 2023, Vol. 43
2023, Vol. 43文章信息
- 马乐, 闫勇智, 于佳伟, 弓晓倩, 李奉时, 张庆
- MA Le, YAN Yongzhi, YU Jiawei, GONG Xiaoqian, LI Fengshi, ZHANG Qing
- 基于功能性状的毛乌素沙地不同演替阶段适生植物筛选
- Screening of suitable plants based on functional traits at different succession stages in Mu Us Sandy Land
- 生态学报. 2023, 43(20): 8598-8607
- Acta Ecologica Sinica. 2023, 43(20): 8598-8607
- http://dx.doi.org/10.20103/j.stxb.202301290143
-
文章历史
- 收稿日期: 2023-01-29
- 采用日期: 2023-07-21





2. 草原生态安全省部共建协同创新中心, 呼和浩特 010021
2. Collaborative Innovation Center for Grassland Ecological Safety, Hohhot 010021, China
沙漠化指干旱半干旱地区由于自然因素和人类活动引起生态系统破坏、地表呈现类似沙漠景观的土地退化现象[1]。我国是受沙漠化影响最严重的国家之一, 沙化面积约264万km2[2]。沙化土地面积的迅速扩展, 已严重威胁到人们的生存与居住安全, 恢复和重建沙漠化区域的植被无疑是生态恢复的关键环节[3]。
植被恢复过程中, 选择合适物种是关键问题之一[4]。国内外学者多采用试验法、价值法、综合效益评估法等对物种进行评价与筛选。试验法通过种植和培育植物幼苗或种子, 探讨其对环境的响应以提高抗逆性。Andivia等[5]培育抗胁迫敏感种子, 提高湿润生境下被子植物的存活率。王铁娟等[6]在沙地种植乌丹蒿等固沙植物, 揭示其幼苗对沙埋和水分的响应。价值法分析植物潜在恢复价值以提供植被恢复的理论依据。如研究物种组成差异及生态价值, 为近自然林恢复提供了方案[7]。也有学者对盐漠带植物的物种多样性及生态位宽度进行分析以筛选耐旱耐盐植物[8]。综合效益评价法在试验法、价值法的基础上获得指标, 对所选物种进行整体判别, 如通过层次分析法构建植物综合评价体系筛选绿地植物[9]。然而, 以上方法多基于生态恢复专家的基础知识和实践经验, 易造成一定偏差或所需时间过长, 需要依赖当地历史背景信息[10]。近年来, 恢复生态学家发现, 生态系统的成功恢复需要基于生态系统功能, 物种特征反映了对生态系统资源的使用与权衡, 基于植物特征的方法可有效地确定适合恢复目标的物种[11]。
植物在长期适应环境的过程中, 内部功能之间协同进化, 形成能够响应外部环境变化, 并对生态系统功能产生一定影响的植物性状, 称为植物功能性状[12]。植物性状特征可以解释植物形态与生理面对环境胁迫的变化, 如长期缺水条件下植物通过降低叶面积、增加比叶面积等叶性状减少蒸腾、降低光合速率, 促进更有效的散热[13]。许多研究证明, 基于功能性状的方法已成为恢复生态学领域的突破点。如利用功能性状构建物种筛选指标体系, 建立优良物种名录[14]。Modolo等[15]测量了生长在2种光环境下的6种热带树种的树冠性状及叶性状, 通过性状对光环境的响应以识别强光环境下生长性能最好的树种。Bochet等[16]分析了10个植物性状对半干旱路坡侵蚀-生产力梯度对物种定植的贡献, 并建立296个物种包含叶片、种子和根的性状数据库。选择合适的功能性状指标可在一定程度上反映植物适应环境的能力, 更有效地选择适生物种。
植物是发展到各演替阶段最敏感和直观的标志, 群落演替动态反映植被对环境变化的响应[17]。沙地植被恢复过程中, 植物生活型功能群组成会发生变化。恢复早期, 生长在流动沙丘上的植被多以一年生植物为主, 演替中后期多年生植物占主导地位[18]。菊科、小型禾草随演替进行占据优势地位, 而多年生禾草和豆科固氮植物在整个演替中所占比例较大[19]。群落演替受环境驱动, 环境因子作用不同。如我国西南喀斯特地区的主要环境驱动力是高景观异质性、地形、土壤理化性质[20]。群落演替为物种筛选提供理论支撑, 跨演替阶段进行植被恢复不易成功, 应充分考虑群落的演替阶段及当前生态环境。
毛乌素沙地是我国四大沙地之一, 是具有特殊地理位置和环境条件的生态过渡带, 也是生态环境脆弱区, 其沙化问题严重威胁北方地区生态安全, 生态恢复刻不容缓[21]。飞播即将适宜当地环境的草种或树种由飞机撒播, 对宜播地进行封闭管护直至植被恢复的措施, 以达到防沙治沙的目的[22]。毛乌素沙地是我国最早开始飞播治沙并取得显著效果的地区之一, 植物多样性随恢复年限的增加形成不同的演替序列, 为研究群落恢复的演替特征提供了理想条件。本研究以毛乌素沙地32个飞播样地的植物群落为调查对象, 分析不同演替阶段环境因子的影响, 确定各演替阶段目标物种, 建立了包含161种沙地植物, 10个性状的物种库。探究不同演替阶段下毛乌素沙地的适生植物, 旨在揭示基于功能性状下筛选物种对于生态恢复的可行性及意义, 以期为沙漠化区域的植被保护和生态恢复提供理论依据。
1 研究地区与研究方法 1.1 研究区概况毛乌素沙地位于北纬37°27.5′—39°22.5′, 东经107°20′—111°30′之间, 包括陕西省榆林市的北部, 内蒙古自治区鄂尔多斯市南部及宁夏回族自治区盐池县东北部, 海拔1000—1600m, 总面积达4.00×104 km2。沙地地处我国东部季风区边缘地带, 属温带干旱、半干旱大陆性季风气候, 年平均气温6—8℃, 降水集中于7—9月, 年降水量300—400mm, 年蒸发量约为降水量的4—10倍, 在2200—2800mm之间。地带性土壤为风沙土、栗钙土, 自然植被由南向北从森林草原向荒漠草原过渡, 主要植物种类有碱茅(Puccinellia distans)、白草(Pennisetum centrasiaticum)、软毛虫实(Corispermum puberulum)等, 主要人造乔木树种有旱柳(Salix matsudana)、榆树(Ulmus pumila L.)等, 其次还有较多的人造灌木及草类, 如沙打旺(Astragalus adsurgens)、花棒(Hedysarum scoparium)等[23]。
1.2 植物群落调查研究区自20世纪80年代开始进行飞播措施, 本研究于2017年8月进行野外取样, 选取1983—2015年30个飞播样地(无1986、1988、2003年样地)以及一个未飞播流动沙丘样地和一个多年未干扰原始样地, 共32个样地(图 1)。样地平均飞播年限17年, 最长飞播年限34年, 最短飞播年限2年, 每个样地主要飞播物种有羊柴(Hedysarum laeve)、花棒(Hedysarum scoparium)、沙打旺(Astragalus adsurgens)等, 且混播比例相近。所有飞播样地恢复前的初始状态均为流动沙丘。样地均设置在沙丘间平坦处, 地形地貌保持一致。

|
| 图 1 毛乌素沙地飞播样地分布图 Fig. 1 Distribution of aerial seeding plot in Mu Us Sandy Land |
每个样地设置3个1m×1m的样方, 逐一按照植物物种名称记录其株丛数, 分种齐地面剪下植物地上部分, 置于65℃烘箱中烘干至恒重。每个样方中心位置使用土钻钻取0—60cm土壤, 去除可见的植物根、石块、枯枝落叶和碎屑, 混合均匀的土壤分为两份, 一份常温保存用于土壤养分含量测定, 另一份低温保存用于土壤微生物测定。
本研究测定6个土壤养分指标, 分别为土壤全氮、土壤硝态氮、土壤铵态氮、土壤总有机碳、土壤有效磷、土壤全磷。土壤全氮采用凯氏定氮法测定, 土壤硝态氮、铵态氮使用AA3型连续流动分析仪测定, 土壤总有机碳采用重铬酸钾容量法测定, 土壤有效磷采用碳酸氢钠浸提-钼锑抗比色法测定, 土壤全磷采用碱熔-钼锑抗比色法测定。
根据每个样地的经纬度, 在全球气候数据库(www.worldclim.org) 获取与植物生长分布密切相关的四个气候因子作为气候指标, 分别是年平均降水(MAP)、年平均气温(MAT)、最冷月最低温度(LTCM)、最干燥月份降水(PDM)。
1.3 目标物种选择及潜在种库建立选择飞播后不同演替阶段中相对重要性高的物种作为目标物种, 采用物种的相对干重表示其在群落中相对重要性。物种库中物种来自于两部分, 一部分是文献搜集, 包括毛乌素沙地及其他沙地物种, 另一部分是在毛乌素沙地常年开展调查所记录的物种[24]。初始共统计物种918种, 进一步选择具有抗旱、固沙喜沙、根系发达、易繁殖等特点, 且生境是荒漠和沙地的植物, 最终共161种物种构成潜在种库。
根据毛乌素沙地环境特点, 选取与资源获取能力、抗旱能力相关的10个功能性状(表 1)。所需性状数据在全球植物性状数据库(TRY plant trait database, www.try-db.org)中进行搜集。
| 性状类别 Classification of functional traits |
性状 Functional traits |
单位 Unit |
生态相关性 Ecological correlation |
| 全株性状 | 生活型 | 群落形态结构、气候适应 | |
| Whole plant characters | 生长史 | 群落形态结构、气候适应 | |
| 系统发育类型 | 群落形态结构、气候适应 | ||
| 植物高度 | cm | 群落形态结构、气候适应 | |
| 光合途径 | 气候适应、资源利用 | ||
| 叶片性状 | 比叶面积 | cm2/g | 养分获取、资源保留 |
| Leaf characters | 叶干物质含量 | g/g | 养分获取、资源保留 |
| 叶片碳含量 | mg/g | 养分吸收利用、防御保护 | |
| 叶片磷含量 | mg/g | 养分吸收利用、防御保护 | |
| 叶片氮含量 | mg/g | 养分吸收利用、防御保护 |
本研究计算了不同样地间物种组成的Sørensen相异性指数, 以量化物种组成差异[25]。进而构建32个飞播样地的物种组成差异矩阵, 采用主坐标分析(PCoA)和双向指示种分类方法(TWINSPAN)对飞播样地的演替阶段进行划分。
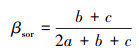
|
式中, βsor为两个样地之间的Sørensen相异性指数, a为两个样地间前一个样地的特有物种数, b为两个样地间的共有物种数, c为两个样地间后一个样地的特有物种数。
使用冗余分析(RDA)对影响因素和样地之间的关系进行分析, 确定影响飞播后群落演替的关键驱动因素。最后进行适宜物种筛选, 依据已确定的目标物种和种库性状数据, 计算161个物种和6个目标物种之间的高尔距离, 并进行聚类分析, 以衡量物种间相似性[26]。高尔距离计算公式如下[27]:
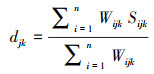
|
式中, n为功能性状数量, i为功能性状, j、k为物种, Wijk是功能性状i在物种j与k之间功能性状矩阵的权重, Sijk为物种j与k之间对于性状i的相似度。
TWINSPAN分析使用Wintwins 2.3软件完成, 其他分析均通过R 4.3.1完成, Sørensen相异性指数计算采用“betapart”程序包“beta.pa”函数, 主坐标分析采用“vegan”程序包“cmdscale”函数, 冗余分析采用“vegan”程序包“capscale”函数, Gower距离计算采用“FD”程序包“gowdis”函数。使用“ggplot2”、“pheatmap”包进行制图。
2 结果与分析 2.1 毛乌素沙地植物群落演替阶段划分飞播恢复下的毛乌素沙地植被排序结果见图 2, PCoA前两个排序轴(PCoA1, PCoA2)分别解释了植物群落物种组成变化的36.42%和23.72%, 两者对群落组成差异解释的贡献率达60.14%, 能较好反映群落组成的变化。PCoA排序图中将所有样地分类A、B、C具有明显界限的三类。TWINSPAN将所有样地也划分为Ⅰ、Ⅱ、Ⅲ三类, 三类分别对应PCoA排序图中A、B、C三个区域(图 3)。有30个样地在PCoA排序和TWINSPAN分类中是吻合的, 因此, 本研究根据TWINSPAN分类和PCoA排序结果, 确定飞播恢复下毛乌素沙地植物群落演替序列, 即演替第一阶段(A):固沙先锋物种群落, 如蒙古虫实(Corispermum mongolicum)群落、沙蓬(Agriophyllum squarrosum)群落。演替第二阶段(B):沙生植物为主的杂类草群落, 如蒙古虫实群落、白草群落、车前(Plantago asiatica)群落等。演替第三阶段(C):中生植物为主的杂类草群落, 如阿尔泰狗娃花(Heteropappus altaicus)群落、华北白前(Cynanchum hancockianum)群落等。

|
| 图 2 毛乌素沙地植物群落的主坐标分析 Fig. 2 Principal co-ordinates analysis of plant communities in Mu Us Sandy Land A—C表示样地在图中聚集的不同区域 |

|
| 图 3 双向指示种分析分类树状图 Fig. 3 Two-way indicator species analysis classification tree 方框内数字表示样地号, Ⅰ—Ⅲ表示演替阶段; Ⅰ:演替第一阶段, 固沙先锋物种群落; Ⅱ:演替第二阶段, 沙生植物为主的杂类草群落; Ⅲ:演替第三阶段, 中生植物为主的杂类草群落 |
如RDA分析结果所示(图 4), 影响植物群落演替最主要的环境因素是土壤全氮、土壤总有机碳、土壤硝态氮。

|
| 图 4 毛乌素沙地不同演替阶段群落与环境变量冗余分析 Fig. 4 Redundancy analysis of community and environmental variables at different succession stages in Mu Us Sandy Land * 表示显著相关(P < 0.05), * * 表示极显著相关(P < 0.01) |
三个演替阶段的植物物种组成及各物种相对重要性存在明显差异(图 5)。演替第一阶段沙蓬的相对重要性为75.25%、蒙古虫实的相对重要性为24.75%, 选取演替第一阶段的优势物种, 沙蓬、蒙古虫实为目标物种。演替第二阶段蒙古虫实相对重要性73.46%、白草相对重要性7.08%、车前相对重要性6.68%, 选取蒙古虫实、白草、车前为演替第二阶段的目标物种; 演替第三阶段蒙古虫实相对重要性29.32%、猪毛蒿(Artemisia scoparia)相对重要性12.41%、地锦草相对重要性5.63%、华北白前相对重要性5.43%、阿尔泰狗娃花相对重要性4.81%。这些物种成为演替第三阶段的优势物种, 蒙古虫实、猪毛蒿、地锦草的相对重要性较高, 但为一、二年生植物, 竞争力较弱不利于生态恢复, 因此选取竞争力强的多年生植物华北白前和阿尔泰狗娃花为演替第三阶段目标物种。

|
| 图 5 不同演替阶段物种组成及相对重要性 Fig. 5 Species composition and relative importance at different succession stages |
根据聚类分析结果选取与各目标物种距离最近的前5种植物, 共30种。在这30种植物中, 由于狼毒为毒草, 不利于沙地修复, 因此去除有毒植物狼毒, 最终得到29种优势物种(表 2)。
| 优势物种 Dominant species |
目标物种 Target species |
与目标物种Gower距离 Gower distance from target species |
演替阶段 Stage of succession |
| 狗尾草 Setaria viridis | 沙蓬 | 0.17 | 阶段1 |
| 羊茅 Festuca ovina | 0.14 | ||
| 短花针茅 Stipa breviflora | 0.12 | ||
| 锋芒草 Tragus racemosus | 0.11 | ||
| 画眉草 Eragrostis pilosa | 0.11 | ||
| 拂子茅 Calamagrostis epigeios | 白草 | 0.07 | 阶段2 |
| 无芒隐子草 Cleistogenes songorica | 0.06 | ||
| 假苇拂子茅 Calamagrostis pseudophragmites | 0.06 | ||
| 冰草 Agropyron cristatum | 0.06 | ||
| 沙鞭 Psammochloa villosa | 0.02 | ||
| 瓦松 Orostachys fimbriatus | 车前 | 0.23 | 阶段2 |
| 狼毒 Stellera chamaejasme | 0.21 | ||
| 盐生车前 Plantago maritima | 0.16 | ||
| 蚓果芥 Torularia humilis | 0.16 | ||
| 条叶车前 Plantago minuta | 0.12 | ||
| 细叶益母草 Leonurus sibiricus | 华北白前 | 0.19 | 阶段3 |
| 蓝堇草 Leptopyrum fumarioides | 0.19 | ||
| 蒙古芯芭 Cymbaria mongolica | 0.19 | ||
| 香青兰 Dracocephalum moldavica | 0.19 | ||
| 地稍瓜 Cynanchum thesioides | 0.11 | ||
| 冷蒿 Artemisia frigida | 阿尔泰狗娃花 | 0.02 | 阶段3 |
| 乳苣 Mulgedium tataricum | 0.02 | ||
| 草地风毛菊 Saussurea amara | 0.01 | ||
| 猪毛蒿 Artemisia scoparia | 0.01 | ||
| 火绒草 Leontopodium leontopodioides | 0.01 | ||
| 雾冰藜 Bassia dasyphylla | 蒙古虫实 | 0.10 | 阶段1、阶段2 |
| 刺藜 Chenopodium aristatum | 0.05 | ||
| 猪毛菜 Salsola collina | 0.05 | ||
| 绳虫实 Corispermum declinatum | 0.03 | ||
| 蒙古猪毛菜 Salsola ikonnikovii | 0.02 |
物种更替、环境演变、物种对环境的适应性等方面, 对毛乌素沙地演替起到重要作用[28]。本研究将飞播造林后的毛乌素沙地植物群落分为三个演替阶段:固沙先锋物种群落阶段、沙生植物为主的杂类草群落阶段、中生植物为主的杂类草群落阶段。刘美珍等[29]将退化沙地恢复过程同样分为三个阶段, 物种的丰富度、生活型等先增加而后稳定。证明了各演替阶段植物群落特征存在差异。李禾等[30]根据飞播年限将毛乌素沙地植物群落划分为四个阶段:沙打旺、白沙蒿、沙蓬群落, 羊柴、芦苇+沙地先锋物种群落, 羊柴+虫实、雾冰藜杂草群落, 油蒿、羊柴+狗尾草杂草群落。这与本文演替划分结果基本一致, 群落物种组成变化从固沙先锋物种到具有耐旱性质的杂类草, 与其结果不同的是本研究主要关注了对草本植物的演替。本研究中, 各演替阶段与自然演替序列的植物群落组成不同, 其原因是飞播恢复下的植物群落处于较早期演替阶段, 尚未恢复到顶级演替群落。
环境因子对群落演替的影响存在差异, 如气候、土壤、地形等[31—32]。已有研究证明, 土壤全氮含量是毛乌素沙地飞播后植被演替的重要驱动力, 这与本研究得出的土壤是演替的主导因子, 且对群落影响土壤全氮>土壤总有机碳>土壤硝态氮的研究结果一致。其原因是飞播年份的增加使土壤环境得到改善, 根系发育、微生物活动等使土壤营养元素供应程度提高, 导致土壤中氮素源增加[33]。也有研究认为, 碳元素通过光合作用为植物生长发育提供能量, 随着演替的进行, 土壤总有机碳增加, 土壤腐殖质化加速, 导致土壤全氮含量增加[34]。因此, 从侧面证明了土壤有机碳对演替也具有重要作用。氮素可被植物利用的形态主要有硝态氮和铵态氮, 本研究中硝态氮对群落的影响更大, 这与李荣等研究结果一致。李荣等[35]发现干旱-半干旱地区的土壤中大部分为硝态氮, 铵态氮多被微生物和植物固定、吸收或转化为硝态氮。由于毛乌素沙地土壤整体呈碱性, pH升高使硝化作用加强, 所以碱性条件下土壤中硝态氮多于铵态氮[36]。土壤全磷及有效磷对演替影响不显著的原因可能是沙地土壤以风沙土为主, 对磷的吸附能力较低且主要被固沙植物所吸收[37]。而气候因子长期影响土壤物理结构与化学组成, 推动土壤变化控制植物发育, 与土壤耦合作用共同驱动演替的进行。探讨沙地环境因子之间的关系, 有助于了解物种的适应性, 更好地筛选适生物种。
本研究确定了具有较高重要性的6种植物作为目标物种。演替第一阶段的目标物种具有耐旱固沙的特性, 如沙蓬、蒙古虫实, 性状表现为植株矮小、表皮毛发达、低比叶面积等[38]。演替第二阶段的目标物种白草、车前是根茎发达的禾本科植物, 有较强的吸水吸肥能力, 这类快速生长型植物在此阶段对资源更具竞争力[39]。演替第三阶段的目标物种阿尔泰狗娃花和华北白前是多年生植物, 具有较高植株高度, 种子小而轻等性状使其自繁衍生能力较强, 在此阶段可获得更多养分[40]。目标物种的特征进一步证实了功能性状是了解物种适应退化生态系统的有效方法, 本研究筛选出与目标物种具有相似适应策略的适生植物。演替第一、二阶段以藜科、禾本科为主。藜科植物多生活在荒漠地区, 其根系深且发育迅速, 叶片缩小甚至消失, 整体呈现旱生的适应现象[41]。禾本科植物的根系发达、须根多易分蘖, 且对土质要求不大, 可起到改善作用。所以藜科、禾本科植物适合演替第一阶段及向下一阶段过渡时作为沙地适生物种。演替第三阶段菊科植物居多, 菊科植物在此阶段以繁殖力和生产力占优势[42]。因此, 认为利用功能性状可在退化生态系统中找到更多具有适应恶劣环境能力的物种。
本研究所筛选的适生物种可为人工辅助植被恢复时提供选择方案, 如演替第一阶段可选雾冰藜、猪毛菜、蒙古虫实等耐旱固沙植物。演替第二阶段可选拂子茅、无芒隐子草等根糵发达的禾本科植物。草地风毛菊、猪毛蒿等具有较高竞争能力的菊科植物可用于演替第三阶段。所选适生植物不仅有助于不同演替阶段的植被恢复, 还可提供一定的经济、社会效益。如华北白前全草可作绿肥和杀虫药, 种子可榨工业用油; 雾冰藜可与其他一年生藜科、禾本科野草混贮, 起到抗灾保畜的作用; 短花针茅、冰草等是优良饲草, 瓦松、蚓果芥、细叶益母草等具有良好的药用价值[32, 43]。所筛选出的适生植物在进行恢复时要根据植物特性与所恢复的环境进行选择, 充分挖掘植物的生产潜力。
| [1] |
王涛, 朱震达. 我国沙漠化研究的若干问题——1.沙漠化的概念及其内涵. 中国沙漠, 2003, 23(3): 209-214. |
| [2] |
王涛, 赵哈林, 肖洪浪. 中国沙漠化研究的进展. 中国沙漠, 1999, 19(4): 299-311. |
| [3] |
牛兰兰, 张天勇, 丁国栋. 毛乌素沙地生态修复现状、问题与对策. 水土保持研究, 2006, 13(6): 239-242, 246. |
| [4] |
程晔, 庄毓琼, 黄林冲. 高速公路边坡生态防护物种选择的模糊多属性决策方法. 水土保持通报, 2014, 34(3): 201-206. |
| [5] |
Andivia E, Villar-Salvador P, Oliet J A. Climate and species stress resistance modulate the higher survival of large seedlings in forest restorations worldwide. Ecological Applications, 2021, 31(6): e02394. DOI:10.1002/eap.2394 |
| [6] |
王铁娟, 杨持, 吕桂芬, 马春明, 陈海英. 中国北部六种沙蒿的地理替代规律及其主导生态因子. 生态学报, 2005, 25(5): 1012-1018. |
| [7] |
Ho K V, Kröel-Dulay G, Tölgyesi C, Bátori Z, Tanács E, Kertész M, Török P, Erdös L. Non-native tree plantations are weak substitutes for near-natural forests regarding plant diversity and ecological value. Forest Ecology and Management, 2023, 531: 120789. DOI:10.1016/j.foreco.2023.120789 |
| [8] |
潘旭东, 吴英翔, 张凤华. 玛纳斯河流域绿洲盐漠带适生植物筛选及物种多样性研究. 农学学报, 2011, 1(5): 34-37. |
| [9] |
刘本本, 孙清琳, 柳鑫, 易三桂, 郭姣姣, 周继华, 来利明, 郑元润. 基于群决策和层次分析法的雄安新区公园绿地植物综合评价体系构建与物种筛选. 应用与环境生物学报, 2022, 28(3): 770-778. |
| [10] |
Ostertag R, Warman L, Cordell S, Vitousek P M. Using plant functional traits to restore Hawaiian rainforest. Journal of Applied Ecology, 2015, 52(4): 805-809. DOI:10.1111/1365-2664.12413 |
| [11] |
Stokes A, Douglas G B, Fourcaud T. Ecological mitigation of hillslope instability: ten key issues facing researchers and practitioners. Plant and Soil, 2014, 377(1-2): 1-23. DOI:10.1007/s11104-014-2044-6 |
| [12] |
Diaz S, Cabido M. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology & Evolution, 2001, 16(11): 646-655. |
| [13] |
Yan Y Z, Liu Q F, Zhang Q, Ding Y, Li Y H. Adaptation of dominant species to drought in the inner Mongolia grassland-species level and functional type level analysis. Frontiers in Plant Science, 2019, 10: 231. DOI:10.3389/fpls.2019.00231 |
| [14] |
徐艺露, 杨晓东, 许月, 谢一鸣, 王良衍, 阎恩荣. 基于功能性状的亚热带常绿植物抗雨雪冰冻能力评价. 应用生态学报, 2012, 23(12): 3288-3294. |
| [15] |
Modolo G S, Santos V A H F, Ferreira M J. Testing for functional significance of traits: effect of the light environment in tropical tree saplings. Ecology and Evolution, 2021, 11(11): 6480-6492. DOI:10.1002/ece3.7499 |
| [16] |
Bochet E, García-Fayos P. Identifying plant traits: a key aspect for species selection in restoration of eroded roadsides in semiarid environments. Ecological Engineering, 2015, 83: 444-451. DOI:10.1016/j.ecoleng.2015.06.019 |
| [17] |
赵丽娅, 赵哈林. 我国沙漠化过程中的植被演替研究概述. 中国沙漠, 2000, 20(S1): 8-15. |
| [18] |
徐彩琳, 李自珍. 干旱荒漠区人工植物群落演替模式及其生态学机制研究. 应用生态学报, 2003, 14(9): 1451-1456. |
| [19] |
詹瑾, 韩丹, 杨红玲, 李玉霖. 科尔沁沙地植被恢复过程中群落组成及多样性演变特征. 中国沙漠, 2022, 42(2): 194-206. |
| [20] |
文丽, 宋同清, 杜虎, 王克林, 彭晚霞, 曾馥平, 曾昭霞, 何铁光. 中国西南喀斯特植物群落演替特征及驱动机制. 生态学报, 2015, 35(17): 5822-5833. |
| [21] |
张海欧. 毛乌素沙地综合整治现状分析及新思路. 农学学报, 2018, 8(5): 55-59. |
| [22] |
刘玉平. 毛乌素沙地飞播植被演替研究. 中国草地, 1996, 18(4): 24-27. |
| [23] |
周淑琴, 荆耀栋, 张青峰, 吴发启. 毛乌素沙地南缘植被景观格局演变与空间分布特征. 生态学报, 2013, 33(12): 3774-3782. |
| [24] |
刘翠英. 毛乌素沙地野生观赏植物资源及开发利用研究. 西北林学院学报, 2006, 21(4): 158-161. |
| [25] |
陈圣宾, 欧阳志云, 徐卫华, 肖燚. Beta多样性研究进展. 生物多样性, 2010, 18(4): 323-335. |
| [26] |
Petchey O L, Gaston K J. Dendrograms and measuring functional diversity. Oikos, 2007, 116(8): 1422-1426. |
| [27] |
Gower J C. A general coefficient of similarity and some of its properties. Biometrics, 1971, 27(4): 857. |
| [28] |
慕宗杰, 刘果厚, 桂荣, 苏海林, 李红颖, 刘利红, 曹瑞, 姜莹, 王亚东. 浑善达克沙地飞播区不同恢复阶段植物群落结构动态变化. 华中农业大学学报, 2020, 39(4): 46-56. |
| [29] |
刘美珍, 蒋高明, 于顺利, 李永庚, 高雷明, 牛书丽, 姜闯道, 彭羽. 浑善达克退化沙地恢复演替18年中植物群落动态变化. 生态学报, 2004, 24(8): 1734-1740. |
| [30] |
李禾, 吴波, 杨文斌, 张武文, 崔力强, 刘静. 毛乌素沙地飞播区植被动态变化研究. 干旱区资源与环境, 2010, 24(3): 190-194. |
| [31] |
王飞, 郭树江, 纪永福, 张莹花, 韩福贵, 张裕年, 张卫星, 宋达成. 不同演替阶段白刺灌丛沙堆土壤因子与叶功能性状关系研究. 干旱区地理, 2022, 45(1): 176-184. |
| [32] |
席璐璐, 缑倩倩, 王国华, 宋冰. 荒漠绿洲过渡带一年生草本植物对干旱胁迫的响应. 生态学报, 2021, 41(13): 5425-5434. |
| [33] |
赵朋波, 邱开阳, 谢应忠, 赵香君, 陈林, 刘王锁, 李海泉, 孟文芬, 虎学琴, 何毅, 黄业芸, 李亚园. 毛乌素沙地南缘不同固沙灌木下土壤养分的空间异质性. 草地学报, 2021, 29(9): 2040-2048. |
| [34] |
陈晨, 王寅, 王健铭, 杨欢, 王雨辰, 徐超, 李景文, 褚建民. 科尔沁沙地植物群落物种多样性及其主要影响因素. 北京林业大学学报, 2020, 42(5): 106-114. |
| [35] |
李荣, 何兴东, 张宁, 邬畏. 沙丘固定过程中土壤铵态氮和硝态氮的时空变化. 土壤学报, 2010, 47(2): 295-302. |
| [36] |
Narteh L T, Sahrawat K L. Potentially mineralizable nitrogen in West African lowland rice soils. Geoderma, 1997, 76(1/2): 145-154. |
| [37] |
苏永中, 赵哈林, 张铜会, 赵学勇. 不同退化沙地土壤碳的矿化潜力. 生态学报, 2004, 24(2): 372-378. |
| [38] |
李善家, 苏培玺, 张海娜, 周紫鹃, 解婷婷. 荒漠植物叶片水分和功能性状特征及其相互关系. 植物生理学报, 2013, 49(2): 153-160. |
| [39] |
张磊, 吕光辉, 蒋腊梅, 李周康, 张仰. 干旱区荒漠植物初级生产力及碳储量与功能性状之间的关系. 新疆大学学报: 自然科学版, 2020(1): 63-74. |
| [40] |
姜晓燕, 高圣杰, 蒋燕, 田赟, 贾昕, 查天山. 毛乌素沙地植被不同恢复阶段植物群落物种多样性、功能多样性和系统发育多样性. 生物多样性, 2022, 30(5): 18-28. |
| [41] |
邓彦斌, 姜彦成, 刘健. 新疆10种藜科植物叶片和同化枝的旱生和盐生结构的研究. 植物生态学报, 1998, 22(2): 164-170. |
| [42] |
陈拓, 冯虎元, 徐世建, 强维亚, 安黎哲. 荒漠植物叶片碳同位素组成及其水分利用效率. 中国沙漠, 2002, 22(3): 288-291. |
| [43] |
刘翠英. 毛乌素沙地植物资源调查及开发利用研究. 干旱地区农业研究, 2006, 24(2): 183-188. |