 2023, Vol. 43
2023, Vol. 43文章信息
- 郭兴健, 邵全琴
- GUO Xingjian, SHAO Quanqin
- 大型野生食草动物生境资源利用竞争关系
- Population and habitat resource utilization of large wild herbivores based on UAV remote sensing
- 生态学报. 2023, 43(20): 8671-8681
- Acta Ecologica Sinica. 2023, 43(20): 8671-8681
- http://dx.doi.org/10.20103/j.stxb.202303030389
-
文章历史
- 收稿日期: 2023-03-03
- 采用日期: 2023-07-21

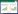




2. 陕西省地表系统与环境承载力重点实验室, 西安 710127;
3. 中国科学院地理科学与资源研究所, 陆地表层格局与模拟院重点实验室, 北京 100101;
4. 中国科学院大学, 北京 100049
2. Shaanxi Key Laboratory of Earth Surface System and Environmental Carrying Capacity, Xi'an 710127, China;
3. Key Laboratory of Land Surface Patterns and Simulation, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China;
4. University of Chinese Academy of Sciences, Beijing 100049, China
三江源位于青藏高原, 是长江、黄河和澜沧江的源区, 因特殊的自然地理格局与生态系统状况, 是我国极为重要的生态安全屏障[1], 发挥着包括生物多样性保护在内的诸多重要生态作用[2]。然而, 由于过度放牧等人类不合理活动对该地区脆弱生态系统造成的破坏, 三江源区生态系统生物多样性维持功能显著下降。尤其是各类大型野生食草动物, 在上世纪由于人类活动的干扰, 种群数量及分布区域在过度捕猎、草原退化、灾害等影响下不断缩减。我国高度重视三江源区生态退化的问题, 并于2005年批准了《青海三江源自然保护区生态保护和建设总体规划》。通过规划中一期工程的实施, 区内过度放牧得到了有效遏制, 草地载蓄压力明显下降[3], 野生动物种群数量有所恢复。2016年, 为进一步保护和恢复三江源区的生态环境及生物多样性, 我国建立了第一个国家公园——三江源国家公园, 与发达国家建立的国家公园相同, 野生动物保护是三江源国家公园的重要议题之一[4—5]。
为了准确掌握三江源区内大型野生动物恢复状况, 合理规划与科学动态调整保护区范围, 识别野生动物保护重点区域, 需要对区内野生动物的栖息地状况开展调查研究。野生动物调查最常用的传统方法是截线取样法[6—7], 即在调查区域内所有生境中随机地设置调查样线, 记录观察到的实体相对于样线的位置信息, 并结合探测函数来表示发现动物的概率与人跟动物距离的关系[8], 从而较高精度的进行种群数量估计。该方法在种群数量调查方面具有优势, 但生境调查中需要以发现野生动物活动痕迹、粪便或实体等处为中心设置样方, 采集动物痕迹样本和食草样本等[9]以获取对应的环境因子信息, 费时费力且成本较高[10]。
无人机遥感方法的出现为野生动物生境调查提供了一种全新的手段。虽然由于动物的警戒行为[11]及在家域范围内的移动[12—13], 基于无人机的种群数量估算存在不确定性[14], 但无人机遥感数据可实时准确记录动物空间位置信息的特点使其可在地理信息系统的支持下与各环境因子[15—16]进行空间叠加分析, 在生境调查研究中具有独特优势, 可以准确识别野生动物生境, 为保护区规划和研究提供基础, 判断保护区规划的合理性、生境斑块的连通性[17]、气候变化和人类活动干扰背景下栖息地范围的变化等[18—19]。近年来, 越来越多的研究人员尝试着采用这项新的技术手段开展野生动物调查研究, 包括白尾鹿(Odocoileus virginianus)[20]、驯鹿(Rangifer tarandus)[21]等, 无人机遥感已经被证明是一种有效的野生动物调查手段。
目前针对大型野生动物的栖息地研究大多针对单一物种[22]。但在实际情况下, 同一区域内的多种相似食性的野生动物之间的栖息地资源和营养资源之间可能存在重叠与冲突关系[23], 即它们的生态位重叠。在野生动物保护规划中, 为了科学开展野生动物保护工作的综合规划和管理, 实现资源的合理配置与效益最大化, 各野生动物之间的生态位重叠关系是不可忽略的, 进一步明确主导这种关系的因素更是合理分配栖息地资源的关键。然而受限于大范围数据获取的限制, 此类研究成果较少。
基于此, 本研究选取藏野驴(Asinus kiang)、藏原羚(Procapra picticaudata)和岩羊(Pseudois nayaur)三种青藏高原特有, 同时也是我国二级保护动物的大型野生食草动物作为研究对象, 在三江源区玛多县境内开展无人机调查研究, 通过目标动物解译结果与各类环境因子的空间叠加分析, 预期得到:(1)玛多县境内藏野驴、藏原羚及岩羊在样带内的分布特征; (2)三种大型野生食草动物的栖息地选择偏好及适宜栖息地范围; (3)藏野驴、藏原羚及岩羊各自的生态位, 互相之间的资源利用重叠与冲突程度及主导因素。本研究将综合利用3S技术, 形成完整的野生动物生境范围识别及限制因素分析以及多物种生境资源利用冲突的研究方法, 相关成果可为青藏高原区域野生动物栖息地监测提供新方法, 为国家公园与自然保护区内多物种综合保护规划提供支持与参考。
1 材料与方法 1.1 研究区概况玛多县位于青海省果洛藏族自治州内, 地理位置介于东经96°50′—99°20′, 北纬33°50′—35°40′, 全县土地面积约25300 km2, 海拔范围在3902—5243 m之间, 地形起伏不大, 相对平坦。县域内河湖密布, 其中扎陵湖与鄂陵湖为黄河源头。由于人类活动和气候变化的双重影响, 上世纪末期玛多县内草地退化、湿地面积萎缩、野生动物数量大幅减少, 曾是三江源区生态系统退化最严重的县份之一。自2005年起, 三江源自然保护区生态保护和建设工程的实施有效遏制了生态系统的退化, 区内野生动物的种群数量明显恢复。2016年我国在三江源建立国家公园, 其中黄河源园区就位于玛多县境内。作为生态治理与野生动物保护的典型区域, 本研究选择玛多县作为研究区开展藏野驴、藏原羚和岩羊的生境分析研究(图 1)。

|
| 图 1 研究区概况 Fig. 1 Study Area |
本研究采用两款无人机搭载可见光传感器于2017年春季开展野外航拍调查工作, 调查样带遵循随机抽样的原则布设, 分布在扣除非植被类型区域(如裸岩石砾地、水体、冰川、城镇、高速和国道道路等)后的生境范围内。
航拍调查采用了两款无人机:第一款为成都山地所自主研发的电动无人机, 续航时间90 min, 飞行速度为72 km/h; 第二款为深圳飞马机器人科技有限公司F1000无人机, 续航时间60 min, 飞行速度为60 km/h(图 2)。2017年4月9日—18日, 在玛多县共计航拍有效架次14个, 拍摄高度200—350 m, 成图分辨率4—7 cm, 获取23784张影像, 有效拍摄面积326.6 km2。

|
| 图 2 航拍调查所用成都山地所无人机和飞马F1000无人机 Fig. 2 The UAVs of Institute of Mountain Hazards and Environment and Feima F1000 used in aerial surveys |
获取的无人机影像采用两台高性能工作站进行拼接, 并开展人工目视解译工作。影像拼接使用Pix4Dmapper、飞马智拼图与LiMapper软件。目视解译工作使用ArcGIS软件, 依据遥感解译七要素, 即色调、颜色、纹理、阴影、大小、形状、图案, 并结合野外调查中观察到的动物活动规律和习性产生的位置和分布规律进行, 航拍影像中的动物群体、个体及地面实际观察到的个体特征如图 3所示。

|
| 图 3 藏野驴、藏原羚和岩羊无人机航拍影像中群体、个体特征及地面拍摄外观形态 Fig. 3 The group and individual characteristics of Kiang, Tibetan gazelle and Blue sheep in UAV aerial photography and the appearance of ground |
本研究采用选择指数Ei来表示藏野驴、藏原羚和岩羊对环境因子的选择偏好或回避[24], 具体包括距农村居民点距离、距公路距离、距水源距离、坡度、植被覆盖度以及植被类型等6个环境因子, 计算方法如下:
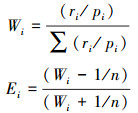
|
式中, Wi为选择系数, Ei为选择指数, i指某环境特征, ri为物种选择具有i特征的样方数, pi指环境中具有i特征的总样方数, n指某环境特征的等级数(n=1, 2, …n); Ei介于-1到1之间, 当Ei=1时为特别偏好, Ei=-1为不选择, Ei<-0.1为负选择, Ei>0.1为正选择, Ei=0为随机选择。-0.1≤Ei≤0.1为几乎随机选择。
该方法是基于地面调查样方提出的, 本研究中将无人机调查影像中每个30 m×30 m的栅格作为一个样方, 相当于共有调查样方356468个。若栅格范围内存在目标动物, 无论动物头只数多少, 均计为1个被选择样方。
1.4 藏野驴、藏原羚及岩羊生境重叠度及竞争分析方法本研究采用生境生态位宽度表示某种大型野生食草动物对某种环境因子的选择范围, 利用生态位重叠指数表征藏野驴、藏原羚和岩羊在对生境资源选择利用过程中的重叠及竞争程度。
生境生态位宽度通过Shannon-Wiener公式计算:
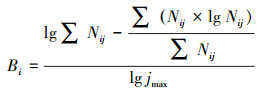
|
式中, Bi表示物种i(藏野驴、藏原羚或岩羊)在某种生境因子中的生境生态位宽; j为生境因子等级; jmax为生境因子等级数量, 本研究中为各环境因子的分级/分类个数; Nij为物种i利用某生境因子第j级的数量。
采用Pianka生态位重叠指数表征藏野驴、藏原羚和岩羊的生态位重叠程度, 计算公式如下:
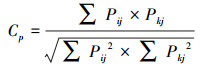
|
式中, Cp为物种i与物种k在某种生境因子上的生态位重叠指数; Pij为物种i利用某生境因子第j级的比例; Pkj为物种k利用某生境因子第j级的比例。Cp介于0—1之间, 0代表两种物种不存在重叠, 即生境完全不同, 1代表两种物种生境完全重叠, 及生境完全相同。
1.5 其他数据来源居民地、水源地与公路数据等来自2015年高分遥感影像解译得到的玛多县土地覆被数据, 由青海省生态环境遥感中心提供, 空间分辨率为2 m×2 m; 地面数字高程模型(DEM)数据来自星载热发射和反射辐射仪全球数字高程模型(ASTER GDEM), 坡度数据由DEM数据在ArcGIS软件中计算得到, 空间分辨率均为30 m×30 m; 草地类型矢量数据来自于中国1:100万植被类型数据集; 植被覆盖度数据由Landsat TM影像(NDVI)数据计算得到, 由中国测绘科学研究院提供, 空间分辨率为30 m×30 m。
2 结果 2.1 无人机样带调查解译结果结果显示, 14条样带中的9条发现藏野驴个体, 共计252只, 样带内平均密度为0.77只/km2; 6条样带中发现藏原羚个体, 共计发现199只, 样带内的平均种群密度为0.61只/km2; 2条样带内发现岩羊个体, 共计203只, 样带内平均密度为0.62只/km2(表 1)。从样带结果来看, 藏野驴分布范围最广, 藏原羚次之, 岩羊仅在玛多县东北部花石峡镇内地形陡峭区域的两条样带中被发现。
| 样带编号 Transect numbers |
藏野驴Kiang | 藏原羚Tibetan gazelle | 岩羊Blue sheep | |||||
| 个体数量/只 Number of individuals |
密度/(只/km2) Density |
个体数量/只 Number of individuals |
密度/(只/km2) Density |
个体数量/只 Number of individuals |
密度/(只/km2) Density |
|||
| 1 | 107 | 2.66 | 23 | 0.57 | 0 | 0 | ||
| 2 | 6 | 0.39 | 23 | 1.48 | 0 | 0 | ||
| 3 | 4 | 0.09 | 10 | 0.22 | 0 | 0 | ||
| 4 | 2 | 0.06 | 0 | 0 | 0 | 0 | ||
| 9 | 48 | 1.84 | 0 | 0 | 0 | 0 | ||
| 10 | 85 | 4.75 | 1 | 0.06 | 0 | 0 | ||
| 11 | 24 | 0.62 | 70 | 1.81 | 195 | 5.03 | ||
| 12 | 20 | 0.48 | 72 | 1.73 | 8 | 0.19 | ||
| 13 | 82 | 3.13 | 0 | 0 | 0 | 0 | ||
| 合计Total | 252 | 0.77 | 199 | 0.61 | 203 | 0.62 | ||
| 5—8号及14号样带中未发现目标野生动物个体 | ||||||||
生境选择的计算结果显示, 藏野驴偏好于选择距农村居民点1—3 km、距公路距离1—3 km、距水源地距离1 km以内、坡度小于2°且植被覆盖度介于0.4—0.8之间的植被类型为高山嵩草+杂类草以及高山嵩草+紫花针茅草地型的区域作为栖息地; 藏原羚喜好选择远离农村居民点3 km以上、距公路和水源地距离1—3 km、坡度介于5—10°、植被覆盖度大于0.6的藏嵩草草地型以及高山嵩草+杂类草草地型区域作为栖息地; 而岩羊则偏好于选择距农村居民点1 km以内、距公路3 km以上、距水源1—3 km、坡度大于5°、植被覆盖度介于0—0.4之间且植被类型为紫花针茅+杂类草以及高山嵩草草地型的区域作为栖息地(表 2)。
| 环境因子 Environmental factors |
环境因子分级 Environmental factors classification |
调查样方总数/个 Amount of survey quadrats |
物种选择样方数/个 Selected quadrats′ amount |
样方内总头只数/只 Amount of species in quadrat |
选择系数 Selection coefficient |
选择指数 Selection index |
选择性 Selection |
||||||||||||||
| 藏野驴 Kiang |
藏原羚 Tibetan gazelle |
岩羊 Blue sheep |
藏野驴 Kiang |
藏原羚 Tibetan gazelle |
岩羊 Blue sheep |
藏野驴 Kiang |
藏原羚 Tibetan gazelle |
岩羊 Blue sheep |
藏野驴 Kiang |
藏原羚 Tibetan gazelle |
岩羊 Blue sheep |
藏野驴 Kiang |
藏原羚 Tibetan gazelle |
岩羊 Blue sheep |
|||||||
| 距农村居民点距离/km | ≤1 | 21815 | 4 | 10 | 18 | 34 | 27 | 86 | 0.12 | 0.35 | 0.71 | -0.47 | 0.02 | 0.36 | NS | RS | PS | ||||
| Distance from rural | 1—3 | 66347 | 53 | 16 | 23 | 117 | 50 | 105 | 0.53 | 0.16 | 0.25 | 0.23 | -0.36 | -0.13 | PS | NS | NS | ||||
| settlements | >3 | 143114 | 76 | 43 | 3 | 101 | 122 | 12 | 0.35 | 0.50 | 0.04 | 0.03 | 0.20 | -0.79 | RS | PS | NS | ||||
| 距公路距离/km | ≤1 | 6178 | 0 | 9 | 1 | 0 | 23 | 2 | 0.00 | 0.37 | 0.14 | -1.00 | 0.05 | -0.40 | NOS | RS | NS | ||||
| Distance from road | 1—3 | 27642 | 27 | 22 | 2 | 131 | 50 | 5 | 0.65 | 0.44 | 0.14 | 0.32 | 0.14 | -0.41 | PS | PS | NS | ||||
| >3 | 197456 | 106 | 38 | 41 | 121 | 126 | 196 | 0.35 | 0.19 | 0.72 | 0.03 | -0.28 | 0.36 | RS | NS | PS | |||||
| 距水源距离/km | ≤1 | 206250 | 128 | 42 | 21 | 236 | 125 | 120 | 0.75 | 0.25 | 0.16 | 0.39 | -0.15 | -0.35 | PS | NS | NS | ||||
| Distance from water | 1—3 | 24744 | 5 | 27 | 23 | 16 | 74 | 83 | 0.25 | 0.75 | 0.84 | -0.15 | 0.39 | 0.43 | NS | PS | PS | ||||
| >3 | 282 | 0 | 0 | 0 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | -1.00 | -1.00 | -1.00 | NOS | NOS | NOS | |||||
| 坡度/(°) | ≤2 | 39317 | 43 | 2 | 2 | 82 | 8 | 2 | 0.54 | 0.07 | 0.04 | 0.24 | -0.66 | -0.80 | PS | NS | NS | ||||
| Slope | 2—5 | 96814 | 70 | 19 | 5 | 130 | 54 | 25 | 0.36 | 0.30 | 0.04 | 0.03 | -0.05 | -0.77 | RS | RS | NS | ||||
| 5—10 | 38643 | 20 | 48 | 11 | 40 | 137 | 53 | 0.10 | 0.63 | 0.41 | -0.53 | 0.31 | 0.10 | NS | PS | PS | |||||
| >10 | 56502 | 0 | 0 | 29 | 0 | 0 | 123 | 0.00 | 0.00 | 0.52 | -1.00 | -1.00 | 0.22 | NOS | NOS | PS | |||||
| 植被覆盖度 | 0≤FVC < 0.2 | 42198 | 2 | 11 | 18 | 2 | 35 | 77 | 0.02 | 0.10 | 0.61 | -0.84 | -0.33 | 0.51 | NS | NS | PS | ||||
| FVC | 0.2≤FVC < 0.4 | 128686 | 78 | 27 | 24 | 148 | 75 | 120 | 0.22 | 0.10 | 0.31 | 0.06 | -0.35 | 0.22 | RS | NS | PS | ||||
| 0.4≤FVC < 0.6 | 37099 | 40 | 15 | 2 | 68 | 52 | 6 | 0.40 | 0.16 | 0.08 | 0.33 | -0.11 | -0.44 | PS | NS | NS | |||||
| 0.6≤FVC < 0.8 | 13950 | 12 | 9 | 0 | 22 | 21 | 0 | 0.32 | 0.29 | 0.00 | 0.23 | 0.19 | -1.00 | PS | PS | NOS | |||||
| 0.8≤FVC < 1 | 9343 | 1 | 7 | 0 | 12 | 16 | 0 | 0.04 | 0.35 | 0.00 | -0.67 | 0.27 | -1.00 | NS | PS | NOS | |||||
| 植被类型 | 高山嵩草+杂类草草地型 | 35662 | 61 | 7 | 0 | 107 | 23 | 0 | 0.33 | 0.14 | 0.00 | 0.49 | 0.11 | -1.00 | PS | PS | NOS | ||||
| Vegetation type | 高山嵩草+紫花针茅草地型 | 13013 | 42 | 1 | 0 | 71 | 1 | 0 | 0.62 | 0.05 | 0.00 | 0.69 | -0.34 | -1.00 | PS | NS | NOS | ||||
| 禾叶风毛菊草地型 | 3949 | 0 | 0 | 0 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | -1.00 | -1.00 | -1.00 | NOS | NOS | NOS | |||||
| 紫花针茅+杂类草草地型 | 107698 | 28 | 15 | 29 | 72 | 46 | 139 | 0.05 | 0.08 | 0.35 | -0.38 | -0.19 | 0.52 | NS | NS | PS | |||||
| 藏嵩草+苔草草地型 | 10754 | 0 | 0 | 0 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | -1.00 | -1.00 | -1.00 | NOS | NOS | NOS | |||||
| 藏嵩草草地型 | 4105 | 0 | 0 | 0 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | -1.00 | -1.00 | -1.00 | NOS | NOS | NOS | |||||
| 高山嵩草草地型 | 3123 | 0 | 32 | 13 | 0 | 91 | 58 | 0.00 | 0.61 | 0.60 | -1.00 | 0.69 | 0.69 | NOS | PS | PS | |||||
| 赖草草地型 | 8189 | 0 | 0 | 0 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | -1.00 | -1.00 | -1.00 | NOS | NOS | NOS | |||||
| 紫花针茅草地型 | 44783 | 2 | 14 | 2 | 2 | 38 | 6 | 0.01 | 0.12 | 0.04 | -0.86 | 0.04 | -0.45 | NS | RS | NS | |||||
| PS:正选择Positive selection; RS:随机选择Random selection; NS:负选择Negative Selection; NOS:不选择No Selection; FVC:植被覆盖度Fractional Vegetation Cover | |||||||||||||||||||||
在实际情况下, 各环境因子并非相互独立, 譬如地形陡峭的区域通常距离公路较远, 因此藏野驴、藏原羚和岩羊对某些环境因子所表现出来的选择偏好可能存在被更加重要的环境因子所胁迫的情况。例如岩羊大多选择陡峭的悬崖峭壁作为生境[23], 悬崖峭壁下是否存在居民点对其影响有限, 造成对于距居民点1 km以内区域的栖息地选择偏好可信度下降。
为了解决上述问题, 明确影响各大型野生食草动物栖息地选择偏好的主要因子, 更加科学的判定优质栖息地范围, 对各环境因子在SPSS软件中进行了因子分析。三种野生动物因子分析的前2个公因子对于变异系数的累积解释度均超过了75%, 因此选择前2个公因子进行分析。
如表 3所示, 对于藏野驴, 公因子1中载荷绝对值较大的为距农村居民点距离、距公路距离, 公因子2中载荷绝对值较大的为距水源距离以及坡度; 对于藏原羚, 公因子1中载荷绝对值较大的为植被类型、植被覆盖度与距水源距离, 公因子2中载荷绝对值较大的为坡度; 对于岩羊, 公因子1中载荷绝对值较大的为距水源距离、坡度以及植被类型, 公因子2中载荷绝对值较大的为距公路距离。通过分析结果, 有理由相信上述各因子为野生动物栖息地选择时的主导因子。
| 动物种类 Animal species |
环境因子 Environmental factors |
公因子1 Factor 1 |
公因子2 Factor 2 |
| 藏野驴 | 距农村居民点距离 | -0.929 | -0.010 |
| Kiang | 距公路距离 | 0.911 | 0.228 |
| 距水源距离 | 0.078 | 0.858 | |
| 坡度 | 0.007 | 0.839 | |
| 植被覆盖度 | 0.076 | 0.025 | |
| 植被类型 | 0.695 | -0.370 | |
| 藏原羚 | 距农村居民点距离 | 0.075 | -0.567 |
| Tibetan gazelle | 距公路距离 | -0.046 | -0.688 |
| 距水源距离 | -0.712 | 0.374 | |
| 坡度 | -0.029 | 0.828 | |
| 植被覆盖度 | 0.721 | 0.226 | |
| 植被类型 | 0.806 | -0.057 | |
| 岩羊 | 距农村居民点距离 | 0.463 | 0.615 |
| Blue sheep | 距公路距离 | 0.152 | -0.901 |
| 距水源距离 | 0.900 | 0.246 | |
| 坡度 | 0.881 | 0.026 | |
| 植被覆盖度 | 0.057 | 0.667 | |
| 植被类型 | -0.817 | 0.060 |
根据藏野驴、藏原羚和岩羊的生境分析结果, 将各主导因子中的倾向选择(PS)以及随机选择(RS)区域定义为适宜生境。即藏野驴适宜生境为距离农村居民点及公路1 km以上, 距水源1 km以内的坡度小于5°的区域; 藏原羚的适宜生境为距水源1—3 km范围内, 坡度介于2—10°, 植被覆盖度大于0.6的高山嵩草+杂类草、高山嵩草以及紫花针茅草地区域; 岩羊的适宜生境为距公路3 km以外, 距水源1—3 km范围内, 坡度大于5°的紫花针茅+杂类草及高山嵩草草地区域。
通过与研究区各环境因子数据的空间叠加分析, 识别出了玛多县各大型食草动物的适宜生境范围, 结果如图 4所示。在玛多县县域范围内, 藏野驴的适宜生境面积最大, 为3559.43 km2, 广泛分布在整个县域内, 东北部分布密度稍低; 藏原羚的适宜生境面积次之, 为283.76 km2, 主要分布在玛多县中部及东南部; 岩羊的适宜生境面积最小, 仅为29.97 km2, 这可能与其生境选择中对地形坡度的严苛要求有关, 适宜生境主要分布在东北部山地区域。

|
| 图 4 玛多县藏野驴、藏原羚及岩羊适宜生境范围及重叠区域 Fig. 4 Suitable habitat area of Kiang, Tibetan gazelle and Blue sheep in Maduo County and overlapping areas with each other |
通过将三种动物的适宜栖息地进一步叠加分析, 发现虽然藏野驴的适宜栖息地范围最大, 但是其与藏原羚或者岩羊的栖息地重叠程度很小, 重叠面积均不足0.01 km2, 藏原羚与岩羊的适宜栖息地重叠面积则是6.04 km2, 仅占到了藏原羚适宜栖息地的2.12%。整体上, 三种野生大型食草动物生境重叠程度均较弱。
2.3 藏野驴、藏原羚及岩羊的生境资源竞争分析通过对三种目标研究动物的生境生态位宽度(Bi)及生态位重叠程度(Cp)的定量计算, 更直观的反映了三种野生食草动物在生境资源利用上的限制条件及相互冲突竞争关系(表 4)。
| 环境因子 Environmental factors |
Bi | Cp | |||||
| 藏野驴 Kiang |
藏原羚 Tibetan gazelle |
岩羊 Blue sheep |
藏野驴-藏原羚 Between Kiang and Tibetan gazelle |
藏野驴-岩羊 Between Kiang and Blue sheep |
藏原羚-岩羊 Between Tibetan gazelle and Blue sheep |
||
| 距农村居民点距离 Distance from rural settlements |
0.90 | 0.84 | 0.79 | 0.90 | 0.76 | 0.49 | |
| 距公路距离Distance from road | 0.63 | 0.81 | 0.16 | 0.89 | 0.70 | 0.92 | |
| 距水源距离Distance from water | 0.22 | 0.60 | 0.62 | 0.89 | 0.86 | 1.00 | |
| 坡度Slope | 0.91 | 0.67 | 0.55 | 0.56 | 0.08 | 0.26 | |
| 植被覆盖度FVC | 0.66 | 0.91 | 0.49 | 0.92 | 0.78 | 0.83 | |
| 植被类型Vegetation type | 0.51 | 0.59 | 0.33 | 0.36 | 0.45 | 0.71 | |
藏野驴在坡度及距农村居民点距离两个因子上的生态位宽度分别达到了0.91及0.90, 说明藏野驴对这两种环境因子的利用范围较大。而在距水源距离上的生态位宽度仅有0.22, 说明藏野驴在选择栖息地时距水源1 km以内的偏好导致其在该因子上的利用范围较小, 该因素是藏野驴栖息地范围的主要限制因子。
藏原羚在各环境因子上的生态位宽度更加均衡, 大多介于0.6—0.9之间。其中, 藏原羚在植被覆盖度上更广的选择偏好使得其在植被覆盖度因子上的生态位宽度达到了0.91。其在坡度、距水源距离以及植被类型上的生境生态位宽度相比之下较低, 分别为0.67、0.60和0.59, 说明藏原羚在选择栖息地时对上述三个因子的偏好要求成为制约栖息地面积的主要因素。
相比于藏野驴及藏原羚, 岩羊在各生境因子上的生态位宽度整体偏低。其中, 在距公路距离因子上的生境生态位宽度仅0.16, 在植被类型和坡度上分别仅0.36和0.55。说明岩羊对于上述因子的选择范围较窄, 岩羊这种对于环境因子的特定选择偏好范围可能是导致其适宜生境面积较小的主要原因。
三种大型野生食草动物之间的生态位重叠程度揭示了他们在生境资源利用上的竞争关系与程度。其中, 藏野驴与藏原羚在植被覆盖度因子上的重叠程度最高, 达到了0.92;在距农村居民点距离、距公路距离以及距水源距离因子上分别达到了0.90、0.89以及0.89;但在坡度因子上重叠程度较低, 为0.56。藏野驴与岩羊在距水源地距离因子上的重叠程度最高, 为0.86;而在坡度因子上的重叠程度仅为0.08。藏原羚和岩羊之间的生境因子重叠程度整体较高, 距水源距离、距公路距离及植被覆盖度分别达到了1.00、0.92和0.83;但坡度上较低, 为0.26。
结合对三种大型野生食草动物适宜生境范围重叠面积的分析结果, 三者在距农村居民点、公路及水源地的距离上, 以及植被盖度等的选择上重叠度均较高, 但整体生境重叠面积较小, 这是三种动物对坡度选择的差异造成的(坡度上的重叠度分别为0.56、0.08及0.26)。
3 讨论三江源区是我国重要的动植物资源宝库, 对于我国第一个国家公园——三江源国家公园来说, 生物多样性保护尤为重要。藏野驴、藏原羚及岩羊作为我国濒危野生动物, 是生物多样性保护工作中的主要对象。以国家公园为主体, 各项人为保护干预措施可以很大程度上的帮助区内野生动物种群数量恢复及生境质量提高[25]。为了该目的, 需要制定科学、合理、高效的管理手段, 而掌握野生动物的种群数量、分布以及生境状况是前提[26—27]。
本研究将无人机遥感技术应用于大范围的野生动物栖息地调查研究, 充分发挥了无人机快速便捷、受地形影响小、获取数据具有地理空间属性等优势, 为相关野生动物研究提供了全新的技术手段与研究思路[28]。在本研究中, 使用4—7 cm的无人机航拍图像, 可以清晰地识别出藏野驴、藏原羚与岩羊。因此, 建议在未来的大型野生食草动物无人机调查中, 图像分辨率不应低于7 cm, 最佳分辨率至少为5 cm左右。无人机航拍过程中我们设置了80%的航向重叠和60%的旁向重叠, 但是由于研究区域的特征点较少, 仍然存在图像拼接精度低的问题。这些结果表明, 在未来青藏高原或类似地区的研究中, 进一步增强图像重叠对提高拼接精度的作用有限, 应采用地面控制点放置等方法。此外, 影像拼接精度问题会导致成图中野生动物计数的准确性降低, 因此根据其进行种群数量估计时需要谨慎。但无人机遥感在栖息地监测中优势明显, 尤其是在青藏高原等地形平坦开阔, 面积广大的区域。但受限于无人机自上而下的平台视角, 在森林等遮挡严重的区域无法获取林下动物的影像, 这些区域更适用传统的样线法、痕迹法等[29]。
掌握野生动物的栖息地选择偏好, 划定合理的适宜栖息地范围, 给野生动物生存和繁衍提供优质的生境资源无疑会促进其种群数量恢复的快速实现。但值得注意的是, 国家公园在野生动物保护中需要进行综合管理, 这就包括多目标的平衡问题[30]。因此, 除了单一目标的生境识别, 需要对保护区内的各种习性或食性相近的野生动物进行生境资源竞争关系分析, 明确栖息地重叠状况, 并揭示主要原因。在本研究中, 受到坡度因子的主导, 三种大型野生食草动物虽然在其他因子上的生态位重叠程度较高, 但实际适宜栖息地的重叠程度很小。因此, 在玛多县县域内针对藏野驴、藏原羚及岩羊进行栖息地划定分配时应以坡度地形为主要依据。此外, 由于三江源区自然环境较为脆弱, 为保护生态, 草地的承载能力一直是备受关注的热点问题。已有研究综合考虑家畜及野生食草动物种群数量来估算草地载蓄压力[31]。未来在明确各野生动物栖息地选择及分布关系的基础上, 可以进一步更加科学的估算草地承载能力[32], 尤其是在多物种的生境重叠区域。
4 结论本研究利用无人机遥感对三江源玛多县藏野驴、藏原羚及岩羊的适宜生境开展范围研究, 并揭示各物种间的生境生态位关系和竞争情况, 取得以下结论:
(1) 无人机样带内发现藏野驴个体252只, 平均种群密度为0.77只/km2; 藏原羚个体199只, 平均种群密度为0.61只/km2; 岩羊个体203只, 平均种群密度为0.62只/km2。
(2) 藏野驴偏好选择距离农村居民点及公路1 km以上, 距水源1 km以内的坡度小于5°的区域作为生境; 藏原羚偏好选择距水源1—3 km范围内, 坡度介于2—10°, 植被覆盖度大于0.6的高山嵩草+杂类草、高山嵩草以及紫花针茅草地区域作为生境; 岩羊偏好选择距公路3 km以外, 距水源1—3 km范围内, 坡度大于5°的紫花针茅+杂类草及高山嵩草草地区域作为生境。
(3) 玛多县藏野驴适宜生境面积3559.43 km2, 主要受到距水源距离因子的制约; 藏原羚的适宜生境面积为283.76 km2, 主要受到其在坡度、距水源距离以及植被类型上选择偏好的制约; 岩羊的适宜生境面积为29.97 km2, 主要受到其在坡度、距公路距离和植被类型上的选择偏好制约。在栖息地坡度选择中的生态位差异, 是造成三种大型野生食草动物的生境重叠面积均较小, 生境竞争关系较弱的主要原因。
| [1] |
Xue J, Li Z X, Feng Q, Gui J, Zhang B J. Construction of ecological conservation pattern based on ecosystem services of Three River Headwaters, Western China. Global Ecology and Conservation, 2023, 44: e2491. |
| [2] |
孙鸿烈, 郑度, 姚檀栋, 张镱锂. 青藏高原国家生态安全屏障保护与建设. 地理学报, 2012, 67(1): 3-12. |
| [3] |
Shao Q Q, Cao W, Fan J W, Huang L, Xu X L. Effects of an ecological conservation and restoration project in the Three-River Source Region, China. Journal of Geographical Sciences, 2017, 27(2): 183-204. DOI:10.1007/s11442-017-1371-y |
| [4] |
陈东军, 钟林生, 樊杰, 虞虎, 杨定, 曾瑜皙. 青藏高原国家公园群功能评价与结构分析. 地理学报, 2022, 77(1): 196-213. |
| [5] |
郑德凤, 郝帅, 吕乐婷, 徐文瑾, 王燕燕, 王辉. 三江源国家公园生态系统服务时空变化及权衡-协同关系. 地理研究, 2020, 39(1): 64-78. |
| [6] |
Aryal A, Gastaur S, Menzel S, Chhetri T B, Hopkins J. Estimation of blue sheep population parameters in the Dhorpatan Hunting Reserve, Nepal. Academic Journals, 2010. |
| [7] |
初红军, 蒋志刚, 葛炎, 蒋峰, 陶永善, 王臣. 卡拉麦里山有蹄类自然保护区蒙古野驴和鹅喉羚种群密度和数量. 生物多样性, 2009, 17(4): 414-422. |
| [8] |
李欣海, 郜二虎, 李百度, 詹祥江. 用物种分布模型和距离抽样估计三江源藏野驴、藏原羚和藏羚羊的数量. 中国科学: 生命科学, 2019, 49(2): 151-162. |
| [9] |
张洪茂, 胡锦矗. 川西北高原藏原羚夏季生境选择. 四川动物, 2002, 21(1): 12-15. DOI:10.3969/j.issn.1000-7083.2002.01.004 |
| [10] |
邵全琴, 郭兴健, 李愈哲, 汪阳春, 王东亮, 刘纪远, 樊江文, 杨帆. 无人机遥感的大型野生食草动物种群数量及分布规律研究. 遥感学报, 2018, 22(3): 497-507. |
| [11] |
李忠秋. 三种珍稀有蹄类动物的警戒行为数据. 生物多样性, 2016, 24(12): 1335-1340. |
| [12] |
Gates C E, Burnham K P, Anderson D R, Laake J L. Estimation of density from line transect sampling of biological populations. Biometrics, 1981, 37(2): 413. |
| [13] |
Schweder T. Advanced distance sampling: estimating abundance of biological populations. Journal of the American Statistical Association, 2007, 102(478): 763-764. DOI:10.1198/jasa.2007.s192 |
| [14] |
郭兴健, 邵全琴, 杨帆, 李愈哲, 汪阳春, 王东亮. 无人机遥感调查黄河源玛多县岩羊数量及分布. 自然资源学报, 2019, 34(5): 1054-1065. |
| [15] |
Lian X M, Li X X, Zhou D X, Yan P S. Avoidance distance from Qinghai-Tibet Highway in sympatric Tibetan antelope and gazelle. Transportation Research Part D: Transport and Environment, 2012, 17(8): 585-587. DOI:10.1016/j.trd.2012.06.009 |
| [16] |
黄青东智, 陈刘阳, 李尚鹏, 孙章运, 杨明新, 张德喜, 李成先, 毛锐锐, 李泽宇, 张立勋. 道路对三江源国家公园黄河源园区藏野驴和藏原羚种群数量及其栖息地的影响. 兽类学报, 2022, 42(1): 34-48. |
| [17] |
Zhao H D, Liu S L, Dong S K, Su X K, Liu Q, Deng L. Characterizing the importance of habitat patches in maintaining landscape connectivity for Tibetan antelope in the Altun Mountain National Nature Reserve, China. Ecological Research, 2014, 29(6): 1065-1075. DOI:10.1007/s11284-014-1193-7 |
| [18] |
Wu X Y, Dong S K, Liu S L, Su X K, Han Y H, Shi J B, Zhang Y, Zhao Z Z, Sha W, Zhang X, Gao F, Xu D H. Predicting the shift of threatened ungulates' habitats with climate change in Altun Mountain National Nature Reserve of the Northwestern Qinghai-Tibetan Plateau. Climatic Change, 2017, 142(3): 331-344. |
| [19] |
Zhang J J, Jiang F, Li G Y, Qin W, Wu T, Xu F S, Hou Y, Song P, Cai Z, Zhang T Z. The four antelope species on the Qinghai-Tibet Plateau face habitat loss and redistribution to higher latitudes under climate change. Ecological Indicators, 2021, 123: 107337. DOI:10.1016/j.ecolind.2021.107337 |
| [20] |
Chrétien L P, Théau J, Ménard P. Visible and thermal infrared remote sensing for the detection of white-tailed Deer using an unmanned aerial system. Wildlife Society Bulletin, 2016, 40(1): 181-191. DOI:10.1002/wsb.629 |
| [21] |
Carr N L, Rodgers A R, Kingston S R, Hettinga P N, Thompson L M, Renton J L, Wilson P J. Comparative woodland caribou population surveys in Slate Islands Provincial Park, Ontario. Rangifer, 2012, 205-217. DOI:10.7557/2.32.2.2270 |
| [22] |
Sells S N, Costello C M, Lukacs P M, Roberts L L, Vinks M A. Grizzly bear habitat selection across the Northern Continental Divide Ecosystem. Biological Conservation, 2022, 276: 109813. DOI:10.1016/j.biocon.2022.109813 |
| [23] |
Valeix M, Fritz H, Sabatier R, Murindagomo F, Cumming D, Duncan P. Elephant-induced structural changes in the vegetation and habitat selection by large herbivores in an African savanna. Biological Conservation, 2011, 144(2): 902-912. DOI:10.1016/j.biocon.2010.10.029 |
| [24] |
Lechowicz M J. The sampling characteristics of electivity indices. Oecologia, 1982, 52(1): 22-30. DOI:10.1007/BF00349007 |
| [25] |
Jiao W J, Liu X Y, He S Y. Establishing an ecological monitoring system for National Parks in China: a theoretical framework. Ecological Indicators, 2022, 143: 109414. DOI:10.1016/j.ecolind.2022.109414 |
| [26] |
Jiang F, Li G Y, Qin W, Zhang J J, Lin G H, Cai Z Y, Gao H M, Zhang T Z. Setting priority conservation areas of wild Tibetan gazelle (Procapra picticaudata) in China's first National Park. Global Ecology and Conservation, 2019, 20: e00725. DOI:10.1016/j.gecco.2019.e00725 |
| [27] |
Li S, Yu D, Huang T, Hao R F. Identifying priority conservation areas based on comprehensive consideration of biodiversity and ecosystem services in the Three-River Headwaters Region, China. Journal of Cleaner Production, 2022, 359: 132082. DOI:10.1016/j.jclepro.2022.132082 |
| [28] |
Fritsch C J, Hanekom C, Downs C T. Hippopotamus population trends in Ndumo Game Reserve, South Africa, from 1951 to 2021. Global Ecology and Conservation, 2021, 32: e01910. DOI:10.1016/j.gecco.2021.e01910 |
| [29] |
Schwegmann S, Hendel A L, Frey J, Bhardwaj M, Storch I. Forage, forest structure or landscape: what drives roe Deer habitat use in a fragmented multiple-use forest ecosystem?. Forest Ecology and Management, 2023, 532: 120830. DOI:10.1016/j.foreco.2023.120830 |
| [30] |
Di Bitetti M S, Iezzi M E, Cruz P, Varela D, De Angelo C. Effects of cattle on habitat use and diel activity of large native herbivores in a South American rangeland. Journal for Nature Conservation, 2020, 58: 125900. |
| [31] |
Cai Z, Song P, Wang J, Jiang F, Liang C B, Zhang J J, Gao H M, Zhang T Z. Grazing pressure index considering both wildlife and livestock in Three-River Headwaters, Qinghai-Tibetan Plateau. Ecological Indicators, 2022, 143: 109338. DOI:10.1016/j.ecolind.2022.109338 |
| [32] |
Yan N N, Zhu W W, Wu B F, Tuvdendorj B, Chang S, Mishigdorj O, Zhang X W. Assessment of the grassland carrying capacity for winter-spring period in Mongolia. Ecological Indicators, 2023, 146: 109868. DOI:10.1016/j.ecolind.2023.109868 |