段中幼苗(Ⅰ)、小树(Ⅱ—Ⅲ)、中树(Ⅳ—Ⅴ)、大树(Ⅵ—Ⅶ)和老树(Ⅷ)占总数比例分别为22.08%、41.56%、27.27%、7.79%、和1.30%, 整体来看小树个体数量最多, 在往后各发育阶段个体数量下降平缓, 但幼苗个体数量却远少于小树, 说明该种群繁殖更新能力略显不足, 而后期生长趋势较好。
榅桲种群V1, V3-V5, V7大于0, 而V2和V6等于0, 说明种群龄级结构表现为“增长-稳定-增长-稳定-增长”的动态关系(表 4)。数量动态变化指数Vpi>V′pi>0, 说明榅桲种群整体呈现稳定增长趋势, 但V′pi趋近于0, 可见该种群在干扰情况下增长趋势不明显。随机干扰风险极大值P极大=12.5%, 说明榅桲种群对外界随机干扰具有高敏感性, 种群抗干扰能力弱。
| V1 | V2 | V3 | V4 | V5 | V6 | V7 | Vpi | V′pi | P极大 |
| 5.88 | 0.00 | 12.50 | 50.00 | 57.14 | 0.00 | 66.67 | 21.05 | 2.63 | 12.5 |
榅桲种群标准化存活数量(lx)随着龄级的增加而逐渐减少(表 5), 符合物种生物学特性。生命期望值(ex)整体呈逐渐下降趋势, 仅在第Ⅵ龄级略有波动, 说明榅桲在幼苗(Ⅰ)和小树(Ⅱ—Ⅲ)发育时期的生理活动最为旺盛。榅桲龄级间死亡率(qx)和消失率(kx)在第Ⅱ、Ⅵ龄级为0, 说明在小树(Ⅱ—Ⅲ)和大树(Ⅵ—Ⅶ)发育阶段内, 竞争未造成其个体数量的减少。在不考虑第Ⅱ、Ⅵ龄级的情况下, 龄级间死亡率(qx)和消失率(kx)从Ⅰ—Ⅷ整体呈逐渐上升的趋势, 这与生命期望值(ex)逐渐下降紧密相关。
| 龄级 Age class |
径级/cm DBH class/cm |
Ax | lx | lnlx | dx | qx | Lx | Tx | ex | Kx | Sx |
| Ⅰ | DBH<2 | 85 | 1000.00 | 6.91 | 58.82 | 0.06 | 970.59 | 4029.41 | 4.03 | 0.06 | 0.94 |
| Ⅱ | 2≤DBH<4 | 80 | 941.18 | 6.85 | 0.00 | 0.00 | 941.18 | 3058.82 | 3.25 | 0.00 | 1.00 |
| Ⅲ | 4≤DBH<6 | 80 | 941.18 | 6.85 | 117.65 | 0.13 | 882.35 | 2117.65 | 2.25 | 0.13 | 0.88 |
| Ⅳ | 6≤DBH<8 | 70 | 823.53 | 6.71 | 411.76 | 0.50 | 617.65 | 1235.29 | 1.50 | 0.69 | 0.50 |
| Ⅴ | 8≤DBH<10 | 35 | 411.76 | 6.02 | 235.29 | 0.57 | 294.12 | 617.65 | 1.50 | 0.85 | 0.43 |
| Ⅵ | 10≤DBH<12 | 15 | 176.47 | 5.17 | 0.00 | 0.00 | 176.47 | 323.53 | 1.83 | 0.00 | 1.00 |
| Ⅶ | 12≤DBH<14 | 15 | 176.47 | 5.17 | 117.65 | 0.67 | 117.65 | 147.06 | 0.83 | 1.10 | 0.33 |
| Ⅷ | 14 ≤DBH<16 | 5 | 58.82 | 4.07 | 58.82 | 1.00 | 29.41 | 29.41 | 0.50 | 4.07 | 0.00 |
拟合回归得到榅桲种群相应存活曲线模型: y=2408.2e-0.404x(R2=0.8738), y=1977.2x-1.222(R2=0.6599), 可见指数函数的R2更大, 说明榅桲种群的存活曲线更趋近于Deevey-Ⅱ型。
2.2.3 种群的生存分析榅桲种群生存率(S(i))和累计死亡率(F(i))随龄级增大而分别呈单调递减和单调递增趋势, 在第Ⅰ—Ⅲ和Ⅴ—Ⅷ龄级变化缓和, 而在第Ⅲ—Ⅴ龄级升降变化剧烈, 且相交于第Ⅲ、Ⅳ龄级之间(图 3), 危险率(λ(ti))变化趋势与累计死亡率基本一致, 死亡密度(f(ti))在第Ⅰ—Ⅱ龄级从最高急剧降至最低, 而后整体呈波动变化, 说明榅桲在幼苗和小树阶段面临的竞争压力较小, 在中树阶段因为较强的种内竞争而使得个体数量逐渐减少, 直至生长为大树具有更强的竞争优势, 从而使得升降变化趋于缓和。整体来看, 榅桲种群动态表现为前期优势, 中期减少, 后期稳定的特点。

|
| 图 3 榅桲种群的生存率(S(i))、累计死亡率(F(i))、死亡密度(f(ti))和危险率(λ(ti))函数 Fig. 3 The survival rate, cumulative mortality rate, mortality density rate and hazard rate function curves of Cydonia oblonga population |
如(图 4)可见, 在经历2个龄级时间后, 未来除第Ⅲ、Ⅶ龄级外, 其余龄级个体数量皆有小幅度上升, 而经历4、6、8个龄级时间后, 未来所有龄级个体数量均呈增长趋势, 说明榅桲种群在未来个体数量会稳定增加。整体来看, 未来榅桲种群个体数量于第Ⅳ龄级之后增幅较大, 而Ⅱ—Ⅳ龄级增幅较小。

|
| 图 4 榅桲种群的时间序列预测模型 Fig. 4 The time sequence prediction of Cydonia oblonga population M0, 各龄级现有个体数Number of individuals in each age group; M2, 各龄级在未来2个龄级时间后的个体数Number of individuals in each age class after the next two age classes; M4, 各龄级在未来4个龄级时间后的个体数Number of individuals in each age class after the next four age classes; M6, 各龄级在未来6个龄级时间后的个体数Number of individuals in each age class after the next six age classes; M8, 各龄级在未来8个龄级时间后的个体数Number of individuals in each age class after the next eight age classes |
植物群落的区系成分取决于该地区的生存条件和该地区植物的适应性。研究区地处中亚热带地区, 本地植物区系应当表现出明显的亚热带过渡性质, 即热带性质属和温带性质属比例相当[37]。而榅桲灌丛温带性质属和热带性质属分别占75%和25%, 两者比达3∶1, 温带性质较大围山杜鹃灌丛[29]和庐山灌丛[38]更为显著。从物种组成上来看, 其原因在于榅桲群落生于孤山顶峰, 冷凉多风的环境和贫瘠干燥的土壤等条件致使热带性质为主的常绿木本植物(3种)生存几率大大减小, 而以温带性质为主的落叶灌木(13种)占据主要地位, 如灌木层(除榅桲外)重要值前5的中华绣线菊、杜鹃、白马骨(Serissa serissoides)皆为温带性质, 另2种则为世界分布。同样, 在草本层中重要值前5的种子植物也仅有芒为热带性质, 而如三脉紫菀、凹叶景天(Sedum emarginatum)、山麦冬(Liriope spicata)等皆为温带性质。起源于高加索地区的榅桲本就属于温带内陆性质, 能够适应干旱期长、降水量小的Iran-Turanian地理分布区气候[39], 因此对于研究区阴凉干燥的生境同样具有极强的适应能力。
垂直结构是指群落在空间上的垂直分化现象, 它是群落内物种对生境适应能力的体现, 也是种间竞争的直接结果。榅桲群落分层明显, 灌木层个体以1—1.5 m高度级最多(图 2), 这是因为在孤山顶峰低温风大、贫瘠干燥等不利条件的影响下, 群落内灌木层植物集体采取了较为保守的生活史策略以适应不利环境, 使得中等高度个体集中[29, 40]。榅桲种群个体集中于1—3 m的中等高度(图 2), 占总个体数的71.43%, 说明其同样采取了减少高生长的适应策略。其个体数在1.5—2 m高度级最多, 而在0.5—1 m高度级榅桲种群个体最少, 可能是因为灌木层个体多集中于1—1.5 m高度级, 一定程度上限制了榅桲幼苗的高生长, 当榅桲幼苗获得高生长机会时, 便会迅速突破这种的限制。该群落灌木层1.5 m以上高度级中榅桲个体数占总个体数的98%, 说明榅桲突破低矮灌木层对其高生长的限制后, 迅速占领了灌木层上层空间, 最终形成了孤山顶峰以榅桲为单优势种的群落。
3.2 种群动态与发展趋势种子和幼苗是植物生活史的关键环节, 幼苗阶段是种群天然更新的瓶颈[32]。实地调查表明, 榅桲种群存在有性繁殖和根蘖繁殖两种方式, 但以根蘖繁殖为绝对优势, 这是因为榅桲种子存在休眠特性[41], 且易被鸟兽取食, 再加上该种群生于土层瘠薄干燥的孤山顶峰, 致使该种群有性繁殖几率较低。幼苗的数量是影响一个种群能否长期稳定的重要因素[42], 如海南省的蕉木(Oncodostigma hainanense)种群[43]第Ⅰ龄级幼苗个体数量与第Ⅱ龄级之比为0.36, 幼苗数量稀少, 致使种群自我更新能力不佳, 呈衰退趋势; 而昆嵛山五莲杨(Populus wulianensis)[32]种群第Ⅰ级个体数量与第Ⅱ龄级之比却高达22, 尽管因强烈的自疏和他疏作用会导致幼苗死亡率最高可达95.5%, 但依旧能够保证种群的增长趋势。本研究中榅桲种群幼苗第Ⅰ龄级个体数量与第Ⅱ龄级之比为1.06, 介于上述两种情况之间, 第Ⅰ—Ⅲ龄级个体数量基本稳定, 生存期望值均达2以上, 危险率和累计死亡率保持较低水平, 说明该种群繁殖更新能力一般, 幼苗数量较少, 可以避免自疏作用的筛选, 但生理活动旺盛, 即使存在较强的物种竞争, 也能顺利进入小树发育阶段, 基本维持种群前期的自我更新。
种群动态是生物因素与非生物因素共同作用的结果[44]。静态生命表和生存分析结果显示, 榅桲种群在第Ⅳ—Ⅴ龄级消失率、累计死亡率和危险率急剧升高, 死亡密度达到最高值, 而生存率和生存期望值则持续降低, 说明该种群在中树阶段经受了强烈的环境筛作用, 造成了个体数量的大量死亡, 与云南省的馨香木兰种群[45]中龄级个体大量死亡的情况一致。其原因在于种群在幼龄阶段集群分布有利于发挥群体效应得以生存, 到成年阶段种群会通过拥挤效应和自疏作用来降低集群强度以保证个体获取足够的资源[46], 且榅桲种群根蘖繁殖为主所导致的聚集分布模式会进一步加强种内竞争的自疏作用。随着榅桲生长至中树阶段, 种内竞争强度逐渐达到峰值, 从而大量淘汰了较弱个体, 剩余个体的生存能力则得到了增强, 这也解释了整体呈逐渐下降趋势的生命期望值为何在第Ⅵ龄级会出现增长。而后在Ⅶ—Ⅷ龄级, 榅桲种群生命期望值、生存率持续下降至最低, 累计死亡率和危险率平缓上升至最高, 说明在不利生境的限制下, 树木个体生长至老龄阶段已逐渐进入生理衰老期, 环境抗逆性减弱。在现有因素不变的情况下, 榅桲种群具有明显的增长趋势。其龄级结构完整, 增长曲线表现出“增长-稳定-增长-稳定-增长”的动态关系, 说明其合理的龄级个体数量分布保证了种群的增长潜力。存活曲线趋近于Deevey-Ⅱ型, 表明当前种群龄级结构总体处于稳定状态, 这可能是因为榅桲种群幼苗及小树阶段能够适应生境, 个体数量稳定, 从而保证了后续所有龄级个体数量的正常补给。时间序列预测结果显示榅桲种群个体数量在未来整体呈增长趋势, 其中Ⅱ—Ⅳ龄级增长缓慢, Ⅴ—Ⅷ增长幅度较大, 说明榅桲种群的增长趋势主要依靠中树和大树个体数量的增长, 与长白山鱼鳞云杉(Picea jezoensis var. microsperma)种群和蒙古栎(Quercus mongolica)种群类似[35], 而其幼苗和小树数量会持续维持在一个稳定水平, 有限的繁殖更新能力成为了限制榅桲种群增长速率提高的主要因素。然而, V′pi趋近于0和P极大为12.5%体现了该种群在外界干扰情况下的脆弱性, 一旦在外界环境的强干扰下导致幼苗数量减少, 其稳定的龄级结构和增长趋势易遭到破坏, 最终转变为衰退种群。
3.3 归化及入侵性分析归化植物是指在无人类干预的前提下可以自行繁衍, 并且能够长期维持种群自我更新的外来植物[2]。实地调查发现, 榅桲集中分布于褒忠山顶峰, 远离人为栽植环境, 表明了其逸生性, 榅桲种群果期为7—9月, 林下存在实生苗, 说明其具有独立完成生命周期的能力, 且具有完整的高度结构和龄级结构(图 2), 表明该种群已建立稳定种群。榅桲在其所在群落的灌木层的相对盖度达77.11%, 重要值达67.12%(表 2), 表明榅桲种群是群落中的优势种群, 对群落的结构和群落环境的形成有明显的控制作用。在无人为干扰的情况下, 种群具有小幅增长趋势(表 4), 且经历2、4、6、8个龄级后, 种群数量将稳定增加, 表明该种群可长期维持优势地位。基于褒忠山榅桲远离人为栽植环境、可独立完成生命周期并建立优势种群、可长期维持种群稳定和优势地位等几点事实, 榅桲符合外来木本植物归化定义及广义标准[25], 因此判定榅桲已在中国成功归化。
植物在原生生境外成为入侵植物要经历引入、逃逸、归化、入侵四个阶段[47], 当归化植物改变且威胁本地生物多样性并造成经济损失时, 就构成了入侵, 因此对于外来植物是否入侵的判断应当慎重。经走访调查, 我们了解到报恩寺百年前曾有位住持喜珍贵名木, 要求弟子收集并栽植于寺庙内, 住持曾访遍东南诸省研讨佛学, 累传弟子数十万, 并前后两次访问福建涌泉寺[48]。榅桲最先在两汉至南北朝时期由丝绸之路引入中国, 是使臣献贡的皇家贡品, 可见其高贵[49], 榅桲在福建省明溪县有大规模种植, 且栽培历超过300年[50], 因此我们推测褒忠山的榅桲可能引种于明溪县, 而后逸生。参照郑景明[51]外来木本植物入侵预测和评价方法, 从入侵历史、物种特征、繁殖压力等预测指标分析, 榅桲在自然分布区之外没有过入侵记录; 褒忠山榅桲种群仅于顶峰形成优势种群, 散生个体均距离该种群不超过200 m, 且皆生于山坡草丛或荒地, 在相近的黄檀群落、山胡椒群落中未见榅桲的踪迹; 褒忠山“成年”榅桲株数在40株左右, 每株母树挂果较少, 均在10枚以下, 且榅桲种子存在休眠特性, 易被鸟兽取食。因此初步认为褒忠山榅桲种群入侵性较低, 但基于一次的调查结果来讨论其潜在的入侵风险可能存在偏差, 其入侵性还需更多数据支撑。
4 结论本研究在对湖南褒忠山榅桲所在群落的组成和结构调查的基础上, 对榅桲种群结构和动态进行了定量分析, 结果表明榅桲在中国已归化, 能够在野外独立完成生命周期, 在局域特殊生境中建立优势种群, 并能长期维持种群的稳定和群落中的优势地位。榅桲作为一种古老而珍稀的果树, 在经济林、医药和香料等方面都具有广阔的应用价值和发展前景, 其成功归化对榅桲的进一步开发利用和区域外来物种管理具有重要意义。由于本研究仅在目前中国发现的唯一归化地就群落和种群现状开展研究, 关于榅桲种群的实际发展动态及其对本地生态系统的影响, 今后还需要扩大调查区域和持续监测其种群动态。
| [1] |
Richardson D M, Pysek P, Rejmanek M, Barbour M G, Dane Panetta F, West C J. Naturalization and invasion of alien plants: concepts and definitions. Diversity & Distributions, 2000, 6(2): 93-107. |
| [2] |
Pyšek P, Richardson D M, Rejmánek M, Webster G L, Williamson M, Kirschner J. Alien plants in checklists and floras: towards better communication between taxonomists and ecologists. TAXON, 2004, 53(1): 131-143. DOI:10.2307/4135498 |
| [3] |
Khuroo A A, Reshi Z A, Malik A H, Weber E, Rashid I, Dar G H. Alien flora of India: taxonomic composition, invasion status and biogeographic affiliations. Biological Invasions, 2012, 14(1): 99-113. DOI:10.1007/s10530-011-9981-2 |
| [4] |
Zenni R D. The naturalized flora of Brazil: a step towards identifying future invasive non-native species. Rodriguésia, 2015, 66(4): 1137-1144. DOI:10.1590/2175-7860201566413 |
| [5] |
Ansong M, Pergl J, Essl F, Hejda M, van Kleunen M, Randall R, Pyšek P. Naturalized and invasive alien flora of Ghana. Biological Invasions, 2019, 21(3): 669-683. DOI:10.1007/s10530-018-1860-7 |
| [6] |
Wu S H, Sun H T, Teng Y C, Rejmánek M, Chaw S M, Yang T Y A, Hsieh C F. Patterns of plant invasions in China: Taxonomic, biogeographic, climatic approaches and anthropogenic effects. Biological Invasions, 2010, 12(7): 2179-2206. DOI:10.1007/s10530-009-9620-3 |
| [7] |
Jiang H, Fan Q, Li J T, Shi S, Li S P, Liao W B, Shu W S. Naturalization of alien plants in China. Biodiversity and Conservation, 2011, 20(7): 1545-1556. DOI:10.1007/s10531-011-0044-x |
| [8] |
Yan X L, Wang Z H, Ma J S. The Checklist of the Naturalized Plants in China. Shanghai: Shanghai Scientific and Technical Publishers, 2019.
|
| [9] |
许光耀, 李洪远, 莫训强, 孟伟庆. 中国归化植物组成特征及其时空分布格局分析. 植物生态学报, 2019, 43(7): 601-610. |
| [10] |
林秦文, 于胜祥, 唐赛春, 崔夏, 高信芬, 王焕冲, 刘全儒, 马金双. 中国外来归化植物的编目现状及有关问题. 植物生态学报, 2021, 45(11): 1275-1280. |
| [11] |
张云霞, 杨浩哲. 河南省归化植物一新记录种: 银背委陵菜(Potentilla argentea L.). 郑州师范教育, 2022, 11(4): 13-16. |
| [12] |
高亚红, 丁华娇, 胡梦霄, 陈晓玲. 中国归化植物新记录——北美细叶芹. 亚热带植物科学, 2022, 51(3): 217-218. |
| [13] |
王刚狮, 时宝凌, 贾泽凡, 权立森. 山西省归化植物新资料. 安徽农业科学, 2022, 50(7): 123-125. |
| [14] |
严靖, 闫小玲, 李惠茹, 杜诚, 马金双. 华东地区归化植物的组成特征、引入时间及时空分布. 生物多样性, 2021, 29(4): 428-438. |
| [15] |
Cantamutto M, Poverene M, Peinemann N. Multi-scale analysis of two annual Helianthus species naturalization in Argentina. Agriculture, Ecosystems & Environment, 2008, 123(1/2/3): 69-74. |
| [16] |
Baturin S O. Naturalization of Fragaria×ananassa duch. in western Siberia. Contemporary Problems of Ecology, 2016, 9(3): 376-383. DOI:10.1134/S1995425516030021 |
| [17] |
Liccari F, Boscutti F, Sigura M, Tordoni E, Carpanelli A, Valecic M, Bacaro G. First report of naturalization of Houttuynia cordata Thunb. 1783(Saururaceae) in Italy. Rendiconti Lincei Scienze Fisiche e Naturali, 2021, 32(2): 287-293. DOI:10.1007/s12210-021-00986-2 |
| [18] |
Howard L F, Minnich R A. The introduction and naturalization of Schinus molle (pepper tree) in Riverside, California. Landscape and Urban Planning, 1989, 18(2): 77-95. DOI:10.1016/0169-2046(89)90001-7 |
| [19] |
Frappier B, Eckert R T. Utilizing the USDA PLANTS database to predict exotic woody plant invasiveness in New Hampshire. Forest Ecology and Management, 2003, 185(1/2): 207-215. |
| [20] |
Vianna-Filho M D, Alves R J V, Peng Y Q, Pereira R A S. Naturalization of the bodhi fig tree (Ficus religiosa L.-Moraceae) in Brazil. Bioscience Journal, 2017, 33(1): 177-182. |
| [21] |
谭菊荣, 袁位高, 李婷婷, 许国良, 吴正柱, 陈小明, 李卫忠, 许惠春, 库伟鹏, 朱锦茹, 吴初平. 极小种群野生植物细果秤锤树种群结构与动态特征. 生态学报, 2022, 42(9): 3678-3687. |
| [22] |
Abdollahi H. A review on history, domestication and germplasm collections of quince (Cydonia oblonga Mill.) in the world. Genetic Resources and Crop Evolution, 2019, 66(5): 1041-1058. DOI:10.1007/s10722-019-00769-7 |
| [23] |
刘启振, 王思明. 陆上丝绸之路传入中国的域外农作物. 中国野生植物资源, 2016, 35(6): 5-11, 20. |
| [24] |
罗华, 郝兆祥, 张忠涛, 毕润霞, 陈颖. 国内榅桲种质资源研究现状. 山西果树, 2018(6): 18-22, 27. |
| [25] |
严靖, 闫小玲, 李惠茹, 杜诚, 马金双. 华东地区归化植物的组成特征、引入时间及时空分布. 生物多样性, 2021, 29(4): 428-438. |
| [26] |
祁承经. 湖南植被. 长沙: 湖南科学技术出版社, 1990.
|
| [27] |
生态系统固碳项目技术规范编写组. 生态系统固碳观测与调查技术规范. 北京: 科学出版社, 2015.
|
| [28] |
谢宗强, 唐志尧. 中国灌丛生态系统固碳现状、变化和机制研究调查规范, 20111.
|
| [29] |
李家湘, 张旭, 谢宗强, 卢从发, 涂向阳, 寻院. 湖南大围山杜鹃灌丛的群落组成及结构特征. 生物多样性, 2015, 23(6): 815-823. |
| [30] |
方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [31] |
吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1991, 增刊Ⅳ: 1-139. |
| [32] |
吴其超, 臧凤岐, 李呈呈, 马燕, 高燕, 郑勇奇, 臧德奎. 濒危树种五莲杨种群结构与动态特征. 生态学报, 2021, 41(12): 5016-5025. |
| [33] |
陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214-217. |
| [34] |
Farahat E A. Age structure and static life tables of the endangered Juniperus phoenicea L. in North Sinai Mountains, Egypt: implication for conservation. Journal of Mountain Science, 2020, 17(9): 2170-2178. |
| [35] |
陈科屹, 张会儒, 张博, 何友均. 长白山北坡天然次生林典型建群种的种群结构及动态特征. 生态学报, 2021, 41(13): 5142-5152. |
| [36] |
Liyew B, Tamrat B, Sebsebe D. Woody species composition and structure of Amoro Forest in West Gojjam Zone, North Western Ethiopia. Journal of Ecology and the Natural Environment, 2018, 10(4): 53-64. |
| [37] |
陈卫娟. 中亚热带常绿阔叶林植物区系地理研究[D]. 上海: 华东师范大学, 2006.
|
| [38] |
罗光坦, 李景信. 庐山灌丛的群落特征. 哈尔滨师范大学自然科学学报, 1993, 9(3): 100-107. |
| [39] |
Ghafari S, Ghorbani A, Moameri M, Mostafazadeh R, Bidarlord M. Composition and structure of species along altitude gradient in Moghan-Sabalan rangelands, Iran. Journal of Mountain Science, 2018, 15(6): 1209-1228. |
| [40] |
Bin Y, Ye W H, Muller-Landau H C, Wu L F, Lian J Y, Cao H L. Unimodal tree size distributions possibly result from relatively strong conservatism in intermediate size classes. PLoS One, 2012, 7(12): e52596. |
| [41] |
包建平, 张绍铃, 成希, 崔艳波, 刘妮. 豆梨和榅桲种子生活力的快速测定. 种子, 2010, 29(2): 119-122. |
| [42] |
解婷婷, 苏培玺, 周紫鹃, 张海娜, 李善家. 荒漠绿洲过渡带沙拐枣种群结构及动态特征. 生态学报, 2014, 34(15): 4272-4279. |
| [43] |
江杏香, 陈玉凯, 吴石松, 陈庆. 海南濒危植物蕉木种群结构与动态特征. 南京林业大学学报: 自然科学版, 2021, 45(1): 116-122. |
| [44] |
Velázquez E, Kazmierczak M, Wiegand T. Spatial patterns of sapling mortality in a moist tropical forest: consistency with total density-dependent effects. Oikos, 2016, 125(6): 872-882. |
| [45] |
徐加汉, 司马永康, 徐涛, 猫玉白, 余鸿. 极小种群野生植物馨香木兰的种群结构及动态. 西部林业科学, 2016, 45(2): 35-40, 51. |
| [46] |
Molles M. Ecology: concepts and applications. New York: Mcgraw-hill, 1998.
|
| [47] |
Williamson M, Fitter A. The varying success of invaders. Ecology, 1996, 77(6): 1661-1666. |
| [48] |
湘乡县志编纂委员会. 湘乡县志. 长沙: 湖南人民出版社, 1993.
|
| [49] |
刘启振, 王思明. 陆上丝绸之路传入中国的域外农作物. 中国野生植物资源, 2016, 35(6): 5-11, 20. |
| [50] |
张诚. 明溪县榅桲资源调查. 福建果树, 2006(1): 37-38. |
| [51] |
郑景明, 李俊清, 孙启祥, 周金星. 外来木本植物入侵的生态预测与风险评价综述. 生态学报, 2008, 28(11): 5549-5560. |