 2023, Vol. 43
2023, Vol. 43文章信息
- 袁换欢, 严家宝, 张建亮, 王智, 徐网谷, 张昊楠
- YUAN Huanhuan, YAN Jiabao, ZHANG Jianliang, WANG Zhi, XU Wanggu, ZHANG Haonan
- 上海市气候变化和人类活动对植被物候的影响
- Influences of climate change and human activities on vegetation phenology of Shanghai
- 生态学报. 2023, 43(21): 8803-8815
- Acta Ecologica Sinica. 2023, 43(21): 8803-8815
- http://dx.doi.org/10.20103/j.stxb.202209192675
-
文章历史
- 收稿日期: 2022-09-19
- 网络出版日期: 2023-06-27





2. 中国科学院地理科学与资源研究所, 北京 100101
2. Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China
随着社会经济的发展和人类生产生活方式的改变, 人类活动对生态环境的影响日益显著[1]。IPCC第五次气候变化评估报告明确指出人类活动是20世纪中期以来全球气候变暖的重要原因[2]。然而, 植被物候是气候变化最敏感的指示器[3—5], 其动态变化与植被生长期长度密切相关, 并且物候期变化影响地表反照率、粗糙度、蒸散发、CO2浓度等从而调节气候变化[6—7]。此外, 上海作为我国经济高速发展的超大城市, 拥有与主城区相比受人类活动影响较小和环境差异较大的河口沙岛(崇明岛), 是模拟未来气候变化和人类活动对生态环境影响的良好试验场。因此, 深入研究上海市植被物候及其驱动因子, 尤其是人类活动因子, 对城市应对未来气候变化和改善城市生态环境具有重要的意义。
目前已有大量关于植被物候时空变化及其驱动因子的研究, 认为生长开始期(start of growth season, SOS)呈总体提前趋势, 生长结束期(end of growth season, EOS)呈总体延迟趋势, 并且不同区域植被物候变化存在较大空间差异[8]。如我国东北、华北和新疆等大部分随气候变暖呈现SOS提前和EOS延迟趋势[9], 而长江中下游部分地区春季物候呈延迟趋势[10]。温度、降水和光照等被认为是主要调控植被物候变化的气候因子, 主要原因是温度影响解除植被生理休眠和生态休眠期所需的冷激和积温, 是植被物候的主要影响因子[11—13]。不对称的增温效应和对春季/秋季物候的差异响应, 促使温度对植被物候呈非线性响应并对温度敏感性下降[3, 14—16]。光周期可以弥补冷激或积温需求不足时通过光照的生长许可阈值促使春季物候(Start of growing season, SOS)和秋季物候(End of growing season, EOS)的发生[17—18]。降水主要影响受水分限制的干旱半干旱区生态系统的植被物候[19—20], 甚至导致干湿季分明地区的植被形成多个生长周期[21]。随着人类活动影响强度的增加, 人类活动因子对植被物候的影响逐渐受到关注[22]。植树造林和退耕还草/林等生态修复工程显著提高植被覆盖度, 促使碳汇量增加[23—25]。然而, 伴随城市人口密度[26]、地表透水性(土地覆盖变化)[27]、城市光照(光周期延长)[28]和污染性气体排放(太阳辐射减弱)等引发的城市热岛和其他局部小气候严重影响植被生长环境[29]。此外, 夜间灯光改变自然夜间环境影响动物的行为、生理和生活史[30]。同时诱导植物光合作用影响维管植物的生长和资源分配[31], 甚至群落结构组成和稳定[32], 导致研究夜间灯光的生态影响成为近年来全球变化研究的重要焦点和预测城市植被对人类活动响应的关键[33]。
鉴于长时间序列夜间灯光数据兼具包含人类活动和光照强度信息, 适于动态分析和直接影响植被物候的特点, 被认为是监测区域尺度人类活动和土地利用强度的重要数据源[34—35]。因此, 本文首先通过2001—2020年上海市SOS和EOS及其驱动因子(温度、降水、光合有效辐射和夜间灯光)重心偏移情况, 分析郊区和城区物候及其驱动因子的时空变化差异; 随后利用地理探测器、偏相关分析和变量投影重要性指标, 从时间和空间两个尺度讨论植被物候对其驱动因子的响应; 在此基础上, 揭示上海市人类活动(夜间灯光)和环境因子(温度、降水和光合有效辐射)对郊区和城区植被物候的贡献。本研究将有利于加深城市生态系统中人与自然交互作用的理解, 并为城市应对气候变化和改善生态环境提供理论支持。
1 研究区和研究方法 1.1 研究区概况上海市地处地势平坦的长江三角洲平原东南, 长江与钱塘江的入海口。上海市辖徐汇区、虹口区、长宁区、静安区、杨浦区、闵行区、黄埔区、普陀区、金山区、松江区、宝山区、嘉定区、浦东新区、青浦区、奉贤区和崇明区16市辖区。上海市气候温和、湿润, 土壤疏松, 雨量充沛属于亚热带季风气候。2020年平均气温17.80℃, 年降水量为1660.8mm, 平均海拔2.19m左右。2020年上海市土地利用变化主要是城市和建筑用地, 其城市建成区面积是2001年的1.25倍。由于上海市城区和郊区受人类活动和土地利用变化剧烈程度存在显著差异, 导致其植被物候的气候响应也存在差异。崇明区与上海市隔江相望, 较好的剥离主城区人类活动对其植被的潜在影响, 因此将崇明区作为郊区, 崇明区以外的作为城区, 开展城区和郊区植被物候对气候变化和人类活动响应的对比分析研究。
1.2 数据来源物候数据来源于美国NASA的MODIS地表覆盖动态产品MCD12Q2(V6.1版), 时间段为2001—2020年, 空间分辨率为500m。平均温度和降水数据为气象数据来源于英国East Anglia大学的Climatic Research Unit (CRU, https://crudata.uea.ac.uk/cru/data/), 考虑卫星驱动产品采用不同模型公式对逐月数据采用薄板样条(Thin plate spline, TPS)生成1952—2019年ChinaClim_baseline和逐月气候异常面。进一步采用气候辅助插值法(Climatologically aided interpolation, CAI)将逐月异常面与ChinaClim_baseline叠加生成1km空间分辨率和月尺度的ChinaClim_time-series数据集[36]。辐射数据为参与光合生物量累积且影响植被类型、覆盖状况和植被生长状况的光合有效辐射分量(Fraction of photosynthetically Active Radiation, 辐射)数据, 是植被群体生理状况的重要指示参数[37], 该数据来源于MOD15A2H(V6.1版), 经过Savitzky-Golay滤波进行去噪处理, 并结合LDOPE工具随机对空间分辨率为500m, 时间分辨率为8d的辐射数据的辐射Lai_QC数据层进行质量评价[38]。
DMSP/OLS夜间灯光数据为2001—2012年美国军事气象卫星(Defense Meteorological Satellite program, DMSP)搭载的传感器(Operational Linescan System, OLS)该数据来自美国国家海洋和大气管理局(National Oceanic and Atmospheric Administration, NOAA)下属的国家地球物理数据中心(National Geophysical Data Center, NGDC)。此外, 采用2012—2020年美国科罗拉多矿业大学的SNPP-VIIRS数据, 通过数据融合获取的2001—2020年的夜间灯光数据[39], 该数据消除了云及火光等偶然噪声影响, 数据灰度值范围为0—63, 空间分辨率为1km, 该数据能够捕捉到城市灯光甚至小规模居民地、车流等发出的地强度灯光, 为城市人类活动研究提供有力的数据支撑。
1.3 分析方法重心分析法主要用于分析物候及其驱动因子的时空变化, 是地理学及生态学领域中引入物体所受的合力作用点的概念, 并将其运用于描述地理现象的变化程度和变化趋势的分析方法, 其计算公式如下:

|
(1) |
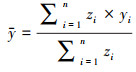
|
(2) |
式中, zi表示空间单元的中心的属性值, 点(xi, yi)为空间地i个单元的坐标, 点(x, y)表示空间所有单元的平均值。
地理探测器主要用于分析植被物候空间上的驱动因子, 探寻地理空间分异性, 揭示自然和社会经济背后驱动因子的一种统计学方法, 主要由4部分组成: 风险区探测(Risk area detector)、风险因子探测(Risk factor detector)、生态探测(Ecological detector)和交互探测(Interaction detector)。其中风险探测器主要用于探索单个驱动因子对植被春季物候开始和秋季物候结束的适宜性。因子探测主要用于检验驱动因子对植被物候变化的空间分布格局的影响强度。生态探测器主要是用来探究驱动因子在影响植被物候分布方面是否有显著差异。交互探测器可以识别驱动因子之间的交互作用效果[40]。
偏相关分析(公式3)用于分析研究某一因子对物候的影响时, 排除其他因子的影响。评价偏相关分析和相关分析显著的指标是t检验(公式4)。

|
(3) |
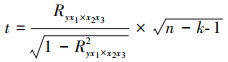
|
(4) |
式中, y为因变量, x1、x2和x3为自变量, Ryx1×x2x3表示控制x2和x3, 研究y表示x1的偏相关系数, Ryx1×x2表示控制x2研究y与x1的偏相关, Ryx1表示y与x1的相关系数。n表示构建模型使用的样本数量, k表示自变量的个数。
变量投影重要性指标(variable importance in projection, VIP, 公式5)用于定量比较分析不同因素对物候变化的影响程度, 识别驱动植被物候变化的主导驱动因子。

|
(5) |
式中, wjf是j变量和f分量的权重值, SSYf是第f个分量解释方差的平方和, J是解释变量的个数, SSYtotal是因变量解释方差的平方总和, F是分量的总数。wjf2给出了每个f组分中第j个变量的重要性, VIPj表征了j变量的贡献度。
2 结果与分析 2.1 上海市植被物候及驱动因子空间分布利用逐像元运算方法得到2001—2020年的得到上海市植被物候指标及其驱动因子的时空分布(图 1), 该空间分布直观显示了城区和郊区植被物候、环境因子以及夜间灯光的差异。其中植被EOS郊区为295d早于主城区植被EOS(302d)约7d, 春季物候城区SOS为77d早于郊区植被SOS(87d)约10d左右, 并且植被物候(EOS/SOS)沿主城区—城区—郊区的变化趋势逐渐变化(延迟/提前)。与此相似, 夜间灯光、温度、辐射也呈现出由主城区—城区—郊区逐步降低或增加的变化趋势, 并且城区的平均灯光为45.96高于郊区夜间灯光(15.66)2.9倍作用, 城区的平温度为16.82℃高于郊区(16.19℃)约0.63℃, 城区辐射为0.27低于郊区(0.34)0.07, 主要原因是城市高密度人口和第二产业造成的废气和烟尘高于郊区, 使日光照时间长度和太阳辐射总量减弱。此外, 城区年降水约1019.10mm高于郊区(1040.96mm)21.87mm, 可能原因包括(1)城市热岛效应导致热力对流增强, 空气层稳定性下降, 有利形成对流性降水; (2)城市空气中的凝结核高于郊区, 这有利于降水的形成。(3)城市下垫面粗糙度较大, 阻碍降水天气系统经过城市时间, 从而延长降水时长(增加降水量)。

|
| 图 1 研究区2001—2020年平均植被物候和驱动因子 Fig. 1 Average vegetation phenology and driving factors from 2001—2020 EOS: 秋季物候; SOS: 春季物候 |
以2001—2020年上海市植被物候及其驱动因子数据为基础, 通过ArcGIS重心模型分别获取不同年份空间分辨率为500m的像元中心点的植被物候和驱动因子重心时空变化信息, 研究植被物候重心演化趋势。如图 2所示, 2001—2020年上海市春季物候的重心呈现先大幅向东南偏移, 后小幅向西北转移, 最后向西南偏移, 并且春季物候的跨经度范围最大(1.48′)。秋季物候的重心呈先东北方向偏移, 后西南方向偏移, 其中2010年后秋季物候重心偏移趋势一致。夜间灯光重心经向上向东偏移, 而纬向上先大幅向北后小幅向北最后向南, 其中夜间灯光的跨经纬度范围仅次于春季物候, 尤其是经度(1.44′), 可能原因是主城区夜间灯光接近饱和, 而沿海扩展较快, 因此夜间灯光重心向沿海地区偏移。春季温度、降水和辐射的重心变化均有向最初位置回归的趋势, 并且春季气候因子纬向变化(1.04′)大于经向变化, 其中春季辐射的重心变化幅度大于温度和降水。秋季温度经向向东偏移纬向向北偏移, 降水经向先小幅向西后大幅向东, 纬向向北偏移, 辐射的重心转移方向为西南方向。总体而言, 2001—2020年春季物候和秋季物候及其驱动因子的重心均在上海市宝山区, 并且物候重心均发生偏移, 说明此期间物候的驱动因子发生变化。结合物候驱动因子重心变化, 发现夜间灯光重心偏移最高, 其次是辐射。由此可知, 夜间灯光和辐射可能是植被物候的主要驱动力。

|
| 图 2 2001—2020年植被物候及其驱动因子重心时空变化 Fig. 2 Spatial—temporal changes of the center of gravity in phenology and its drives during 2001—2020 |
采用地理探测器风险因子探测(表 1)植被物候分布影响发现, 2001—2020年选取的温度、降水、辐射和夜间灯光四个气候因子对植被物候显著影响(P < 0.05), 其中温度和降水对春季/秋季物候的影响权重低于辐射和夜间灯光, 并且夜间灯光对春季物候的影响权重最大为0.15高于辐射(q=0.13), 辐射对秋季物候的影响最大权重为0.11高于夜间灯光。为更深入的探讨植被物候分布的组合驱动及影响程度, 采用地理探测器中的交互探测模块进行评估。结果显示, 温度、降水、辐射和夜间灯光对植被物候影响的组合q值均大于其单因子, 尤其是春季物候多因子驱动影响权重显著提高。此外, 辐射和夜间灯光两个驱动因子在单因子以及交互因子的驱动下, 其q值均大于其他单因子或者交互因子, 并且辐射∩夜间灯光的组合驱动对春季物候和秋季物候的影响最大(q平均=0.22)。表明上海市植被物候驱动因子, 特别是指示人类活动强度的夜间灯光可通过调整优化影响植被物候的空间分布。
| SOS春季物候 | EOS秋季物候 | |||||||||
| 温度 | 降水 | 辐射 | 夜间灯光 | 温度 | 降水 | 辐射 | 夜间灯光 | |||
| 风险因子探测 | q | 0.08 | 0.05 | 0.13 | 0.15 | 0.02 | 0.01 | 0.11 | 0.05 | |
| Risk factor detector | P | < 0.05 | < 0.05 | < 0.05 | < 0.05 | < 0.05 | < 0.05 | < 0.05 | < 0.05 | |
| 交互探测 | 温度 | 0.08 | 0.02 | |||||||
| Interaction detector | 降水 | 0.14 | 0.05 | 0.06 | 0.01 | |||||
| 辐射 | 0.21 | 0.2 | 0.13 | 0.14 | 0.13 | 0.11 | ||||
| 夜间灯光 | 0.18 | 0.2 | 0.29 | 0.15 | 0.06 | 0.06 | 0.15 | 0.05 | ||
| SOS: 春季物候; EOS: 秋季物候; q statistic: q统计, 表示SOS或EOS的驱动因子(温度、降水、辐射和夜间灯光)对SOS或EOS空间分异的解释程度; P value: P值, 判断驱动因子对SOS或EOS的空间分异的解释是否通过显著性检验的依据 | ||||||||||
鉴于2001—2020年空间尺度上人类活动和气候变化对植被物候存在不同程度的影响, 郊区作为人类活动较少的区域, 较少的受外部因素的干扰与城区(人类活动强烈的地区)形成强烈的对比。分别对城区和郊区的春季物候和秋季物候进行风险因子和交互因子分析, 结果与总体相似降水、温度、辐射和夜间灯光对城区和郊区的植被春季物候和秋季物候显著影响(P < 0.05), 而城区的春季物候受夜间灯光的影响权重最大(q=0.26), 城区秋季物候和郊区春季/秋季物候受辐射的影响权重最大。
2.3 植被物候对气候变化及人类活动的响应2001—2020年春季物候驱动因子偏相关分析结果如图 3所示, 总体上温度对SOS的影响(R)作用最大, 高于降水、辐射和夜间灯光(R温度=-0.24、R降水=-0.08、R辐射=-0.20和R夜间灯光=0.05)。温度与城区/郊区的植被SOS的相关性高于其他因子(R城区温度= -0.26, R郊区温度=-0.20), 其次是辐射(R城区辐射=-0.23, R郊区辐射=-0.06)。城区温度、降水、辐射和夜间灯光对SOS的平均偏相关系数(R)高于郊区, 其中平均R城区辐射和R郊区辐射的差异最大为0.17, 夜间灯光对郊区和城区SOS的影响差异最小。

|
| 图 3 城区和郊区春季物候与驱动因子的偏相关系数空间及频次分布曲线 Fig. 3 Spatial and frequency distribution curve of partial correlation coefficients between urban and suburban spring phenology and driving factors |
仅考虑显著像元(P < 0.05)情况下, 总体上温度对SOS的影响依然最大(R温度> R辐射>R夜间灯光>R降水), 其中辐射对郊区和城区的春季物候的影响差异较大为0.34, 表明辐射与城区和郊区SOS密切程度存在显著差异。城区温度对春季物候的影响最大(R温度= -0.58), 即SOS随温度的升高呈现提前的变化趋势, 并且温度与城区春季物候的关系比郊区更密切(R郊区温度= -0.51)。与此相反, 夜间灯光对春季物候的影响呈现正相关关系(R夜间灯光= 0.22), 即夜间灯光越大春季物候越延迟, 夜间灯光对城区春季物候的影响大于郊区(R郊区夜间灯光=0.17, R城区夜间灯光= 0.27)。这与光照条件越好春季物候开始越早的结论相矛盾[41], 可能原因是夜间灯光数据为年度数据同时影响春季物候和秋季物候, 而秋季物候与下一年的春季物候正相关[42], 导致夜间灯光增大引起的SOS提前, 与夜间灯光增大引起的EOS越延迟和进一步SOS延迟抵消, 总体表现为夜间灯光增大SOS延迟。
图 4可知, 2001—2020年总体上温度、降水、辐射和夜间灯光对郊区的秋季物候影响作用大于城区, 尤其是辐射。具体表现为夜间灯光对EOS的影响作用最大(R夜间灯光=0.28), 其次为降水(R降水= 0.13), 温度的影响最小(R温度=0.05)。从REOS驱动因子分布曲线图可知, 城区和郊区辐射对EOS的影响存在显著差异, 城区和郊区温度对EOS的影响差异最小。与春季物候相比温度对秋季物候的影响显著下降, 并且降水对秋季物候的影响高于温度, 这与前人的研究一致[43]。

|
| 图 4 城区和郊区秋季物候与驱动因子的偏相关系数空间及频次分布曲线 Fig. 4 Spatial and frequency distribution curve of partial correlation coefficients between urban and suburban autumn phenology and driving factors |
仅考虑显著像元(P < 0.05), 秋季夜间灯光对EOS的影响最高(R夜间灯光>R降水>R辐射>R温度), 夜间灯光显著高于其他环境因子(R夜间灯光=0.54, R降水=0.38)。与春季物候相似辐射对秋季物候的影响存在较大城区和郊区差异(R郊区辐射- R城区辐射=0.38), 其次是降水(R郊区降水- R城区降水=0.25)。从偏相关分析结果的空间分布格局可知, 降水、辐射和夜间灯光对秋季物候的影响浦东新区和奉贤区与青浦区和松江区存在显著差异。如: 城区秋季物候与降水和辐射的负相关像元, 以及夜间灯光正相关的像元主要分布在在浦东新区和奉贤区(西部环)。可能与2006年提出的上海市五大新城的发展规划有关, 其中青浦新城、松江新城月嘉定新城是重点发展的新城, 而浦东新区在外环的南汇新城且距离主城较远发展较缓。因此, 区域差异引起植被物候对生长环境的响应存在一定的差异性。
2.4 植被物候主导驱动因子图 5所示, 2001—2020年总体上辐射和夜间灯光分别对SOS和EOS的影响面积(范围)最大, 约占研究区面积的40.42%和37.03%。城区春季物候和秋季物候的主导影响因子与总体类似, 仅主导因子占总面积的比例发生变化, 主要原因是城区占总面积的70%以上, 并且该辐射数据直接表示了植被冠层对能量的吸收能力, 影响叶绿素的形成、植被光合作用潜力等植被生长发育关键环节[44]。相比之下, 郊区春季物候和秋季物候的主要影响因子均为夜间灯光, 其百分比分别为40.23%和42.74%, 表明夜间灯光表示的人类活动强度为物候主导因子的面积高于其他环境驱动因子。此外, 温度和降水对城区和郊区植被物候的主导影响面积低于辐射和夜间灯光, 其中VIP温度+降水占研究区面积的百分比小于36.58%, 尤其是春季物候VIP辐射+夜间灯光占总面积的71.88%—72.65%左右。降水对春季和秋季物候的影响范围高于温度尤其是城区, 可能原因城市下垫面性质不仅改变热量平衡也改变地表水量平衡, 虽然城市利于降水的形成但由于地表性质, 使城市湿度低于郊区, 从而使得降水作为物候主导影响因子所占的面积高于温度作为主导因子所占面积。

|
| 图 5 城区和郊区植被物候主导因子分析 Fig. 5 Analysis of dominant factors of vegetation phenology in urban and suburban areas |
由于研究时段、城市、数据和分析方法的差异, 2001—2020年城区和郊区的温度、降水、辐射和夜间灯光及其对春季物候和秋季物候的响应具有明显差异(图 1)。如: 距离城市中心越近春季物候开始越早(城区春季物候早郊区约7d), 秋季物候结束越晚(秋季物候晚郊区约10d)。城区温度高于郊区温度约0.63℃, 城区夜间平均灯光高于郊区约2.9倍, 城区辐射低于郊区。这些结果与其他相关研究基本一致, 即城区和郊区植被生长环境存在显著差异, 城市热岛效应影响植被生理休眠和生态休眠需要的冷激和积温速率, 促使植被春季提前和秋季延迟[45—47]。然而由于城区的平均纬度低于郊区, 导致城区的春季物候提前和秋季物候延迟是纬度地带性和城市局地小气候共同作用的结果, 因此采用同纬度城区和郊区SOS和EOS平均值随纬度的变化(间隔0.004°)进行分析(图 6)。该结果与上述结论一致, 即城区平均春季物候早于郊区物候, 城区秋季平均物候低于郊区。城区和郊区的平均SOS和EOS随纬度的变化可知, 城区春季和秋季物候随纬度的变化趋势较之郊区更加一致和稳定, 可能是由于城区和郊区同纬度面积较小尤其是郊区(52.21%的面积为长兴岛和横沙岛), 并且郊区植被地理环境、植被类型分布和植被本身功能形状的自我调节的不同, 导致郊区植被物候纬度地带性较弱。

|
| 图 6 城区和郊区物候随纬度变化趋势 Fig. 6 The change trend of phenological of urban and suburban with latitude |
植被物候驱动因子的偏相关分析结果表明温度与城区和郊区春季物候相关性高于其他环境因子, 并且春季物候随温度的升高而提前, 这与前人的研究结果一致[48], 主要原因是研究区植被对热量的要求高, 并且温度影响解除生态休眠所需要热量或积温高于阈值(温度累积量)的积累[49]。夜间灯光对城区和郊区的秋季物候影响高于其他环境驱动因子, 可能原因是夜间灯光不仅可以指示人类活动对温度、CO2浓度、水热耦合模式以及废气/物等信息的影响强度[50—51], 也包含对植被生长具有重要的作用的光照强度[34—35], 夜间灯光通过直接影响植被的生长环境延长光照时间和光强增加植被光合作用糖类的累积量, 从而促使秋季物候的延迟[42]。与春季物候相比, 秋季物候对温度的响应相对较弱, 而对郊区的夜间灯光响应更强, 可能原因是城区热岛效应是植被的主要限制因素, 此外秋季物候的驱动机理更为复杂(如: 温度升高导致干旱, 引起脱落酸的形成导致植被提前脱落), 通常需要协同其他环境控制秋季物候[52—53]。
基于地理探测器分析和变量投影重要性指标得到的植被物候驱动因子的结果存在一定相似和差异, 其中上海市春季物候时间维度主要受温度的影响, 空间维度主要受夜间灯光的影响, 秋季物候时间维度主要受夜间灯光的影响, 空间维度主要受辐射的影响。该结论与东南沿海地区夜间灯光对植被动态变化解释率高于其他环境因子结论相一致[54], 即夜间灯光强度指示人类活动强度直接影响植被生长条件的下垫面(不透水面), 从而影响植被生长发育, 此外, 夜间灯光延长光照影响植被对低温的需求和脱落酸的分泌促进/延迟春季物候和秋季物候[55—56]。分析方法角度可能原因包括以下三个方面, 首先, 地理探测器分析方法是针对上海市植被物候空间分异, 探索其空间变化驱动因子的一种统计方法[40]。变量投影重要性指标是利用偏最小二乘提取驱动因子特征信息剔除共线信息[57], 从而确定驱动因子对植被物候的影响值。其次, 地理探测器分析采用的是2001—2020年的逐像元平均物候期和驱动因子, 对空间维度进行计算和分析[58], 而变量重要性指标则是对时间维度进行逐像元VIP计算, 并在空间维度上进行统计分析, 因此地理探测器分析方法对时间维度响应低于变量投影重要性指标。最后, 植被物候的驱动机理复杂, 受多个因子共同作用[3, 49], 导致不同方法对植被物候的主导驱动因子分析结果存在差异。
4 结论本研究利用上海市2001—2020年的温度、降水、辐射和夜间灯光探索人类活动因子和环境因子对植被物候(春季和秋季物候)的影响。首先, 采用上海城区和郊区的植被物候及其驱动因子的重心偏移情况以及年平均值进行时空变化分析; 然后, 利用地理探测器方法揭示植被物候空间维度的城市和郊区人类活动因子和环境因子对植被物候的影响强度及其交互作用; 最后, 结合偏相关分析和变量投影重要性指标对时间维度的城市和郊区植被物候驱动因子进行定量讨论。研究结果表明: 2001—2020年上海市城区和郊区植被物候及其驱动因子存在较大差异, 其中城区温度高于郊区约0.63℃, 春季物候提前郊区10d左右, 秋季物候推迟郊区7d左右, 夜间灯光高于郊区2.9倍。植被物候及其驱动因子重心均分布在宝山区范围内, 并且发生不同程度的偏移, 其中夜间灯光的重心偏移距离最大, 有明显的向沿海方向偏移。空间尺度上夜间灯光对SOS影响权重高于辐射、温度和降水, 其中城区SOS的影响权重最大(q=0.26), 并且辐射∩夜间灯光的组合驱动对城区和郊区的植被物候影响权重高于其他因子组合。时间尺度上SOS随温度的增加而提前(平均R温度=-0.24), 并且城区春季物候的主导影响因子是辐射(占总面积的41.40%), 而郊区春物物候的主导因子是夜间灯光(40.23%)。EOS随夜间灯光的增加而延迟(R夜间灯光=0.22), 城区和郊区EOS的主导因子均为夜间灯光, 其占总面积的百分比为35.79%和42.74%。本文的研究结果将有助于深入理解植被物候对气候变化的时空变化响应, 也为研究人类活动对植被物候的影响研究提供了一种新思路。
| [1] |
刘焱序, 吴文恒, 温晓金, 张东海. 晋陕蒙能源区城镇化过程及其对生态环境的影响. 地理研究, 2013, 32(11): 2009-2020. |
| [2] |
陈泓瑾, 刘琳, 张正勇, 刘亚, 田浩, 康紫薇, 王统霞, 张雪莹. 天山北坡人类活动强度与地表温度的时空关联性. 地理学报, 2022, 77(5): 1244-1259. |
| [3] |
Piao S, Liu Q, Chen A, Janssens I A, Fu Y, Dai J, Liu L, Lian X, Shen M, Zhu X. Plant phenology and global climate change: current progresses and challenges. Global Change Biology, 2019, 25(6): 1922-1940. DOI:10.1111/gcb.14619 |
| [4] |
丛楠, 张扬建, 朱军涛. 北半球中高纬度地区近30年植被春季物候温度敏感性. 植物生态学报, 2022, 46(2): 125-135. |
| [5] |
Richardson AD, Black TA, Ciais P, Delbart N, Friedl MA, Gobron N, Hollinger DY, Kutsch WL, Longdoz B, Luyssaert S, Migliavacca M, Montagnani L, Munger JW, Moors E, Piao S, Rebmann C, Reichstein M, Saigusa N, Tomelleri E, Vargas R, Varlagin A. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philosophical Transactions: Biological Sciences, 2010, 365(1555): 3227-3246. DOI:10.1098/rstb.2010.0102 |
| [6] |
Fu YH, Piao S, Delpierre N, Hao F, Hänninen H, Liu Y, Sun W, Janssens I A, Campioli M. Larger temperature response of autumn leaf senescence than spring leaf-out phenology. Global Change Biology, 2018, 24(5): 2159-2168. DOI:10.1111/gcb.14021 |
| [7] |
Stucky B J, Guralnick R, Deck J, Denny E G, Bolmgren K, Walls R. The plant phenology ontology: a new informatics resource for large-scale integration of plant phenology data. Frontiers in Plant Science, 2018, 9: 1-12. DOI:10.3389/fpls.2018.00001 |
| [8] |
付永硕, 张晶, 吴兆飞, 陈首志. 中国植被物候研究进展及展望. 北京师范大学学报(自然科学版), 2022, 58(3): 424-433. |
| [9] |
王宏, 李晓兵, 李霞, 莺歌, 符娜. 基于NOAA NDVI和MSAVI研究中国北方植被生长季变化. 生态学报, 2007, 27(2): 504-515. |
| [10] |
郑景云, 葛全胜, 郝志新. 气候增暖对我国近40年植物物候变化的影响. 科学通报, 2002, 47(20): 1582-1587. |
| [11] |
Song Z, Song X, Pan Y, Dai K, Shou J, Chen Q, Huang J, Tang X, Huang Z, Du Y. Effects of winter chilling and photoperiod on leaf-out and flowering in a subtropical evergreen broadleaved forest in China. Forest Ecology and Management, 2020, 458: 117766. DOI:10.1016/j.foreco.2019.117766 |
| [12] |
Chuine I, Bonhomme M, Legave J M, García de Cortázar-Atauri I, Charrier G, Lacointe A, Améglio T. Can phenological models predict tree phenology accurately in the future? The unrevealed hurdle of endodormancy break. Global Change Biology, 2016, 22(10): 3444-3460. DOI:10.1111/gcb.13383 |
| [13] |
Delpierre N, Vitasse Y, Chuine I, Guillemot J, Bazot S, Rutishauser T, Rathgeber C B K. Temperate and boreal forest tree phenology: from organ-scale processes to terrestrial ecosystem models. Annals of Forest Science, 2016, 73(1): 5-25. DOI:10.1007/s13595-015-0477-6 |
| [14] |
Davy R, Esau I, Chernokulsky A, Outten S, Zilitinkevich S. Diurnal asymmetry to the observed global warming. International Journal of Climatology, 2017, 37: 79-93. DOI:10.1002/joc.4688 |
| [15] |
Fu Y H, Zhao H, Piao S, Peaucelle M, Peng S, Zhou G, Ciais P, Huang M, Menzel A, Peñuelas J, Song Y, Vitasse Y, Zeng Z, Janssens I A. Declining global warming effects on the phenology of spring leaf unfolding. Nature, 2015, 526(7571): 104-107. DOI:10.1038/nature15402 |
| [16] |
Vitasse Y, Signarbieux C, Fu YH. Global warming leads to more uniform spring phenology across elevations. Proceedings of the National Academy of Sciences, 2018, 115(5): 1004-1008. DOI:10.1073/pnas.1717342115 |
| [17] |
Wareing P F. Photoperiodism in woody plants. Annual Review of Plant Physiology, 1956, 7: 191-214. DOI:10.1146/annurev.pp.07.060156.001203 |
| [18] |
Flynn D B, Wolkovich E M. Temperature and photoperiod drive spring phenology across all species in a temperate forest community. New Phytologist, 2018, 219(4): 1353-1362. DOI:10.1111/nph.15232 |
| [19] |
Tao Z, Wang H, Liu Y, Xu Y, Dai J. Phenological response of different vegetation types to temperature and precipitation variations in Northern China during 1982-2012. International Journal of Remote Sensing, 2017, 38(11): 3236-3252. DOI:10.1080/01431161.2017.1292070 |
| [20] |
Liu Q, Fu Y H, Zeng Z, Huang M, Li X, Piao S. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China. Global Change Biology, 2016, 22(2): 644-655. DOI:10.1111/gcb.13081 |
| [21] |
Hufkens K, Keenan T F, Flanagan L B, Scott R L, Bernacchi C J, Joo E, Brunsell N, Verfaillie J, Richardson A D. Productivity of North American grasslands is increased under future climate scenarios despite rising aridity. Nature Climate Change, 2016, 6(7): 710-714. DOI:10.1038/nclimate2942 |
| [22] |
Piao S, Yin G, Tan J, Cheng L, Huang M, Li Y, Liu R, Mao J, Myneni R B, Peng S, Poulter B, Shi X, Xiao Z, Zeng N, Zeng Z, Wang Y. Detection and attribution of vegetation greening trend in China over the last 30 years. Global Change Biology, 2015, 21(4): 1601-1609. DOI:10.1111/gcb.12795 |
| [23] |
Chen C, Park T, Wang X, Piao S, Xu B, Chaturvedi R K, Fuchs R, Brovkin V, Ciais P, Fensholt R, Tømmervik H, Bala G, Zhu Z, Nemani R R, Myneni R B. China and India lead in greening of the world through land-use management. Nature Sustainability, 2019, 2(2): 122-129. DOI:10.1038/s41893-019-0220-7 |
| [24] |
Liu H, Jiao F, Yin J, Li T, Gong H, Wang Z, Lin Z. Nonlinear relationship of vegetation greening with nature and human factors and its forecast-A case study of Southwest China. Ecological Indicators, 2020, 111: 106009. DOI:10.1016/j.ecolind.2019.106009 |
| [25] |
Piao S, Fang J, Ciais P, Peylin P, Huang Y, Sitch S, Wang T. The carbon balance of terrestrial ecosystems in China. Nature, 2009, 458(7241): 1009-1013. DOI:10.1038/nature07944 |
| [26] |
Li D, Stucky B J, Deck J, Baiser B, Guralnick R P. The effect of urbanization on plant phenology depends on regional temperature. Nature Ecology & Evolution, 2019, 3(12): 1661-1667. |
| [27] |
Walker J J, de Beurs K M, Henebry G M. Land surface phenology along urban to rural gradients in the U.S. Great Plains. Remote Sensing of Environment, 2015, 165: 42-52. DOI:10.1016/j.rse.2015.04.019 |
| [28] |
范良贤. 全球保护区和灯光区植被物候变化特征及影响因子研究[D]. 东北师范大学, 2020.
|
| [29] |
陈效逑, 王林海. 遥感物候学研究进展. 地理科学进展, 2009, 28(1): 33-40. |
| [30] |
Ditmer M A, Stoner D C, Francis C D, Barber J R, Forester J D, Choate D M, Ironside K E, Longshore K M, Hersey K R, Larsen R T, McMillan B R, Olson D D, Andreasen A M, Beckmann J P, Messmer T A, Carter N H. Artificial nightlight alters the predator-prey dynamics of an apex carnivore. Ecography, 2021, 44(2): 149-161. DOI:10.1111/ecog.05251 |
| [31] |
Speißer B, Liu Y, van Kleunen M. Biomass responses of widely and less-widely naturalized alien plants to artificial light at night. Journal of Ecology, 2021, 109(4): 1819-1827. DOI:10.1111/1365-2745.13607 |
| [32] |
Bennie J, Davies T W, Cruse D, Bell F, Gaston K J. Artificial light at night alters grassland vegetation species composition and phenology. Journal of Applied Ecology, 2018, 55(1): 442-450. DOI:10.1111/1365-2664.12927 |
| [33] |
Liu Y, Speißer B, Knop E, van Kleunen M. The Matthew effect: common species become more common and rare ones become more rare in response to artificial light at night. Global Change Biology, 2022, 28(11): 3674-3682. DOI:10.1111/gcb.16126 |
| [34] |
Koen E L, Minnaar C, Roever C L, Boyles J G. Emerging threat of the 21st century lightscape to global biodiversity. Global Change Biology, 2018, 24(6): 2315-2324. DOI:10.1111/gcb.14146 |
| [35] |
Gaston K J, Duffy J P, Bennie J. Quantifying the erosion of natural darkness in the global protected area system. Conservation Biology: the Journal of the Society for Conservation Biology, 2015, 29(4): 1132-1141. DOI:10.1111/cobi.12462 |
| [36] |
Gong H, Xiang X, Liu H, Xu X, Jiao F, Lin Z. 1 km monthly precipitation and temperatures dataset for China from 1952 to 2019 based on a brand-new and high-quality baseline climatology surface. Earth System Science Data, 2022, 1-30. |
| [37] |
王保林, 王晶杰, 杨勇, 常书娟, 陈喜梅, 刘爱军. 植被光合有效辐射吸收分量及最大光能利用率算法的改进. 草业学报, 2013, 22(5): 220-228. |
| [38] |
王正兴, 王亚琴. 利用LDOPE工具解码MODIS陆地产品质量信息. 遥感技术与应用, 2013, 28(3): 459-466. |
| [39] |
Wu Y, Shi K, Chen Z, Liu S, Chang Z. Developing improved time-series DMSP-OLS-like data (1992-2019) in China by integrating DMSP-OLS and SNPP-VIIRS. IEEE Transactions on Geoscience and Remote Sensing, 2022, 60: 1-14. |
| [40] |
王劲峰, 徐成东. 地理探测器: 原理与展望. 地理学报, 2017, 72(1): 116-134. |
| [41] |
张峰, 周广胜, 王玉辉. 内蒙古克氏针茅草原植物物候及其与气候因子关系. 植物生态学报, 2008, 32(6): 1312-1322. |
| [42] |
Shen M, Jiang N, Peng D, Rao Y, Huang Y, Fu Y H, Yang W, Zhu X, Cao R, Chen X, Chen J, Miao C, Wu C, Wang T, Liang E, Tang Y. Can changes in autumn phenology facilitate earlier green-up date of northern vegetation?. Agricultural and Forest Meteorology, 2020, 291: 108077. |
| [43] |
刘璐, 郭梁, 王景红, 栾青, 傅玮东, 李曼华. 中国北方苹果主产地苹果物候期对气候变暖的响应. 应用生态学报, 2020, 31(3): 845-852. |
| [44] |
Epiphanio J N, Huete A R. Dependence of NDVI and SAVI on Sun/sensor geometry and its effect on fAPAR relationships in Alfalfa. Remote Sensing of Environment, 1995, 51(3): 351-360. |
| [45] |
李静, 张彩琴, 张军. 内蒙古典型草原黄囊苔草生长动态对气象因子的滞后响应分析. 草地学报, 2017, 25(2): 267-272. |
| [46] |
Yao R, Wang L, Huang X, Guo X, Niu Z, Liu H. Investigation of urbanization effects on land surface phenology in northeast China during 2001-2015. Remote Sensing, 2017, 9(1): 66. |
| [47] |
胡召玲, 戴慧, 侯飞, 李二珠. 中国东北城乡植被物候时空变化及其对地表温度的响应. 生态学报, 2020, 40(12): 4137-4145. |
| [48] |
Myneni R B, Keeling C D, Tucker C J, Asrar G, Nemani R R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature, 1997, 386(6626): 698-702. |
| [49] |
付永硕, 李昕熹, 周轩成, 耿晓君, 郭亚会, 张雅茹. 全球变化背景下的植物物候模型研究进展与展望. 中国科学: 地球科学, 2020, 50(9): 1206-1218. |
| [50] |
岳文泽, 徐建华. 上海市人类活动对热环境的影响. 地理学报, 2008, 63(3): 247-256. |
| [51] |
沈建国, 白美兰, 李云鹏. 气候变化和人类活动对内蒙古生态环境的影响. 自然灾害学报, 2006, 15(6): 84-91. |
| [52] |
Estrella N, Menzel A. Responses of leaf colouring in four deciduous tree species to climate and weather in Germany. Climate Research, 2006, 32: 253-267. |
| [53] |
Tao Z, Wang H, Dai J, Alatalo J, Ge Q. Modeling spatiotemporal variations in leaf coloring date of three tree species across China. Agricultural and Forest Meteorology, 2018, 249: 310-318. |
| [54] |
陈文裕, 夏丽华, 徐国良, 余世钦, 陈行, 陈金凤. 2000-2020年珠江流域NDVI动态变化及影响因素研究. 生态环境学报, 2022, 31(7): 1306-1316. |
| [55] |
Way D A, Montgomery R A. Photoperiod constraints on tree phenology, performance and migration in a warming world. Plant Cell and Environment, 2015, 38(9): 1725-1736. |
| [56] |
Lang W, Chen X, Qian S, Liu G, Piao S. A new process-based model for predicting autumn phenology: how is leaf senescence controlled by photoperiod and temperature coupling?. Agricultural and Forest Meteorology, 2019, 268: 124-135. |
| [57] |
孙杨, 许承明, 夏锐. 研发资金投入渠道的差异对科技创新的影响分析——基于偏最小二乘法的实证研究. 金融研究, 2009(9): 165-174. |
| [58] |
吴克华, 苏维词, 贾真真, 王慧澄, 叶仕安, 罗时琴. 基于GIS与地理探测器的旅游地空间分布格局及驱动力分析——以贵州为例. 地理科学, 2022, 42(5): 841-850. |