 2023, Vol. 43
2023, Vol. 43文章信息
- 苟小林, 周青平, 涂卫国, 陈有军, 魏小星, 张伟
- GOU Xiaolin, ZHOU Qingping, TU Weiguo, CHEN Youjun, WEI Xiaoxing, ZHANG Wei
- 高寒半湿润沙地植物与土壤微生物多样性对植被恢复的响应
- The responses of biodiversity and community composition in vegetation and soil microbes to restoration in alpine cold and sub-humid desertified land
- 生态学报. 2023, 43(20): 8442-8453
- Acta Ecologica Sinica. 2023, 43(20): 8442-8453
- http://dx.doi.org/10.20103/j.stxb.202110303047
-
文章历史
- 收稿日期: 2021-10-30
- 采用日期: 2023-07-21





2. 西南民族大学青藏高原研究院, 成都 610041;
3. 青海大学畜牧兽医科学院, 青海省青藏高原优良牧草种质资源利用重点实验室, 西宁 810016
2. Institute of Qinghai-Tibetan Plateau, Southwest Minzu University, Chengdu 610041, China;
3. Key Laboratory of Use of Forage Germplasm Resources on Tibetan Plateau of Qinghai Provence, Institute of Grazing & Veterinarian, Qinghai University, Xining 810016, China
青藏高原地处亚洲内陆, 是中国最大的高原, 其作为“亚洲水塔”和“高寒生物资源基因库”, 对长江和黄河流域水源涵养、水土保持、气候调节、生物保护等方面作用巨大[1]。但是受到全球气候变化、人为干扰等综合作用的影响下, 青藏高原生态退化严重, 尤其是沙漠化已经成为青藏高原生态功能稳定的重大威胁[2—4]。目前青藏高原沙化面积已经超过3.6×105 km2, 约占青藏高原总面积的14%, 长江、黄河、雅鲁藏布江源区分布着大片的活动沙丘, 沙化所引发的沙尘甚至能够远达我国北方沿海城市, 青藏高原沙漠化防治形势严峻[5], 亟需进行沙漠化恢复, 以促进生态功能恢复与生物多样性保护。
针对现阶段青藏高原沙漠化防治问题, 已经提出的措施包括围栏休牧、草灌方格、植被恢复等措施, 如在共和盆地[6—7]、青海湖流域[8—9]、川西北[10—11]等地区开展的沙化治理成效显著, 且植被恢复模式应用范围更加广泛。目前沙化植被恢复对土壤微生物和植物多样性的研究相对丰富, 最新研究结果表明, 虽然不同的植被恢复模式会对土壤微生物群落结构产生影响[12], 但是植被恢复能够促进沙化地土壤中细菌和真菌群落发生变化, 微生物功能群落能够极大的促进土壤养分转化过程[13]; 也有相关研究表明植被恢复使沙化地土壤结皮及土壤表层微生物群落演替[14], 引发土壤微生物群落功能发生转变[15]。利用植被恢复沙化土地, 促使地表植物多样性增加, 并认为植物多样性增加促进了生态系统功能恢复[16], 而植物群落的这种演替过程被认为是植被恢复过程中栽培物种与本土物种之间的种间竞争造成的[17]; 也有研究表明, 植被恢复沙化地过程促进了植物多样性增加, 群落趋于稳定, 并且一年生植物占据优势[18], 但是长期恢复后植物群落呈现退化现象, 植物群落多样性降低[19]。而针对青藏高原的植被恢复沙化土地研究则表明不同植被恢复模式促进了植物多样性增加, 只是不同恢复模式下的优势物种不一样[20]。但这些关于青藏高原植被恢复沙化地的研究中, 对于恢复过程中植物群落的响应过程、土壤微生物群落季节性变化的研究较少, 相关结论缺乏。
青藏高原东南缘属典型的高寒半湿润区, 该地区草地分布广泛, 河道密集, 对长江和黄河水源涵养能力巨大, 该区域泥炭储量巨大, 对区域碳平衡具有非常重要的调节作用, 该区地形多变, 从高山森林到高原湿地, 生物种质资源储量丰富[21—22], 是青藏高原极为重要的组成部分。该地区作为我国第五大牧区, 受大量放牧与全球气候变化影响, 区域内草地退化加剧, 沙化迅速扩张, 脆弱生境亟需生态保护和生态修复[4]的相关研究提供恢复建议。针对该地区沙漠化问题, 适生于高寒、贫瘠生境, 抗风沙的根茎型草本老芒麦(Elymus sibiricus L.)进行植被恢复已经被成功应用于该地区沙化治理工作, 同时该区开展了恢复模式效果比较[23]、恢复后生态功能评价[10]、恢复过程中养分变化过程[24]等研究, 但该区域人工种植草本恢复沙化草地, 植物种类相对单一且群落结构并不稳定, 那么植物群落的演替过程中物种组成是否剧烈变化?土壤微生物群落是否存在演替过程?这对沙化草地恢复后续管理非常关键, 但缺乏研究。因此, 本研究以青藏高原东南缘半湿润沙化地恢复为基础, 以老芒麦作为建群种进行植被恢复, 研究植物群落物种多样性是否响应植被恢复过程, 明确优势物种的变化情况, 并研究土壤微生物群落在关键季节与恢复过程中对植被恢复的响应过程, 为高寒脆弱沙化草地生态恢复提供基础研究。
1 材料和方法 1.1 研究区域概况研究区域位于青藏高原东南缘的四川省阿坝州红原县日干乔生态保护区(33.18°N, 102.62°E, 海拔3470 m), 为典型的高寒半湿润区域, 近五年的年均降水量超过800 mm, 该区域年均气温2.5℃左右, 生长季节较短, 雨热集中, 每年的5月至9月都是生长季节, 春、秋二季短暂, 冬季较为漫长, 雪被覆盖持续时间较长, 土壤在经过频繁的冻融条件后进入和结束冬季过程。作为我国主要的高寒牧区, 该地区过度放牧牦牛与人工开沟排水, 造成了大面积草原退化、土地沙化, 沙化土壤贫瘠, 土壤水分含量低。沙化地周边地表植物主要为适应高寒环境的中国沙棘(Hippophae rhamnoides)、西藏沙棘(Hippophae tibetana)、沙蒿(Artemisia desertorum)、蕨麻(Potentilla anserina)、高山嵩草(Carex pygmaea)、四川嵩草(Carex setschwanensis)等。
1.2 植被恢复用于植被恢复的种植物种为老芒麦(Elymus sibiricus L.), 该种为多年生疏丛型禾本科草本, 须根系, 根系繁茂, 茎直立, 分蘖能力强, 该物种广布于青藏高原大部分区域, 对高海拔寒旱环境具有非常强的抗性, 耐贫瘠, 适宜高寒沙地种植, 是进行沙化治理和生态修复的优良材料。
高寒半湿润沙地植被恢复采用老芒麦进行逐年种植, 逐步控制沙化蔓延。老芒麦于2014年至2017年每年5月上旬在沙化地条件相似的不同区域进行种植。为避免施肥干扰, 用于研究地种植材料未施用肥料。老芒麦采用撒种方式进行带播, 播种带宽1 m, 长超过10 m, 每条播种带之间间隔为1 m, 0.5 km×1 km为一个样方, 每年播种5个样方, 每个样方选用半固定沙丘地势较为平坦的沙地, 地势基本相同, 每个样方之间被5 m宽的柳树灌丛隔开。根据超过400万株/hm2的保苗率, 老芒麦的播种量为50 kg/hm2。均匀的撒种后覆土, 深度为5 cm左右, 稍压, 防止水分过度蒸发。在相邻的空白沙地选定5块对照样地, 样地地势和种植地势相似, 并且样方面积基本相同。所有样方处理后都进行人工封育, 防止人为破坏和家畜采食。
1.3 样地设置与样品采集本研究采用空间代替时间方法, 在2016年至2017年完成所有样品的采集工作, 分别选用种植后生长1年、2年和3年的老芒麦作为植被恢复年限, 以选择的空白沙地作为对照组(CK)。根据老芒麦生长周期和区域气候特征, 将每年的样品周期分为生长季节和非生长季节进行样品采集。非生长季节根据冬季气候和土壤环境特征, 分为非生长季节前期(EN, 2016年11月)和非生长季节后期(LN, 2017年4月)进行样品采集; 生长季节根据老芒麦生长周期, 分为生长季节前期(EG, 2017年6月), 生长季节中期(MG, 2017年8月)和生长季节后期(LG, 2017年9月)进行样品采集。在五个0.5 km×1 km的种植样地中, 沿对角线选择5块5 m×5 m的大样方, 用于5个典型季节样品采集, 在大样方中随机选择一块1 m×1 m的小样方, 在小样方中, 调查所有的植物样本, 记录群落内所有植物盖度, 每种植物在群落内盖度, 每种植物的株数, 植物的株高的最高值、最低值、平均值, 每种植物在各样方出现的频率。然后在每个小样方中, 去除地表杂物和凋落物后, 根据区域土壤环境特征与植物根系生长情况, 利用内径为7 cm的切根钻采集土壤层0—15 cm, 15—30 cm, 30—45 cm的原状土柱。三个小样方每个土壤层的土壤分别混合为一个重复, 总计5个重复。每个重复封入自封袋, 放置于低温泡沫盒中, 带回实验室进行处理。
1.4 室内样品分析将采集回的土壤过1 mm筛, 分离土壤与杂物, 将土壤保存在4℃下, 尽快监测土壤微生物群落。土壤微生物种类和数量利用磷脂脂肪酸(PLFAs)方法进行测定[18—20]。利用氢氧化钾和甲醇溶液提取新鲜土壤并甲酯化, 醋酸溶液中和后利用正己烷萃取, 正己烷挥发后加入同等体积的正己烷甲基丁基醚, 再加入内标的19 ∶ 0然后进行测定, 测定的仪器为Agilent 6890N型气象色谱仪。以19:0为内标的微生物脂肪酸估算如(表 1), 由于高寒沙地土壤贫瘠, 原生生物和藻类分布极少, 极难检测到, 因此只考虑细菌、真菌、放线菌群落[25—27]。
| 微生物类型Microbial groups | 磷脂脂肪酸标记Phospholipid fatty acid signature |
| 细菌Bacteria in general | 12∶0, 14∶0, 16∶0, i19∶0, 20∶0 |
| 革兰氏阳性细菌Gram-positive bacteria | a16∶0, i16∶0, a17∶0, i17∶0, i18∶0 |
| 革兰氏阴性细菌Gram-negative bacteria | i15∶0 3 OH, 16∶1ω9c, i17∶0 3 OH, 17∶1ω8c |
| 假单胞杆菌Pseudomonas spp. | 18∶1ω7c |
| 好氧细菌Aerobes | i14∶0, a14∶0, 15∶0 2OH, 15∶0 3 OH, a15∶0, i15∶0 |
| 硫酸盐还原细菌Sulfate-reducing bacterium | 10Me16∶0 |
| 甲烷氧化菌Methane-oxidizing bacteria | 16∶1ω5c |
| 黄杆菌Flavbacterium balustinum | i17∶1ω7 |
| 放线菌Actinomycetes | 10Me17∶0, 10Me18∶0 |
| 纤维菌属Cellulomonas | 11Me18∶ 1ω7c |
| 伯克霍尔德菌Burkholderia | cy19∶0ω8c |
| 真菌Fungi | 18∶3ω6c(6, 9, 12), 18∶1ω9c |
| 嗜热解氢杆菌Hydrogenobacter | 18∶0 |
| 原生生物Protozoa | 20∶4ω6, 9, 12, 15c |
对植物物种丰富度指数、生态优势度指数、物种多样性指数、均匀度指数进行分析, 比较老芒麦生长过程中植物多样性的变化情况。
植物物种重要值[28]:
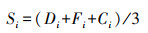
|
式中, Si、Di、Fi、Ci分别为第i个物种的重要值、相对多度、相对频度、相对盖度。相对多度等于第i个物种的个体数除以群落中所有物种的个体数的和; 相对频度等于第i个物种在统计样方中出现的次数除以所有种出现的总频数, 相对盖度等于第i个物种在群落中的盖度除以群落总的盖度。
群落物种丰富度指数[29]:
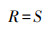
|
式中, R为植物群落物种丰富度指数, S为群落中的植物总物种数。
生态优势度指数(Simpson指数)[30]:
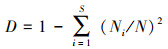
|
式中, D为Simpson指数, N为植物群落中全部种的总个体数, Ni为第i种的个体数。
物种多样性指数(Shannon-Wiener指数)[31]:
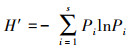
|
式中, H′为植物群落Shannon-Wiener指数, Pi=Ni/N, Pi为群落中第i种个体数占群落中全部种总个体数的比例。
均匀度指数(Pielou指数)[30]:
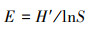
|
式中, E为植物群落Pielou指数。
1.5.2 微生物群落分析比较土壤微生物多样性指标, 包括丰富度指数、生态优势度指数、物种多样性指数、均匀度指数。其中微生物丰富度指数(R)[29]:
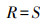
|
式中, R为微生物群落物种丰富度指数, S为群落中微生物群落中总脂肪酸种类。
生态优势度指数(D:Simpson指数)[30]:
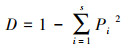
|
式中, D为微生物群落Simpson指数, Pi=Ni/N, Ni为i类脂肪酸个数, N为该实验中总脂肪酸个数。
物种多样性指数(H′:Shannon-Wiener指数)[31]:
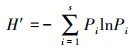
|
式中, H′为微生物群落Shannon-Wiener指数, Pi=Ni/N, Ni为i类脂肪酸个数, N为该实验中总脂肪酸个数。
均匀度指数(E:Pielou指数)[29]:
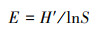
|
式中, E为微生物群落Pielou指数, H′为Shannon-Wiener指数, S为群落中微生物群落中总脂肪酸种类。
1.6 数据处理所有数据均采用均值和标准差进行表述。植物群落盖度、植物群落物种多样性、微生物群落多样性恢复不同年限之间的差异用单因素方法进行比较, 并且利用最小显著差异(L.S.D)方法确定显著水平, 显著水平为5%。所有数据均采用SPSS 26.0分析, 利用Origin 2019b作图。
2 结果与分析 2.1 植被恢复过程中植物多样性的变化连续三年的植被恢复过程中, 对照空白沙地上(CO), 仅有川西当归(Angelica sinensis)、早熟禾(Poa pratensis)、川甘亚菊(Ajania potaninii)、赖草(Leymus secalinus)和以礼草(Roegneria hirsuta)5种植物, 以礼草和赖草重要值最高。自种植老芒麦进行植被恢复以来, 老芒麦的重要值在群落内一直最高, 并且逐年增加, 而赖草与以礼草的重要值逐年减少。自老芒麦进行植被恢复开始, 植物种类逐渐增多, 恢复的第一年植物物种较空白沙地增加了4种, 至恢复的第3年, 群落内能调查到植物物种总量已经增加至18种, 物种数量比对照组增加了13种, 但恢复过程中川甘亚菊从群落中消失, 因此三年后植物种类相较于对照组新增了14个新物种。在恢复的第3年, 新定植的问荆(Equisetum arvense)、沙蒿(Artemisia desertorum)和川西景天(Sedum rosei)的重要值已经高于空白沙地的赖草和以礼草。随着恢复年限增加, 植物物种数量增多, 老芒麦重要值增加, 沙地原生物种重要值变小(图 1)。

|
| 图 1 不同恢复年限植物物种与重要值的变化 Fig. 1 Variations of plant species and important value in different recover year CK:对照组Control, 未种植老芒麦的空白沙地; 空白表示该物种在样方中无分布 |
空白沙地植物群落盖度极低, 利用老芒麦进行植被恢复后, 植物群落盖度迅速增加, 在恢复的第3年, 植物群落盖度已经超过80%, 远高于空白沙地。在空白沙地上, 只有伞形科、菊科、禾本科, 并且主要以禾本科为主; 但是利用老芒麦开始植被恢复后, 群落中逐渐出现了其他科植物, 如菥蓂(Thlaspi arvense)、川西景天(Sedum rosei)等, 且其他科物种数量快速增加。随着老芒麦进行植被恢复年限的增加, 群落中禾本科与其他科的多度增加, 并且禾本科和其他植物多度在恢复第2年和第3年呈爆发式增长, 至恢复的第3年, 禾本科和其他科植物多度已经远远高于空白沙地(图 2)。

|
| 图 2 不同恢复年限植物群落总盖度、不同科物种数量与多度变化 Fig. 2 Variations of plant community coverage, species and abundance of different family in different recover year CK:对照组Control, 未种植老芒麦的空白沙地; 不同小写字母表示不同恢复年限植物群落盖度差异显著(P<0.05) |
老芒麦进行植被恢复后, 植物群落物种丰富度, 生态优势度指数和物种多样性指数都逐年增加, 并且恢复3年后已经高于空白沙地; 植物物种丰富度与物种多样性指数在恢复的第2年和第3年差异不显著, 生态优势度指数自恢复第1年与恢复第3年差异不显著, 但是都高于空白沙地。随着恢复年限增加, 植物群落物种均匀度指数减少, 恢复的第1年与空白沙地差异不显著, 但是恢复的第2年和第3年显著低于空白沙地(图 3)。

|
| 图 3 不同恢复年限植物群落物种多样性变化 Fig. 3 Variations of plant biodiversity in different recover year |
老芒麦进行植被恢复后, 土壤中细菌含量季节性变化加剧; 随着植被恢复年限的增加, 老芒麦种植增加了土壤细菌含量。空白沙地上, 细菌含量从生长季节初期(EG)至非生长季节后期(LN)变化相对平缓, 但是利用老芒麦进行植被恢复以后, 整个土壤层中的细菌含量在不同季节间大幅变化。老芒麦进行植被恢复增加了土壤中细菌含量, 在非生长季节后期, 土壤0—15 cm和15—30 cm层细菌含量由对照组的0.96 mol/g与1.17 mol/g增加至3年后的2.13 mol/g和3.31 mol/g, 相对增加了1.2倍与1.8倍; 其中革兰氏阴性菌含量增加了0.3 mol/g和0.88 mol/g, 革兰氏阳性菌含量增加了0.87 mol/g和1.26 mol/g(图 4)。

|
| 图 4 不同恢复年限各关键季节土壤细菌、革兰氏阴性菌、革兰氏阳性菌含量变化 Fig. 4 Variations of soil bacteria, G- and G+ populations with critical seasons in different recover year EG:生长季节前期Early growing season; MG:生长季节中期Middle growing season; LG:生长季节后期Late growing season; EN:非生长季节前期Early non-growing season; LN:非生长季节后期Late non-growing season |
真菌和放线菌含量在高寒半湿润沙地中含量极低, 老芒麦进行植被恢复后, 增加了土壤中真菌和放线菌含量。老芒麦进行植被恢复使非生长季节土壤中真菌含量增加, 特别是在非生长季节后期, 土壤0—15 cm和15—30 cm层土壤中真菌含量由0.31 mol/g和0.16 mol/g增加至3年后的0.47 mol/g和0.98 mol/g, 相对增加了0.5倍和5.1倍。老芒麦进行植被恢复也增加了非生长季节后期土壤放线菌含量, 土壤0—15 cm和15—30 cm层土壤中放线菌含量由0.1 mol/g和0.13 mol/g增加至3年后的0.25 mol/g和0.65 mol/g, 相对增加了1.5倍和4倍(图 5)。

|
| 图 5 不同恢复年限各关键季节土壤真菌、放线菌含量变化 Fig. 5 Variations of soil fungi and actinomycetes populations at every critical season in different recover year |
老芒麦进行植被恢复后, 土壤中微生物多样性在不同关键季节变化幅度较大, 微生物多样性变化剧烈。在植物生长旺盛的生长季节中期(MG), 表层土壤(0—15 cm)微生物丰度在空白沙地上为17, 植被恢复后, 在第2年迅速增加至27, 虽然第3年降低, 但是植被恢复后微生物丰度依然高于空白沙地; 在微生物含量较高的非生长季节后期, 表层和深层(30—45 cm)土壤微生物丰度随着植被恢复年限降低, 但是中层确呈增加趋势。植被恢复并没有大幅改变恢复年际间土壤微生物物种多样性指数, 特别是表层土壤, 随着植被恢复, 物种多样性指数并没有显著差异, 但是植被恢复使土壤微生物物种多样性指数在不同关键季节剧烈变化。老芒麦进行植被恢复, 使土壤微生物均匀度和生态优势度指数变化幅度变小(图 6)。

|
| 图 6 不同恢复年限微生物群落物种多样性变化 Fig. 6 Variations of microbial biodiversity in different recover year |
利用植被重建对沙化生态系统进行生态恢复, 植被恢复过程中植物群落物种多样性对于恢复过程长效保障具有非常重要的意义。老芒麦作为青藏高原东南缘的广布根茎型乡土草本[32], 是该地区植被恢复的理想物种。高寒半湿润沙地上植物多样性变化积极响应植被恢复, 植物物种多样性随着恢复年限增加而增加; 并且以乡土物种老芒麦作为植被恢复的物种, 植被恢复过程中老芒麦在群落中重要值逐年增加, 原空白沙地的物种重要值降低, 并且恢复3年后群落中新定植物种重要值已经高于原空白沙地。其原因可能是:首先, 老芒麦作为人工种植物种促进植被恢复过程, 随着恢复年限的增加, 老芒麦一直占据主要生态位, 重要值逐渐增加; 其次, 老芒麦生长过程中, 植被群落盖度增加, 遮盖作用阻挡了过多日晒以及保存了地表水分, 为地表其他植物定植提供了良好的生境, 在每年降雨等有利条件下, 更多植物进入并定植, 导致物种丰度增加; 最后, 原空白沙地上的原生物种生存空间被压缩, 没有较好的光照、水分等条件, 物种重要值进一步降低, 甚至退出生态系统中。在相似的研究中, 植被恢复增加了植物物种多样性, 并且引导物种作为建群种, 也在生态系统中占据重要生态位[33—35], 但这些研究中植物群落变化过程缓慢, 这是由于本研究地处于高寒半湿润地区, 每年具有较为丰沛的降水, 特别是每年生长季节雨热同季[23], 植物的生长和定植具有良好的水分环境, 而与半干旱地区环境差异较为明显[36]。也有研究认为植被恢复对植物物种丰度促进不明显, 主要是增加了植物群落盖度[37], 这可能是由于修复的生态类型差异导致的, 在其他研究中沙化草地物种分布和多样性远高于本研究沙化地, 而本研究中初始沙地植物物种数量极少, 后期植物选择和定植的条件更加良好, 因此物种数量增加迅速。
在高寒半湿润沙地进行植被恢复, 3年后, 黄帚橐吾(Ligularia virgaurea)、瞿麦(Dianthus superbus)、问荆、川西景天等对环境水分要求较高的植物在植物群落中出现且定植, 这表明经过3年的植被恢复, 生态系统环境水分条件得到较大的改善, 适宜于对水分要求较高的植物定植。这可能是由于本研究所处的试验区为高寒半湿润区域, 近几年的年降水量最高超过800 mm[38], 良好的水分条件为植物定植提供了条件。伴随着植被恢复, 群落中首次出现了1年生短命植物菥蓂(Thlaspi arvense), 藜(Chenopodium album)和川西景天, 短命植物主要是利用短期降水条件, 在适宜水分条件下快速完成生活史的植物[39], 这表明植被恢复增强了生态系统功能, 使短命植物能够适应生境, 在群落中定植。
3.2 微生物多样性对植被恢复的响应老芒麦进行植被恢复后, 土壤微生物含量季节性变化明显, 尤其在非生长季节土壤中微生物含量大幅增加。这与干旱地区沙化恢复中植物生长季节(夏季)土壤微生物含量最高的结论不同[40]。青藏高原东南缘[41]与青藏高原典型湿地[42]相关研究表明进入非生长季节时期土壤微生物含量最高, 而青藏高原北部地区[43]的研究则表明土壤微生物含量季节性变化不明显, 并且高寒草甸的研究表明4月土壤细菌含量最高[44], 该时期土壤中细菌和真菌含量最高[45], 部分研究认为其主要原因是土壤微生物数量受土壤养分限制[46], 并且在此基础上提出了高寒地区的微生物会在非生长季节代替植被将碳储存在土壤中[47], 但相关研究还需要继续深入探讨。而高寒沙化植被恢复下非生长季节土壤微生物含量较高的原因是:(1)研究区域、研究对象都存在一定差异, 本研究中所研究的对象主要是高寒半湿润的贫瘠沙地, 利用老芒麦进行植被恢复, 生长季节植物生长和土壤中的微生物争夺养分, 使微生物的生长和繁殖受到限制[48], 因此生长季节的微生物数量较低; (2)非生长季节, 植物群落高盖度所残余的凋落物归还至土壤中, 增加了微生物生长的基质, 为冬季分解食物链的微生物类群, 如细菌、真菌的增长提供了良好的环境[49], 导致非生长季节微生物数量增加。
老芒麦种植进行生态恢复后, 土壤微生物群落优势度减小, 是由于植被恢复过程中, 土壤中微生物功能群落出现了变化, 不同功能类型的微生物可能在不同的季节或土壤层中出现, 单一的优势功能类群在所有的微生物类群中普遍分布, 因此群落优势度减小, 特别是随着植物物种增多, 不同功能类群的植物定植在生态系统中, 使土壤中微生物不同功能类群出现, 微生物功能呈现多样化, 如分解食物网上的不同微生物类群、固氮类群、硝化和反硝化类群、厌氧类群等的出现使整个微生物群落的组成更加丰富, 分布更加均匀, 因此微生物群落均匀度指数变化幅度减小[50]。
4 结论在高寒半湿润沙地上, 利用老芒麦作为人工建植物种促进植被恢复过程, 使地表植物数量增加, 植物群落盖度增加, 植物群落物种多样性增加。植被恢复使土壤微生物群落在不同季节剧烈变化, 但是随着植被恢复, 土壤细菌、真菌、放线菌含量增加, 土壤微生物多样性增加, 物种优势度和均匀度减小。老芒麦恢复高寒半湿润沙地, 有效促进了植物群落物种多样性与土壤微生物群落多样性的初期恢复, 地表植被和土壤微生物功能提升将有助于高寒沙化生态恢复过程。
| [1] |
董世魁, 王学霞, 张勇. 青藏高原高寒草地植物-土壤系统的生物多样性及其对全球变化的响应. 北京: 科学出版社, 2018: 8-16.
|
| [2] |
Dong S K, Shang Z H, Gao J X, Boone R B. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau. Agriculture, Ecosystems & Environment, 2020, 287: 106684. |
| [3] |
Harris R B. Rangeland degradation on the Qinghai-Tibetan Plateau: a review of the evidence of its magnitude and causes. Journal of Arid Environments, 2010, 74(1): 1-12. DOI:10.1016/j.jaridenv.2009.06.014 |
| [4] |
Dong Z B, Hu G Y, Yan C Z, Wang W L, Lu J F. Aeolian desertification and its causes in the Zoige Plateau of China's Qinghai-Tibetan Plateau. Environmental Earth Sciences, 2010, 59(8): 1731-1740. DOI:10.1007/s12665-009-0155-9 |
| [5] |
李森, 董玉祥, 董光荣, 杨萍, 张春来. 青藏高原土地沙漠化区划. 中国沙漠, 2001, 21(4): 418-427. |
| [6] |
Li Q X, Jia Z Q, Liu T, Feng L L, He L. Effects of different plantation types on soil properties after vegetation restoration in an alpine sandy land on the Tibetan Plateau, China. Journal of Arid Land, 2017, 9(2): 200-209. DOI:10.1007/s40333-017-0006-6 |
| [7] |
古琛, 贾志清, 何凌仙子, 李清雪, 高娅, 张佳鹏, 王陇. 恢复年限对高寒中间锦鸡儿群落组成和多样性的影响. 草业科学, 2022, 39(7): 1303-1311. |
| [8] |
魏小星. 青海湖东沙化治理过程中土壤酶活性及养分含量特征. 草业科学, 2017, 34(7): 1408-1418. |
| [9] |
Zhang H W, Tian L H, Hasi E, Zhang D S, Wu W Y. Vegetation-soil dynamics in an alpine desert ecosystem of the Qinghai Lake watershed, northeastern Qinghai-Tibet Plateau. Frontiers in Environmental Science, 2023, 11: 195. |
| [10] |
胡金娇, 周青平, 吕一河, 胡健, 陈有军, 苟小林. 青藏高原东缘半湿润沙地典型生态恢复模式的效果比较研究. 生态学报, 2020, 40(20): 7410-7418. |
| [11] |
Gou X L, Hu J, Chen Y J, Wei X X, Du Z, Zhou Q P. The effect of artificial vegetation recovery on the soil nutrients and enzyme activities in subhumid desert land on the southeast Qinghai-Tibetan Plateau, China. Ecological Engineering, 2019, 139: 105528. DOI:10.1016/j.ecoleng.2019.06.023 |
| [12] |
Deng J J, Yin Y, Zhu W X, Zhou Y B. Variations in soil bacterial community diversity and structures among different revegetation types in the Baishilazi nature reserve. Frontiers in Microbiology, 2018, 9: 2874. DOI:10.3389/fmicb.2018.02874 |
| [13] |
Yu J, Liu F, Tripathi B M, Steinberger Y. Changes in the composition of soil bacterial and fungal communities after revegetation with Caragana microphylla in a desertified semiarid grassland. Journal of Arid Environments, 2020, 182: 104262. DOI:10.1016/j.jaridenv.2020.104262 |
| [14] |
Liu L C, Liu Y B, Hui R, Xie M. Recovery of microbial community structure of biological soil crusts in successional stages of Shapotou Desert revegetation, northwest China. Soil Biology and Biochemistry, 2017, 107: 125-128. DOI:10.1016/j.soilbio.2016.12.030 |
| [15] |
Hu Y G, Zhang Z S, Huang L, Qi Q, Liu L C, Zhao Y, Wang Z R, Zhou H K, Lv X Y, Mao Z C, Yang Y F, Zhou J Z, Kardol P. Shifts in soil microbial community functional gene structure across a 61-year desert revegetation chronosequence. Geoderma, 2019, 347: 126-134. DOI:10.1016/j.geoderma.2019.03.046 |
| [16] |
Fu B, Qi Y B, Chang Q R. Impacts of revegetation management modes on soil properties and vegetation ecological restoration in degraded sandy grassland in farming-pastoral ecotone. International Journal of Agricultural and Biological Engineering, 2015, 8(1): 26-34. DOI:10.15740/HAS/IJAE/8.1/26-30 |
| [17] |
Miao C, Bai Y X, Zhang Y Q, She W W, Liu L, Qiao Y G, Qin S G. Interspecific interactions alter plant functional strategies in a revegetated shrub-dominated community in the Mu Us Desert, China. Annals of Botany, 2022, 130(2): 149-158. DOI:10.1093/aob/mcac039 |
| [18] |
Tian L H, Wu W Y, Zhou X, Zhang D S, Yu Y, Wang H J, Wang Q Y. The ecosystem effects of sand-binding shrub Hippophae rhamnoides in alpine semi-arid desert in the Northeastern Qinghai-Tibet Plateau. Land, 2019, 8(12): 183. DOI:10.3390/land8120183 |
| [19] |
王明明, 刘新平, 何玉惠, 张铜会, 魏静, 车力木格, 孙姗姗. 科尔沁沙地封育恢复过程中植物群落特征变化及影响因素. 植物生态学报, 2019, 43(8): 672-684. |
| [20] |
常学向, 陈怀顺, 李志刚, 张红萍. 西藏雅鲁藏布江流域典型沙漠区生态修复植物群落物种多样性. 中国沙漠, 2021, 41(6): 187-194. |
| [21] |
Li S C, Zhang Y L, Wang Z F, Li L H. Mapping human influence intensity in the Tibetan Plateau for conservation of ecological service functions. Ecosystem Services, 2018, 30: 276-286. DOI:10.1016/j.ecoser.2017.10.003 |
| [22] |
Chen H, Ju P J, Zhu Q A, Xu X L, Wu N, Gao Y H, Feng X J, Tian J Q, Niu S L, Zhang Y J, Peng C H, Wang Y F. Carbon and nitrogen cycling on the Qinghai-Tibetan Plateau. Nature Reviews Earth & Environment, 2022, 3(10): 701-716. |
| [23] |
苟小林, 刘文辉, 陈有军, 周蓉, 周青平. 植物沙障不同种植模式对川西北沙地的恢复效应. 草业学报, 2019, 28(6): 33-44. |
| [24] |
Hu Y F, Shu X Y, He J, Zhang Y L, Xiao H H, Tang X Y, Gu Y F, Lan T, Xia J G, Ling J, Chen G D, Wang C Q, Deng L J, Yuan S. Storage of C, N, and P affected by afforestation with Salix cupularis in an alpine semiarid desert ecosystem. Land Degradation & Development, 2018, 29(1): 188-198. |
| [25] |
Steinberger Y, Zelles L, Bai Q Y, von Lützow M, Munch J C. Phospholipid fatty acid profiles as indicators for the microbial community structure in soils along a climatic transect in the Judean Desert. Biology and Fertility of Soils, 1999, 28(3): 292-300. DOI:10.1007/s003740050496 |
| [26] |
刘微, 王树涛, 陈英旭, 吴伟祥, 王璟. 转Bt基因水稻根际土壤微生物多样性的磷脂脂肪酸(PLFAs)表征. 应用生态学报, 2011, 22(3): 727-733. |
| [27] |
Stone D, Blomkvist P, Hendriksen N B, Bonkowski M, Jørgensen H B, Carvalho F, Dunbar M B, Gardi C, Geisen S, Griffiths R, Hug A S, Jensen J, Laudon H, Mendes S, Morais P V, Orgiazzi A, Plassart P, Römbke J, Rutgers M, Schmelz R M, Creamer R E. A method of establishing a transect for biodiversity and ecosystem function monitoring across Europe. Applied Soil Ecology, 2016, 97: 3-11. DOI:10.1016/j.apsoil.2015.06.017 |
| [28] |
方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [29] |
尚玉昌. 普通生态学 (3版). 北京: 北京大学出版社, 2010: 272-365.
|
| [30] |
Pielou E C. Mathematical ecology. New York: John Wiley and Sons Incorporation, 1975: 115-142.
|
| [31] |
Magurran A E. Ecological diversity and its measurement. Princeton, NJ: Princeton University Press, 1988.
|
| [32] |
王传旗, 刘文辉, 张永超, 周青平. 野生老芒麦苗期耐旱性研究. 草业学报, 2021, 30(8): 127-136. |
| [33] |
Zhang Y Y, Zhao W Z. Vegetation and soil property response of short-time fencing in temperate desert of the Hexi Corridor, northwestern China. Catena, 2015, 133: 43-51. |
| [34] |
Li X R, Zhang Z S, Tan H J, Gao Y H, Liu L C, Wang X P. Ecological restoration and recovery in the wind-blown sand hazard areas of Northern China: relationship between soil water and carrying capacity for vegetation in the Tengger Desert. Science China Life Sciences, 2014, 57(5): 539-548. |
| [35] |
Chen D L, Zhang B, Fadda C, Jarvis D, Bergamini N, Han G D, Zhao M L, Bai K Y, Zhang Z W. Spontaneous grassland recovery on abandoned croplands in Northern China: different vegetation patterns in desert and typical steppe. Science of the Total Environment, 2021, 790: 148155. |
| [36] |
吴汪洋, 张登山, 田丽慧, 张明远, 周鑫. 近10年青海湖东沙地人工植被群落特征. 生态学报, 2019, 39(6): 2109-2121. |
| [37] |
王新源, 兰芳芳, 马仲武, 陈萍, 唐霞, 连杰, 王旭洋, 曲浩. 治沙措施通过植物群落的质量效应驱动土壤因子变化——以玛曲高寒草甸沙化区为例. 生态学报, 2023, 43(1): 70-81. |
| [38] |
帅林林, 周青平, 陈有军, 苟小林, 周蓉. 高寒半湿润沙地草本修复期土壤微生物变化研究. 草业学报, 2019, 28(9): 11-22. |
| [39] |
袁素芬, 唐海萍. 短命植物生理生态特性对生境的适应性研究进展. 草业学报, 2010, 19(1): 240-247. |
| [40] |
石万里, 王辉, 马维伟. 沙区植被恢复对土壤微生物量及活性的影响. 中国沙漠, 2017, 37(3): 507-513. |
| [41] |
Zhang L, Wang A, Yang W Q, Xu Z F, Wu F Z, Tan B, Liu Y, Chen L H. Soil microbial abundance and community structure vary with altitude and season in the coniferous forests, China. Journal of Soils and Sediments, 2017, 17(9): 2318-2328. |
| [42] |
孙飞达, 李飞, 陈文业, 朱灿, 周俗, 苟文龙, 刘琳, 周冀琼. 若尔盖退化高寒湿地土壤理化性质、酶活性及微生物群落的季节动态. 生态学报, 2020, 40(7): 2396-2406. |
| [43] |
高静, Said Muhanmmad, 岳琳艳, 何永涛, 斯确多吉, 张宪州, 孔维栋. 藏北高原草甸土壤固碳微生物群落特征随海拔和季节的变化. 生态学报, 2018, 38(11): 3816-3824. |
| [44] |
李洋, 王毅, 韩国栋, 孙建, 汪亚峰. 青藏高原高寒草地土壤微生物量碳氮含量特征及其控制要素. 草业学报, 2022, 31(6): 50-60. |
| [45] |
Xu B, Wang J N, Wu N, Wu Y, Shi F S. Seasonal and interannual dynamics of soil microbial biomass and available nitrogen in an alpine meadow in the eastern part of Qinghai-Tibet Plateau, China. Biogeosciences, 2018, 15(2): 567-579. |
| [46] |
França L, Sannino C, Turchetti B, Buzzini P, Margesin R. Seasonal and altitudinal changes of culturable bacterial and yeast diversity in Alpine forest soils. Extremophiles, 2016, 20(6): 855-873. |
| [47] |
Gutiérrez-Girón A, Rubio A, Gavilán R G. Temporal variation in microbial and plant biomass during summer in a Mediterranean high-mountain dry grassland. Plant and Soil, 2014, 374(1): 803-813. |
| [48] |
Cui Y X, Fang L C, Guo X B, Han F, Ju W L, Ye L P, Wang X, Tan W F, Zhang X C. Natural grassland as the optimal pattern of vegetation restoration in arid and semi-arid regions: evidence from nutrient limitation of soil microbes. Science of the Total Environment, 2019, 648: 388-397. |
| [49] |
Sorensen P O, Beller H R, Bill M, Bouskill N J, Hubbard S S, Karaoz U, Polussa A, Steltzer H, Wang S, Williams K H, Wu Y X, Brodie E L. The snowmelt niche differentiates three microbial life strategies that influence soil nitrogen availability during and after winter. Frontiers in Microbiology, 2020, 11: 871. |
| [50] |
Herrera Paredes S, Lebeis S L. Giving back to the community: microbial mechanisms of plant-soil interactions. Functional Ecology, 2016, 30(7): 1043-1052. |