 2023, Vol. 43
2023, Vol. 43文章信息
- 张署军, 尹本丰, 张庆, 陆永兴, 臧永新, 沙亚古丽·及格尔, 殷进飞, 张元明, 买买提明·苏来曼
- ZHANG Shujun, YIN Benfeng, ZHANG Qing, LU Yongxing, ZANG Yongxin, SHAYAGULI·Jigeer, YIN Jinfei, ZHANG Yuanming, MAMTIMIN·Sulayman
- 荒漠藓类植物齿肋赤藓对4年积雪深度变化的生理生化响应
- Ecophysiological and biochemical responses of desert moss Syntrichia caninervis to changes of snow depth in four years
- 生态学报. 2023, 43(2): 838-848
- Acta Ecologica Sinica. 2023, 43(2): 838-848
- http://dx.doi.org/10.5846/stxb202112063447
-
文章历史
- 收稿日期: 2021-12-06

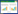




2. 中国科学院新疆生态与地理研究所, 荒漠与绿洲生态国家重点实验室, 乌鲁木齐 830011;
3. 中国科学院大学, 北京 100049
2. State Key Laboratory of Desert and Oasis Ecology, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
干旱和半干旱区作为全球最脆弱的生态系统之一, 对气候变化异常敏感。水分作为该地区限制植物生长的主要因素, 深刻影响着土壤的水文过程、养分循环及植物群落的结构与组成[1—2]。与维管植物相比, 生物土壤结皮凭借更强的耐旱性、较小的水分依赖性, 强抗辐射能力, 使得其在干旱和半干旱地区广泛发育, 并形成了与维管植物镶嵌分布的地表景观[3—4]。生物土壤结皮是由藻类、真菌、地衣、苔藓及其分泌物等与土壤颗粒相互胶结而形成的一种复合体, 能够在高温、低温、高辐射等极端环境条件下生存, 在部分地区占到旱地表面积的70%[5—6]。在荒漠地表稳定性、土壤肥力、水文过程、养分循环及维管植物建成等方面发挥重要作用, 因此也被称为地球的“活皮肤”[7—12]。
藓类结皮作为生物土壤结皮的重要组成部分, 是生物土壤结皮发育过程中的高级阶段。研究表明, 藓类植物在干旱、极端高低温、强辐射等环境下能够通过调节其形态和生理生化等特征来维持其基本的生理功能[13—14]。在形态方面, Wu等[15]在齿肋赤藓叶倾角对复水过程响应的研究中发现, 不同部位的叶倾角对复水过程具有不同的响应, 植株顶部的叶伸展速度与角度明显强于叶基部。藓类植物叶片毛尖不仅具有反射强光的作用, 还可以通过叶片毛尖汲取环境的水分[16—18]。在生理方面, 藓类植物对水分的时空变化异常敏感, 在水分适宜环境下能够快速恢复其生理活性[7]。在干旱、强紫外辐射等环境下, 藓类植物的渗透调节物质含量以及抗氧化酶活性能够随着环境胁迫强度的增加而增加, 以此来增加细胞的渗透压、及时清除细胞内活性氧, 缓解环境胁迫对植物的伤害[19—20]。
初冬和早春冻融季节积雪的融化和低温高湿的环境不仅为藓类植物的生长提供了适宜的环境, 还能改变土壤温度和水分, 减少土壤的冻害程度[21]。研究表明, 古尔班通古特沙漠齿肋赤藓的生物固碳主要集中在初冬和早春冻融季节, 占到全年固碳量的50%左右[22]。近些年来, 诸多学者也对荒漠藓类植物生理生化特征对积雪变化的响应做了大量研究。研究发现, 冬季适量积雪能够改善藓类植物的光合生理特性, 在面临冻融循环时, 藓类植物能够迅速脱水, 能够减缓细胞冻害的程度[23—24]。谢敏等[25]在真藓(Bryum argenteum)和土生对赤藓(Didymodon vinealis)对降雪的生理响应中发现, 2种藓类植物的光合色素、可溶性蛋白含量随着积雪的增加而增加, 而丙二醛(MDA)和可溶性糖含量降低, 并且土生对齿藓的生理变化更为强烈。Hui等[21]采用室内模拟积雪处理对生物土壤结皮的生理生化进行研究, 发现土壤含水量与积雪深度成正相关关系, 随着积雪深度的适度增加, 藓类植物的光合色素和可溶性蛋白含量增加, MDA和可溶性糖含量下降。Zhang等[26]通过一年野外梯度积雪深度变化对荒漠藓类齿肋赤藓的生理生化特性的影响的研究中发现, 积雪深度、积雪覆盖时间以及两者的交互作用对齿肋赤藓的生理生化特性有显著的影响, 可溶性蛋白含量随积雪深度的增加而增加, 积雪融化期的增加最为明显, 而MDA含量和抗氧化酶活性随着积雪深度的增加而减少。因此, 藓类植物能够通过调节体内的渗透调节物质含量、抗氧化酶活性及相关基因的表达来适应积雪的变化, 不同物种间存在差异。但这些研究均是短周期积雪深度(一次降雪或一年降雪), 关于长期积雪深度变化将如何影响荒漠藓类植物的生理适应性的研究还很缺乏。
基于此, 本研究提出以下科学问题: 长期积雪深度变化将如何影响荒漠藓类植物齿肋赤藓生理生化特征?为解答这一科学问题, 选取冬季具有稳定积雪期的古尔班通古特沙漠为研究区, 以该沙漠优势藓类植物齿肋赤藓为研究对象, 通过设置去除积雪(-S)、自然积雪(S)、2倍积雪(2S)和3倍积雪(3S) 4个积雪梯度, 探究4年持续积雪深度变化后齿肋赤藓生理生化特征, 以期为全球气候变化下荒漠生态系统稳定和物种多样性保护提供重要科学支撑。
1 材料与方法 1.1 研究区概况古尔班通古特沙漠(44°11′— 46°20′N, 84°31′—90°00′E;海拔300—600 m)位于新疆北部的准噶尔盆地, 面积约4.88×104 km2, 是我国最大的固定和半固定沙漠。该沙漠年均温度6—10℃, 年积温最高可达3500℃, 年平均降水量一般不超过150 mm, 在沙漠腹地仅有70—100 mm, 年均潜在蒸发量高达2000 mm以上。该沙漠冬季具有稳定的积雪, 积雪厚度通常在20—30 cm, 覆盖持续100—160d。由于该沙漠特殊的地理位置和生存环境, 使得耐干旱性强的植被类型在该地区广泛分布, 例如白梭梭(Haloxylon persicum)、梭梭(Haloxylon ammodendron)等小乔木和蒙古沙拐枣(Calligonum mongolicum)、膜果麻黄(Ephedra przewalskii)等小灌木群落。早春积雪融化能够为短命植物、一年生草本植物的萌发和生长提供了充足的水分, 使草本植物群落广发发育[7, 27]。此外, 该沙漠地表具有发育良好的生物土壤结皮, 与维管植物镶嵌分布, 形成了斑块化的地表景观[28]。
1.2 实验设计于2017年11月在古尔班通古特沙漠腹地选取地势比较平坦、且藓类结皮发育良好的丘间低地设置100 m×100 m大样方, 为避免人类活动及动物干扰, 在样地外围用铁丝网进行围封。在大样地内设置1.5 m×1.5 m小样方, 采用随机分布的方式设置去除积雪(-S)、自然积雪(S)、2倍积雪(2S)和3倍积雪(3S) 4个积雪梯度, 每个处理5个重复, 共计20个小样方(图 1)。为避免不同处理间相互影响, 每个小样方之间距离在2 m以上。在每个处理样方内地表下5 cm处埋设ECH2O监测系统(5TM, Li-COR, Lincoln, USA) 用于监测土壤的水分和温度, 每间隔30 min自动记录一次。

|
| 图 1 实验样地景观和积雪处理 Fig. 1 Landscape of experiment plot and snow treatment |
去除积雪处理样方, 采用长宽各为1.5 m, 高30 cm不锈钢钢架(为保证钢构架稳定插入土壤30 cm, 高出地面30 cm), 架子上方采用透光亚克力板作为遮雪材料(图 1), 同时在保证不影响环境温度的情况下在钢架四周用通风尼龙网进行遮挡防止侧面雪的进入, 遮雪处理于每年开始积雪积累前的11月中旬进行, 并于翌年3月中旬积雪融化期拆除, 不影响自然降雨。在样地周围用地板格固定与样方大小一致的面积继续积雪收集, 在每次降雪后, 根据当次的自然降雪量从与积雪样方同等大小的空地收集积雪, 通过筛子向样方内均匀加入2倍积雪和3倍积雪(图 1)[26]。2017年—2020年4年自然积雪厚度分别为: 23 cm、18.6 cm、20 cm、12 cm。直至2021年3月积雪完全融化后第二天, 用PAM-2500 (Walz, Hernz, Germany)原位测定所有处理齿肋赤藓的荧光活性, 荧光活性测定完成后, 采用双面刀快速将齿肋赤藓地上部分的茎叶进行切除, 将切除后的样品放入液氮罐中带回实验室, 放入-80℃超低温冰箱中保存备用。
1.3 齿肋赤藓生理指标测定 1.3.1 相对含水量测定植株含水量的测定采用烘干法, 将处理好的新鲜齿肋赤藓茎叶部分用电子天平称量(精度0.0001 g) 称量鲜重M1, 随后将其放入75℃烘箱中烘干48 h至恒重, 称量干重M2。按照以下公式计算齿肋赤藓的相对含水量WC:
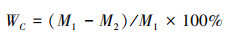
|
于2021年3月利用便携式调制叶绿素荧光仪PAM-2500 (Walz, Hernz, Germany) 测定齿肋赤藓的叶绿素荧光参数。由于减雪处理齿肋赤藓植物处于休眠状态, 为准确评估长期积雪变化下齿肋赤藓的实际光合活性, 在测定前24 h给予了减雪处理齿肋赤藓与自然降雪等量的降水让其恢复光合活性[29]。在测定之前用遮荫盒将整个PVC管遮荫。经过1 h充分暗适应后, 于波峰为650 nm, 强度为0.1 μmol m-2 s-1的红光下测得初始荧光(F0), 于强度为10 000 μmol m-2 s-1的饱和脉冲光下测得最大荧光(Fm), 脉冲时间为0.8 s;在光照强度为250 μmol m-2 s-1的光化光下测定稳态荧光(Ft)。待Ft稳定后, 打开一次饱和脉冲光测得光适应后的最大荧光(Fm′)。根据以上参数可以计算光系统Ⅱ(PSⅡ)的最大光化学效率FV/Fm=(Fm-F0)/Fm、光适应下PSⅡ的实际光化学效率φPSII=Fm′-Ft/Fm′ [30—31]。
1.3.3 渗透调剂物质含量的测定本研究主要测定游离脯氨酸、可溶性糖和可溶性蛋白三种渗透调节物质, 所有指标均用电子天平准确称取0.1500 g (精度0.0001 g) 齿肋赤藓茎叶样品。参照Monreal等的茚三酮方法测定植物体内游离脯氨酸的含量[32]。将样品液氮冷冻后进行研磨, 加入1 mL 3%的磺基水杨酸于沸水浴中浸提10 min, 冷却后用离心机(25℃、12000 r/min)离心5 min, 吸取上清液0.2 mL于2 mL试管内, 加入0.2 mL冰乙酸和0.3 mL茚三酮显色液于沸水浴中加热40 min, 冷却后加入1 mL甲苯, 漩涡震荡20—30 s, 静置后吸取甲苯层, 以0号管为自然降雪, 520 nm进行比色读取吸光度值。
采用考马斯亮蓝G-250法测定可溶性蛋白含量。将样品液氮冷冻研磨后, 加入1.35 mL蒸馏水提取, 用离心机(25℃、12000 rpm)离心10 min, 吸取上清液0.06 mL于2 mL试管内, 加蒸馏水使其定容到1.8 mL。其次吸取清液0.2 mL于2 mL试管内加入1 mL考马斯亮蓝G-250试剂, 混匀后静置3 min, 于595 nm下读取吸光度值[31]。
采用蒽酮法测定可溶性糖含量。将样品液氮冷冻研磨后, 加入1.5 mL蒸馏水提取, 随后将其放于放入50℃水浴锅中水浴20 min, 冷却后用离心机(25℃、12000 rpm)离心10 min, 吸取上清液0.2 mL于2 mL试管内, 加入1 mL蒽酮, 沸水浴中加热10 min, 冷却后在520 nm进行比色读取吸光度值[33]。
1.3.4 MDA含量和抗氧化酶活性的测定(1) MDA含量的测定
釆用硫代巴比妥酸(TBA)比色法对植物组织中的MDA含量进行测定。称取0.1500 g齿肋赤藓样品, 液氮冷冻研磨后加1 ml提取液(10%三氯乙酸(TCA)+0.25%TBA)进行冰浴匀浆, 95℃水浴加热30 min, 在离心机(25℃12000 rpm)中离心10 min, 取上清液。用紫外可见分光光度计测定样品在波长532 nm和600 nm处的吸光度[34]。
(2) 抗氧化酶活性的测定
称取0.1500 g齿肋赤藓样品, 液氮冷冻研磨后加1 ml提取液进行冰浴匀浆。随后在离心机(4℃ 12000 rpm)中离心10 min, 取上清液测定超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性。SOD活性采用羟胺法进行测定, 通过测定SOD抑制氮蓝四氮唑光化学还原能力来测定SOD活性, 并在560 nm处测定吸光度。POD活性采用愈创木酚法进行测定, 将H2O2反应混合物加入到上清液中, 在470 nm处测定吸光度。CAT活性采用可见光法进行测定, 在240 nm波长处测定吸光度[26]。
1.4 统计分析利用Microsoft Excel对原始数据常规处理。使用SPSS 20.0分别对4种不同积雪处理的数据进行正态性检验和方差齐性检验, 采用单因素方差分析(One-way ANOVA)和多重比较(LSD)对4种积雪处理下的齿肋赤藓生理指标进行方差分析, 使用R 4.1.2对植株含水量和土壤温度与相关生理指标进行了相关性分析和作图, 主要用到包为psych和corrplot。最后用Origin 20.0软件对数据进行绘图。
2 结果与分析 2.1 积雪深度对齿肋赤藓植株含水量的影响积雪的存在增加了齿肋赤藓的植株含水量, 表现出随积雪深度的增加齿肋赤藓植株含水量先上升后趋于稳定的趋势(图 2)。2倍积雪和3倍积雪植株含水量均显著高于自然降雪, 同时3种积雪处理均显著高于去除积雪(P < 0.05)。而2倍积雪和3倍积雪处理下齿肋赤藓的植株含水量差异不显著(P>0.05)。

|
| 图 2 4年积雪变化对齿肋赤藓植株含水量的影响 Fig. 2 Changes of water content of S. caninervis after 4 years snow cover treatments 不同字母表示各处理间的差异显著(P < 0.05) |
方差分析结果显示, 4年积雪变化显著影响了齿肋赤藓的最大光化学量子产量Fv/Fm和实际光化学效率Y(II)。与自然降雪相比, 去除积雪显著降低了齿肋赤藓的Fv/Fm和Y(II), 但积雪深度的增加并没有显著增加齿肋赤藓的Fv/Fm, 且随着积雪深度的增加其Y(II)出现递减趋势, 3倍积雪与自然降雪相比达到了显著水平(P < 0.05)(图 3)。

|
| 图 3 4年积雪对齿肋赤藓植株光系统II(PSII)最大光化学效率(Fv/Fm)和实际光化学效率(Y(II))的影响 Fig. 3 Changes of maximum photochemical efficiency (Fv/Fm) and actual photoch-emical efficiency ((Y(II)) of PSII of S. caninervis after 4 years snow cover treatments |
由图 4可知, 积雪量的变化显著影响了齿肋赤藓植株内的游离脯氨酸含量、可溶性糖和可溶性蛋白含量。随着积雪深度的增加, 齿肋赤藓植株游离脯氨酸含量呈现出逐渐下降的趋势, 除积雪去除处理和3倍积雪处理间达到差异显著外(P < 0.05), 积雪去除和2倍积雪与自然降雪相比均没有达到显著性水平(P>0.05)。与游离脯氨酸类似, 积雪去除显著增加了齿肋赤藓植株的可溶性糖和可溶性蛋白含量(P < 0.05), 但积雪量倍增与自然降雪无显著差异(P>0.05)。

|
| 图 4 4年积雪对齿肋赤藓植株渗透调节物质的影响 Fig. 4 Changes of osmoregulation substance of S. caninervis after 4 years snow cover treatments |
单因素方差分析结果显示, 积雪显著影响齿肋赤藓植株体内的MDA含量及SOD、POD、CAT 3种抗氧化酶活性。与自然降雪处理相比, 积雪去除处理的齿肋赤藓植株MDA含量及SOD、POD、CAT3种抗氧化酶活性显著增加(图 5)。除SOD随积雪深度增加而持续降低外, POD和CAT 2种抗氧化酶活性均与自然降雪差异不显著。与抗氧化酶活性随着积雪量增加活性逐渐降低或趋于平稳不同, 齿肋赤藓的MDA含量却表现出随着积雪量增加先降低后增加的趋势(图 5)。

|
| 图 5 4年积雪对齿肋赤藓植株丙二醛(MDA)含量及抗氧化酶活性的影响 Fig. 5 Changes of MDA content and antioxidant enzyme activity of S. caninervis after 4 years snow cover treatments |
由图 6可知, 土壤温度和植株含水量与齿肋赤藓的Fv/Fm和Y(II)呈现显著正相关, 与可溶性蛋白、可溶性糖、MDA含量及SOD、POD、CAT3种抗氧化酶活性呈现负相关关系。

|
| 图 6 齿肋赤藓植株含水量与生理特征的相关性分析 Fig. 6 Correlation analysis between plant water content and physiological characteristics of S. caninervis Temp: 土壤温度; WC: 植株含水量; Pro: 脯氨酸; Protein: 可溶性蛋白; Sugar: 可溶性糖; POD: 过氧化物酶; SOD: 超氧化物歧化酶; CAT: 过氧化氢酶; MDA: 丙二醛; 最大光化学量子产量(Fv/Fm); 实际光化学效率(Y(II)); 点的大小代表相关性, 点越大相关性越强;蓝色代表正相关, 红色代表负相关 |
水分作为干旱半干旱区植物生长的主要限制性因子, 显著影响了植物的生长发育。古尔班通古特沙漠作为中国西北荒漠区降水较少的区域之一, 其水分来源除降水外, 冬季的稳定积雪也是促进该地区植物生长重要水源。冬季积雪的融化能够显著增加荒漠藓类植物齿肋赤藓植株含水量, 使得处于休眠状态的齿肋赤藓在短时间内快速恢复其光合活性, 并通过调节植株体内的渗透调节物质含量和抗氧化酶活性来适应各种环境的变化[25, 35—36]。
3.1 积雪深度变化对齿肋赤藓荧光活性的影响光合作用作为机体生命活动的基础, 对植物的生长发育起着至关重要的作用, 其中叶绿素荧光作为光合作用的探针, 能够快速灵敏的探测植物光系统II对光能的吸收、传递等内在特性, 被广泛应用与植物光合活性的评价[37]。Fv/Fm作为PSII的最大光化学量子产量, 能够反映植物的最大光合潜能[38], Y(II)是PSII反应中心实际的光化学活性[39]。当藓类植物受到干旱、水淹、强辐射、高低温等胁迫时, Fv/Fm和Y(II)将会降低[29, 39—41]。冬季的积雪显著影响藓类植物的光合作用, 藓类植株的荧光活性随着积雪深度的增加呈现上升趋势, 但是持续的积雪量会引起藓类植物的荧光活性下降[21]。这在本研究中也得到证实, 与自然降雪相比, 长期积雪去除和3倍积雪均显著降低了齿肋赤藓的Fv/Fm和Y(II)。其实, 在冬季积雪稳定覆盖期, 由于极端低温齿肋赤藓会进行脱水休眠, 直到冻融季积雪融化可利用水分增加后齿肋赤藓会快速复水在短时间内恢复荧光活性[30]。然而, 对4年积雪去除处理进行自然积雪复水后, 其Fv/Fm和Y(II)仍很难恢复到正常水平。这说明长期冬季积雪去除营造的低温、低水分环境会对齿肋赤藓的光合活性造成不可逆的伤害, 从而影响光合电子的传递和光合磷酸化, 致使齿肋赤藓植株的Fv/Fm和Y(II)降低[42]。这可能由于冻融季是齿肋赤藓的最大固碳季, 而长期积雪去除造成齿肋赤藓年光合产物积累量小于消耗量, 引起齿肋赤藓长期碳饥饿所致[21, 43]。此外, 与自然积雪相比, 虽然加倍积雪没有显著影响齿肋赤藓的Fv/Fm, 但随着积雪量的增加齿肋赤藓的Y(II)呈下降趋势, 这可能是由于积雪融水过多致使齿肋赤藓处于腌渍状态, 导致齿肋赤藓叶片气孔关闭吸收CO2的能力降低, 进而对齿肋赤藓的生长产生负面影响[44—45]。
3.2 积雪深度变化对齿肋赤藓渗透调节物质的影响渗透调节物质作为植物响应环境胁迫的一个重要指标, 对于维持植物细胞的渗透压和调节植物体内活性氧的平衡具有重要作用[46—47]。在干旱和高低温胁迫中, 藓类植物为了缓解因胁迫造成的生理代谢不平衡, 通过失复水和调节植株体内渗透调节物质(脯氨酸、可溶性糖和可溶性蛋白)含量来维持正常的渗透压, 保证细胞的正常生理功能[48—50]。
脯氨酸作为重要的渗透调制物质之一, 常以游离状态分布于植物体内, 当植物受到环境胁迫时, 细胞内会通过积累体内脯氨酸含量来维持植物细胞内的渗透压平衡[51—52]。张静等[20]认为, 齿肋赤藓植物体内游离脯氨酸的含量随着降水的增加呈现降低的趋势, 但随着降水量的持续增加游离脯氨酸含量会止降反升, 以此来维持藓类稳定的植株含水量与膨压势。此外, 藓类植物体内游离脯氨酸含量与积雪的变化也存在显著的相关性, 随着积雪深度的增加真藓和土生对赤藓植株中游离脯氨酸含量呈现下降趋势[25]。在本研究中, 随着积雪深度的增加, 齿肋赤藓植株体内游离脯氨酸含量差异显著, 整体呈现下降趋势, 以此来降低细胞渗透压提高植物对水分的利用效率。这主要是由于积雪的融化为齿肋赤藓带来了充足的水分, 植物干旱胁迫减缓乃至消除所致。
可溶性糖作为生物体的重要成分和能量来源, 既能够为植物提供重要能量也能够通过作为重要渗透调节物质来调节细胞渗透压[53]。研究发现, 植物能够通过积累体内可溶性糖含量来增强植物对冬季低温和干旱的抵御能力[54]。已有研究表明, 稳定的积雪能够在冬季形成一个保温层缓解低温对植物的损伤, 且春季积雪融化又能够为藓类植物生长提供充足水分, 使其体内的可溶性糖含量下降[21]。在本研究中也得到了证实, 与自然降雪相比, 积雪去除处理导致齿肋赤藓可溶性糖含量增加, 这主要是因为积雪的去除致使齿肋赤藓遭受干旱和低温, 导致体内可溶性糖大量积累来提高细胞液浓度, 增强齿肋赤藓细胞的持水能力, 降低细胞质的冰点, 防止细胞损伤, 以此来低温胁迫[55—56]。
可溶性蛋白是以小分子状态存在的易溶于水或者其他溶剂的一类蛋白, 是植物体内重要的渗透调节物质, 与植物新陈代谢密切相关。此外, 部分功能蛋白酶是可溶性蛋白, 其含量的增加与植物的抗寒性密切相关[54]。已有研究表明, 积雪的存在为藓类植物提供适宜的水分条件和生长温度, 导致藓类植物的活性恢复, 新陈代谢加强, 可溶性蛋白含量升高[21]。然而本研究中, 齿肋赤藓植株可溶性蛋白含量随积雪深度的增加呈现降低的趋势, 与植株含水量呈现一种显著的负相关关系, 这与一次积雪导致齿肋赤藓植株可溶性蛋白含量下降结果一致[35], 但是与谢敏等[25]在真藓和土生对齿藓对积雪的生理响应中得到的结果相反。造成这一结果主要原因有以下几个方面, 首先, 齿肋赤藓在经历积雪期时, 雪层的存在为其创造了一种低辐射的适宜环境, 在经历融雪期后遭受到短期强光的辐射, 齿肋赤藓植株内参与UV-B防御系统以及新陈代谢能力降低, 导致齿肋赤藓植株可溶性蛋白降低。这在Hui等[47]在2种藓类植物生理生化特征对不同强度的紫外辐射响应的研究中得以证实。其次, 齿肋赤藓植株经历积雪期的低温以及水分胁迫, 导致齿肋赤藓蛋白质的合成降低或者降解速率增强, 植株可溶性蛋白含量降低[50]。最后, 藓类植物的反复复水过程中, 生理测定前与上次复水的间隔时间也是造成各研究间差异的重要原因之一。
3.3 积雪对齿肋赤藓MDA含量和抗氧化酶活性的影响丙二醛作为脂质过氧化的最终产物, 能够间接的反应细胞的损伤程度。当植物遭受环境胁迫时, 会引起细胞膜的膜脂过氧化反应, 产生大量的MDA。因此, MDA含量常用来衡量植物受到胁迫的大小[21, 57]。在本实验中, 积雪去除处理的齿肋赤藓植株MDA含量显著高于自然降雪和积雪倍增, 这主要是由于积雪的去除致使齿肋赤藓植株遭受干旱胁迫, 脂质过氧化更为严重。值得注意的是, 与2倍积雪相比, 3倍积雪处理下齿肋赤藓植株MDA含量显著升高, 这是由于积雪融水过多致使齿肋赤藓处于腌渍状态, 而此时的抗氧化酶活性较低, 消除自由基的能力下降, 导致膜脂过氧化程度加深, 其MDA含量升高。这一结论与Hui等[21]在腾格里沙漠研究冬季积雪深度变化对生物土壤结皮的生理特征影响中, 生物土壤结皮中MDA含量随冬季积雪深度的变化的趋势一致。
抗氧化防御系统作为衡量植物耐受性的最佳指标, 植物遭受胁迫之后, 抗氧化酶活性增加能够及时清除体内多余的活性氧(ROS)以此保护细胞免受伤害[58]。在人工藓类结皮对脱水-复水过程和热胁迫的生理响应的研究中发现, 干旱胁迫会导致抗氧化酶活性增加, 随着水分的增加抗氧化酶活性逐渐降低[59]。然而冬季积雪作为水分来源的重要途经之一, 导致藓类植物生存环境中的水分、温度、光照强度等因素存在显著的差异。因此, 藓类植物中的抗氧化酶活性发生显著的变化, 随着积雪量的增加藓类植株中的SOD、POD、CAT活性降低[26]。在本研究中, 抗氧化酶性随着积雪量的增加显著降低, 但不同抗氧化酶活性表现并不完全相同。对CAT活性来说, 随着积雪的增加CAT活性没有显著的变化, 说明CAT活性对积雪处理的敏感性低于SOD、POD活性, 这一结论已在Yin等[55]在齿肋赤藓生理特征对积雪量与冻融循环次数响应中得到了证实。此外, 与MDA含量结果相似, 3倍积雪齿肋赤藓中SOD、POD活性较自然降雪而言呈现上升趋势。这说明积雪的增加能够为藓类植物的生长提供充足的水分, 但长期过量积雪的增加对藓类植物的生长会产生不利影响。
4 结论综上所述, 4年积雪深度变化显著影响了荒漠藓类植物齿肋赤藓的生理生化特征。随着积雪深度的增加, 齿肋赤藓的植株含水量、Fv/Fm和Y(II)显著升高, 而游离脯氨酸、可溶性蛋白和MDA含量及SOD、POD、CAT活性均显著降低;Fv/Fm和Y(II)与土壤温度和植株含水量呈显著正相关, 渗透调节物质和抗氧化酶活性与土壤温度和植株含水量呈显著的负相关。说明积雪的存在对齿肋赤藓的生长具有正向的促进作用。同时, 积雪过量(3倍积雪)导致齿肋赤藓植株含水量、Fv/Fm和Y(II)降低, MDA含量及SOD、POD、CAT活性不同程度的上升趋势, 对齿肋赤藓的生长产生严重负面影响, 会导致齿肋赤藓在较短时间内难以恢复到自然的状态。
致谢: 感谢李永刚老师和张世航同学在实验数据处理过程中给予的帮助。| [1] |
Maestre F T, Salguero-Gomez R, Quero J L. It is getting hotter in here: determining and projecting the impacts of global environmental change on drylands. Philosophical Transactions of the Royal Society B: Biological Sciences, 2012, 367(1606): 3062-3075. DOI:10.1098/rstb.2011.0323 |
| [2] |
Zhang C H, Xi N X. Precipitation changes regulate plant and soil microbial biomass via plasticity in plant biomass allocation in grasslands: a meta-analysis. Frontiers in Plant Science, 2021, 12: 614968. DOI:10.3389/fpls.2021.614968 |
| [3] |
张元明, 王雪芹. 荒漠地表生物土壤结皮形成与演替特征概述. 生态学报, 2010, 30(16): 4484-4492. |
| [4] |
Li X R, Song G, Hui R, Wang Z R. Precipitation and topsoil attributes determine the species diversity and distribution patterns of crustal communities in desert ecosystems. Plant and Soil, 2017, 420(1/2): 163-175. |
| [5] |
张元明. 荒漠地表生物土壤结皮的微结构及其早期发育特征. 科学通报, 2005, 50(1): 42-47. DOI:10.3321/j.issn:0023-074X.2005.01.009 |
| [6] |
West N E. Structure and function of microphytic soil crusts in wildland ecosystems of arid to semi-arid regions. Advances in Ecological Research, 1990, 20: 179-223. |
| [7] |
Zhang Y M, Chen J, Wang L, Wang X Q, Gu Z H. The spatial distribution patterns of biological soil crusts in the Gurbantunggut Desert, Northern Xinjiang, China. Journal of Arid Environments, 2007, 68(4): 599-610. DOI:10.1016/j.jaridenv.2006.06.012 |
| [8] |
Chaudhary V B, Bowker M A, O'Dell T E, Grace J B, Redman A E, Rillig M C, Johnson N C. Untangling the biological contributions to soil stability in semiarid shrublands. Ecological Applications, 2009, 19(1): 110-122. DOI:10.1890/07-2076.1 |
| [9] |
Castillo-Monroy A P, Maestre F T, Delgado-Baquerizo M, Gallardo A. Biological soil crusts modulate nitrogen availability in semi-arid ecosystems: insights from a Mediterranean grassland. Plant and Soil, 2010, 333(1/2): 21-34. |
| [10] |
Bowker M A, Reed S C, Maestre F T, Eldridge D J. Biocrusts: the living skin of the earth. Plant and Soil, 2018, 429(1/2): 1-7. |
| [11] |
Elbert W, Weber B, Burrows S, Steinkamp J, Büdel B, Andreae M O, Pöschl U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nature Geoscience, 2012, 5(7): 459-462. DOI:10.1038/ngeo1486 |
| [12] |
Li X R, Zhang P, Su Y G, Jia R L. Carbon fixation by biological soil crusts following revegetation of sand dunes in arid desert regions of China: a four-year field study. CATENA, 2012, 97: 119-126. DOI:10.1016/j.catena.2012.05.009 |
| [13] |
Zheng Y P, Xu M, Zhao J C, Zhang B C, Bei S Q, Hao L H. Morphological adaptations to drought and reproductive strategy of the moss Syntrichia caninervis in the Gurbantunggut Desert, China. Arid Land Research and Management, 2011, 25(2): 116-127. DOI:10.1080/15324982.2011.554956 |
| [14] |
Li X R, Hui R, Zhang P, Song N P. Divergent responses of moss- and lichen-dominated biocrusts to warming and increased drought in arid desert regions. Agricultural and Forest Meteorology, 2021, 303: 108387. DOI:10.1016/j.agrformet.2021.108387 |
| [15] |
Wu N, Zhang Y M, Downing A, Aanderud Z T, Tao Y, Williams S. Rapid adjustment of leaf angle explains how the desert moss, Syntrichia caninervis, copes with multiple resource limitations during rehydration. Functional Plant Biology, 2014, 41(2): 168-177. DOI:10.1071/FP13054 |
| [16] |
陶冶, 张元明. 叶片毛尖对齿肋赤藓结皮凝结水形成及蒸发的影响. 生态学报, 2012, 32(1): 7-16. |
| [17] |
Pan Z, Pitt W G, Zhang Y M, Wu N, Tao Y, Truscott T T. The upside-down water collection system of Syntrichia caninervis. Nature Plants, 2016, 2(7): 16076. DOI:10.1038/nplants.2016.76 |
| [18] |
许红梅, 尹本丰, 李进, 张元明. 采自不同微生境的齿肋赤藓(Syntrichia caninervis)沙培植株生长与叶形态差异. 中国沙漠, 2015, 35(2): 373-379. |
| [19] |
Hui R, Li X R, Zhao R M, Tan H J, Jia R L. Physiological response of moss/cyanobacteria crusts along a precipitation gradient from semi-arid to arid desert in China. Plant and Soil, 2021, 468(1/2): 97-113. |
| [20] |
张静, 张元明. 模拟降雨对齿肋赤藓(Syntrichia caninervis)生理特性的影响. 中国沙漠, 2014, 34(2): 433-440. |
| [21] |
Hui R, Zhao R M, Liu L C, Li Y X, Yang H T, Wang Y L, Xie M, Wang X Q. Changes in winter snow depth affects photosynthesis and physiological characteristics of biological soil crusts in the Tengger Desert. Photosynthetica, 2018, 56(4): 1304-1312. DOI:10.1007/s11099-018-0838-0 |
| [22] |
吴林. 古尔班通古特沙漠生物结皮在地-气界面CO2交换中的作用[D]. 北京: 中国科学院大学, 2015.
|
| [23] |
Lenné T, Bryant G, Hocart C H, Huang C X, Ball M C. Freeze avoidance: a dehydrating moss gathers no ice. Plant, Cell & Environment, 2010, 33(10): 1731-1741. |
| [24] |
李刚, 刘立超, 高艳红, 赵杰才, 杨昊天. 降雪对生物土壤结皮光合及呼吸作用的影响. 中国沙漠, 2014, 34(4): 998-1006. |
| [25] |
谢敏, 回嵘, 刘立超, 杨昊天, 王艳莉, 魏文斐, 王雪芹. 沙漠生物土壤结皮中真藓(Bryum argenteum)和土生对齿藓(Didymodon vinealis)对降雪的生理响应. 中国沙漠, 2017, 37(1): 116-123. |
| [26] |
Zhang J, Zhang Y M. Ecophysiological responses of the biocrust moss Syntrichia caninervis to experimental snow cover manipulations in a temperate desert of central Asia. Ecological Research, 2020, 35(1): 198-207. DOI:10.1111/1440-1703.12072 |
| [27] |
周宏飞, 李彦, 汤英, 周宝佳, 徐宏伟. 古尔班通古特沙漠的积雪及雪融水储存特征. 干旱区研究, 2009, 26(3): 312-317. DOI:10.13866/j.azr.2009.03.009 |
| [28] |
张元明, 潘惠霞, 潘伯荣. 古尔班通古特沙漠不同地貌部位生物结皮的选择性分布. 水土保持学报, 2004, 18(4): 61-64. DOI:10.3321/j.issn:1009-2242.2004.04.016 |
| [29] |
Yin B F, Zhang Y M. Physiological regulation of Syntrichia caninervis Mitt. in different microhabitats during periods of snow in the Gurbantünggüt Desert, northwestern China. Journal of Plant Physiology, 2016, 194: 13-22. DOI:10.1016/j.jplph.2016.01.015 |
| [30] |
张静, 张元明. 冻融过程对生物结皮中齿肋赤藓叶绿素荧光特性的影响. 中国沙漠, 2011, 31(6): 1479-1487. |
| [31] |
尹本丰, 张元明, 娄安如. 灌丛移除对荒漠齿肋赤藓越冬过程中生理生化特性的影响. 植物生态学报, 2016, 40(7): 723-734. |
| [32] |
Monreal J A, Jiménez E T, Remesal E, Morillo-Velarde R, García-Mauriño S, Echevarría C. Proline content of sugar beet storage roots: response to water deficit and nitrogen fertilization at field conditions. Environmental and Experimental Botany, 2007, 60(2): 257-267. DOI:10.1016/j.envexpbot.2006.11.002 |
| [33] |
李合生, 孙群, 赵世杰, 章文华. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2000: 260-261.
|
| [34] |
尹本丰. 荒漠区齿肋赤藓(Syntrichia caninervis Mitt)对微生境差异的生态与生理生化适应性[D]. 北京: 中国科学院大学, 2015.
|
| [35] |
尹本丰, 张元明. 荒漠区不同微生境下齿肋赤藓对一次降雪的生理生化响应. 植物生态学报, 2014, 38(9): 978-989. |
| [36] |
Zhao R M, Hui R, Wang Z R, Liu L C, Xie M, An L Z. Winter snowfall can have a positive effect on photosynthetic carbon fixation and biomass accumulation of biological soil crusts from the Gurbantunggut Desert, China. Ecological Research, 2016, 31(2): 251-262. DOI:10.1007/s11284-016-1335-1 |
| [37] |
司守霞, 李宜轩, 回嵘, 刘立超, 谢敏, 王艳莉. 降雪对荒漠生物土壤结皮光合生理特性的影响. 中国沙漠, 2018, 38(3): 560-567. |
| [38] |
Karsten U, Lembcke S, Schumann R. The effects of ultraviolet radiation on photosynthetic performance, growth and sunscreen compounds in aeroterrestrial biofilm algae isolated from building facades. Planta, 2007, 225(4): 991-1000. |
| [39] |
Krause G H, Weis E. Chlorophyll fluorescence and photosynthesis: the basics. Annual Review of Plant Physiology and Plant Molecular Biology, 1991, 42(1): 313-349. |
| [40] |
Hui R, Li X R, Zhao R M, Liu L C, Gao Y H, Wei Y P. UV-B radiation suppresses chlorophyll fluorescence, photosynthetic pigment and antioxidant systems of two key species in soil crusts from the Tengger Desert, China. Journal of Arid Environments, 2015, 113: 6-15. |
| [41] |
Maxwell K, Johnson G N. Chlorophyll fluorescence—a practical guide. Journal of Experimental Botany, 2000, 51(345): 659-668. |
| [42] |
Li X G, Duan W, Meng Q W, Zou Q, Zhao S J. The function of chloroplastic NAD(P)H dehydrogenase in tobacco during chilling stress under low irradiance. Plant and Cell Physiology, 2004, 45(1): 103-108. |
| [43] |
Druege U, Kadner R. Response of post-storage carbohydrate levels in pelargonium cuttings to reduced air temperature during rooting and the relationship with leaf senescence and adventitious root formation. Postharvest Biology and Technology, 2008, 47(1): 126-135. |
| [44] |
衣英华, 樊大勇, 谢宗强, 陈芳清. 模拟淹水对枫杨和栓皮栎气体交换、叶绿素荧光和水势的影响. 植物生态学报, 2006, 30(6): 960-968. |
| [45] |
Mielke M S, Schaffer B. Leaf gas exchange, chlorophyll fluorescence and pigment indexes of Eugenia uniflora L. in response to changes in light intensity and soil flooding. Tree Physiology, 2010, 30(1): 45-55. |
| [46] |
Gadallah M A A. Effects of proline and glycinebetaine on Vicia faba responses to salt stress. Biologia Plantarum, 1999, 42(2): 249-257. |
| [47] |
Hui R, Zhao R M, Song G, Li Y X, Zhao Y, Wang Y L. Effects of enhanced ultraviolet-B radiation, water deficit, and their combination on UV-absorbing compounds and osmotic adjustment substances in two different moss species. Environmental Science and Pollution Research, 2018, 25(15): 14953-14963. |
| [48] |
Guo Y W, Zhao Y G. Effects of storage temperature on the physiological characteristics and vegetative propagation of desiccation-tolerant mosses. Biogeosciences, 2018, 15(3): 797-808. |
| [49] |
李茜倩, 尹本丰, 张元明. 不同微生境下齿肋赤藓(Syntrichia caninervis)生理生化特性对不同季节的响应. 生态学报, 2018, 38(6): 2092-2100. |
| [50] |
李孝凯, 沙伟, 国春晖, 张梅娟. 低温胁迫对毛尖紫萼藓、东亚砂藓生理生化及光合特性的影响. 江苏农业科学, 2014, 42(10): 355-359. |
| [51] |
吴楠, 魏美丽, 张元明. 生物土壤结皮中刺叶赤藓质膜透性对脱水、复水过程的响应. 自然科学进展, 2009, 19(9): 942-951. |
| [52] |
陈璇, 李金耀, 马纪, 张富春. 低温胁迫对春小麦和冬小麦叶片游离脯氨酸含量变化的影响. 新疆农业科学, 2007, 44(5): 553-556. |
| [53] |
赵江涛, 李晓峰, 李航, 徐睿忞. 可溶性糖在高等植物代谢调节中的生理作用. 安徽农业科学, 2006, 34(24): 6423-6425, 6427. |
| [54] |
Jaleel C A, Manivannan P, Wahid A, Farooq M, Al-Juburi H J, Somasundaram R, Panneerselvam R. Drought stress in plants: a review on morphological characteristics and pigments composition. International Journal of Agriculture & Biology, 2009, 11(1): 100-105. |
| [55] |
Yin B F, Li J W, Zhang Q, Wu N, Zhang J, Rong X Y, Tao Y, Zang Y X, Li Y G, Zhou X B, Zhang Y M. Freeze-thaw cycles change the physiological sensitivity of Syntrichia caninervis to snow cover. Journal of Plant Physiology, 2021, 266: 153528. |
| [56] |
邵文鹏. 几种常绿阔叶植物抗寒性研究[D]. 泰安: 山东农业大学, 2009.
|
| [57] |
Hui R, Li X R, Chen C Y, Zhao X, Jia R L, Liu L C, Wei Y P. Responses of photosynthetic properties and chloroplast ultrastructure of Bryum argenteum from a desert biological soil crust to elevated ultraviolet-B radiation. Physiologia Plantarum, 2013, 147(4): 489-501. |
| [58] |
时丽冉, 刘志华. 干旱胁迫对苣荬菜抗氧化酶和渗透调节物质的影响. 草地学报, 2010, 18(5): 673-677. |
| [59] |
Bu C F, Wang C, Yang Y S, Zhang L, Bowker M A. Physiological responses of artificial moss biocrusts to dehydration-rehydration process and heat stress on the Loess Plateau, China. Journal of Arid Land, 2017, 9(3): 419-431. |