 2023, Vol. 43
2023, Vol. 43文章信息
- 刘明智, 蒋日进, 陈峰, 李德伟, 印瑞, 周永东, 梁静香
- LIU Mingzhi, JIANG Rijin, CHEN Feng, LI Dewei, YIN Rui, ZHOU Yongdong, LIANG Jingxiang
- 乐清湾贝类群落组成及其粒径谱结构特征
- Analysis on community structure of shellfish and its characteristics of biomass particle size spectrum in Yueqing Bay, China
- 生态学报. 2023, 43(2): 777-788
- Acta Ecologica Sinica. 2023, 43(2): 777-788
- http://dx.doi.org/10.5846/stxb202111163221
-
文章历史
- 收稿日期: 2021-11-16
- 网络出版日期: 2022-09-26

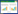




2. 浙江省海洋水产研究所, 农业农村部重点渔场渔业资源科学观测实验站, 浙江省海洋渔业资源可持续利用技术研究重点实验室, 舟山 316021
2. Key Laboratory of Sustainable Utilization of Technology Research for Fisheries Resources of Zhejiang Province, Scientific Observation and Experimental Station of Fishery Resources of Key Fishing Grounds, Ministry of Agriculture and Rural Affairs, Zhejiang Marine Fisheries Research Institute, Zhoushan 316021, China
乐清湾是浙江省内最大的半封闭式海湾[1], 由于其特殊的地理优势, 促使它成为滩涂贝类繁殖生长的优良场所[2]。自从20世纪80年代以来, 乐清湾网箱养殖、滩涂养殖等多种类型的养殖业迅速发展[3], 给临近水域和底栖环境带来巨大威胁, 造成严重污染[4—5], 导致乐清湾处于中潜在水平生态风险[6]。此外, 有报道显示野生海洋生物种群的大幅度减少与养殖逃逸有着密切的关系, 造成生物资源衰退, 物种多样性下降[7]。同样, 乐清湾也存在着养殖逃逸的现象, 综合上述原因导致乐清湾景观破碎化, 许多贝类群落结构改变, 产生衰退演替, 湾区生态系统脆弱性增加。贝类对水体中有机污染物、重金属具有一定的清除能力, 可以起到生态修复与生物调控作用[8—10], 此外贝类还具有生命周期较长、活动范围相对固定(稚贝以后)、生活习性相对稳定等特点, 其群落结构的改变能够表征群落的变化趋势与环境状况[11]。因此, 了解乐清湾贝类群落组成与结构变化显得十分必要。
目前, 有关乐清湾的研究主要集中在浮游生物和大型底栖动物生态位研究和季节变动[12—16], 另外一些是报道底栖生物群落组成以及时空变化等[17—20], 而滩涂贝类群落结构季节性的变化与生物量粒径谱特征却未见报道。为此, 作者在乐清湾滩涂开展海洋资源调查, 旨探明该湾区贝类资源的物种多样性, 丰富我国海湾地区软体动物地理区系的资料基础, 通过分析不同季节的贝类生物量粒径谱, 以期为乐清湾生态保护提供科学依据, 也为生态动力学研究提供基础。
1 材料与方法 1.1 数据来源本文数据取自2016年5月(春)、2016年8月(夏)、2016年11月(秋)和2017年2月(冬), 每个季度调查持续10天。共设置调查站位12个(S1—S12), 站位分布如图 1所示。在乐清湾滩涂选择具有代表性的、潮面底质类型相对均匀、潮带较为完整、无人为破坏或人为扰动较小且相对稳定的区域, 布设了12条断面, 各个断面底质均为泥质底。在调查海区, 选择不同生境的滩涂断面, 在大潮水退潮期间, 在每条断面又布设3站(即高潮带、中潮带和低潮带)。硬相(岩石岸)生物取样, 用25cm×25cm的定量框取2个样方; 软相(泥滩、泥沙滩、沙滩)生物取样, 用25cm×25cm×30cm的定量框随机取3个样方, 用筛网孔目为1.0mm进行筛洗底栖动物标本。同时, 在站位的周围广泛采集定性样品, 以补充定量样品的不足。样本用5%甲醛溶液固定, 带回实验室鉴定、计数、称重, 按照湿重: 干重=5:1换算为干重[21]。计算不同季节资源密度分布。生物种类尽可能鉴定到最小分类单元。生物种类名称及分类地位以《中国海洋生物名录》[22]、《浙江动物志》[23]为依据, 上述调查采样及测定均按《海洋调查规范第6部分:海洋生物调查(GB/T12763.6—2007)》[24]的有关标准进行。

|
| 图 1 乐清湾滩涂贝类调查站分布图 Fig. 1 Sampling station of shellfish in Yueqing Bay S1—S12, 第1至第12采样站位, site1—site12 |
采用Pinkas的相对重要性指数(Index of relative importance, IRI)[25]确定优势种与重要种。相对重要性指数的计算公式如下:
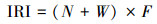
|
式中, N为某一种类的丰度占总丰度的百分比; W为某一种类的生物量占总生物量的百分比; F为某一种类出现的站数占调查总站数的百分比。根据不同贝类和螺类优势度(IRI)的大小, 确定不同种类在群落内的重要性。相对重要性指数(IRI)大于1000的种类定为优势种, 100—1000为重要种, 10—100之间的为常见种, 1—10为一般种, 小于1为稀有种。
1.2.2 群落多样性本文群落多样性分析采用以下公式计算[26]:
Shannon-wiener多样性指数:
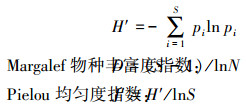
|
描绘ABC曲线, 分析乐清湾滩涂贝类群落受干扰情况。ABC曲线能较好的反应种群结构受污染扰动的变化:ABC曲线由生物量曲线与丰度曲线组成, 前者在上方时, 表明群落处于未受干扰的状态; 当两者相交, 表明处于中度干扰; 后者在上时, 表明群落处于严重干扰的状态。用W统计量(W-statistic)作为ABC曲线方法的一个统计量, 其公式为:
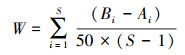
|
式中, Bi和Ai为ABC曲线中种类序号对应的生物量和丰度的累积百分比, S为出现物种数。当生物量优势曲线在丰度优势度曲线之上时, W为正, 反之W为负[27]。
1.2.4 贝类粒径谱的构建贝类生物量粒径谱以log2转换的个体干重生物量为横坐标划分粒级, 以log2转化的单位面积上的各粒径级的总干重生物量为纵坐标, 构建生物量粒径谱。标准化生物量粒径谱以log2转换的个体干重生物量为横坐标划分粒级, 在单位面积上每个粒径级的总干重生物量与该粒径级上个体干重的变化幅度的比值, 经过log2转化后为纵坐标。对标准化生物量粒径谱上的点作回归直线, 同时得到斜率a与截距b[28]。由于贝类生物个体大小差别较大, 丰度不同, 加之采样误差, 因此绘制标准化生物量粒径谱时, 应对数据进行筛选。数据分析与处理时, 群落多样性指数、资源密度及生物量粒径谱的绘制均使用EXCEL进行处理; ABC曲线及W统计量使用PRIMER 5.0软件进行绘图; 站位图、资源密度分布图利用surfer 13.0作图软件进行绘制。
2 结果 2.1 物种组成与优势种2016—2017年在乐清湾滩涂调查中, 根据4个季节的定性与定量样本分析, 共鉴定软体动物总计45种(表 1), 隶属于12目, 26科, 31属。以织纹螺科(Nassariidae)为主, 有7种; 其次为樱蛤科(Tellinidae)4种; 蚶科(Arcidae)、汇螺科(Potamodidae)和玉螺科(Naticidae)分别为3种; 骨螺科(Muricidae)、蓝蛤科(Corbulidae)、帘蛤科(Veneridae)、角贝科(Dentaliidae)和拟沼螺科(Assimineidae)为2种; 其余科均出现1种。从时间尺度分析, 夏季出现的生物种类最多, 为32种; 春季为30种; 秋季和冬季分别为27种和26种。从空间尺度可以看出, S12站位出现的种类最多, 为20种。S3和S7站位出现的种类最少, 分别为12种。
| 序号 Number |
目 Order |
科 Family |
种名 Species |
优势度IRI(Index of relative importance) | |||
| 春季Spring | 夏季Summer | 秋季Autumn | 冬季Winter | ||||
| 1 | 新腹足目Neogastropoda | 骨螺科Muricidae | 疣荔枝螺Thais clavigera | — | 0.09 | — | — |
| 2 | 爪哇荔枝螺Thais javanica | — | 0.06 | — | — | ||
| 3 | 织纹螺科Nassariidae | 半褶织纹螺Nassarius semiplicata | 41.58 | 170.07 | 833.81 | 2059.43 | |
| 4 | 群栖织纹螺Nassarius gregaria | 0.00 | — | — | 1.29 | ||
| 5 | 西格织纹螺Nassarius siquijorensis | 0.00 | 0.71 | — | 0.11 | ||
| 6 | 细肋织纹螺Zeuxis scalaris | — | 0.27 | 0.07 | — | ||
| 7 | 秀丽织纹螺Nassarius festiva | 0.33 | 0.44 | 1.52 | 3.06 | ||
| 8 | 秀长织纹螺Nassarius oveolatus | 2.44 | 1.00 | 337.87 | 29.82 | ||
| 9 | 纵肋织纹螺Nassarius variciferus | 0.17 | — | 1.39 | 0.24 | ||
| 10 | 蛾螺科Buccinidae | 褐管蛾螺Siphonalia spadicea | 0.08 | — | — | — | |
| 11 | 核螺科Columbellidae | 丽小笔螺Mitrella bella | — | — | 0.01 | — | |
| 12 | 海螂目Myoida | 蓝蛤科Corbulidae | 光滑河蓝蛤Potamocorbula laevis | — | 0.07 | — | — |
| 13 | 抱蛤科Corbulidae | 焦河蓝蛤Potamocorbula ustulata | 9655.55 | 5336.61 | 1722.07 | 6584.18 | |
| 14 | 蚶目Arcoida | 蚶科Arcidae | 毛蚶Scapharca kagoshimensis | — | — | 0.02 | — |
| 15 | 泥蚶Tegillarca granosa | 18.30 | 48.97 | 72.60 | 371.89 | ||
| 16 | 青蚶Barbatia obliquata | — | — | — | 0.25 | ||
| 17 | 胡桃蛤目Nuculoida | 吻状蛤科Nuculanidae | 薄云母蛤Yoldia similis | — | 0.03 | — | — |
| 18 | 帘蛤目Veneroida | 帘蛤科Veneridae | 薄壳镜蛤Dosinia corrugata | 0.64 | 0.09 | 0.15 | 2.97 |
| 19 | 青蛤Cyclina sinensis | 0.22 | 0.08 | 0.01 | 8.35 | ||
| 20 | 樱蛤科Tellinidae | 彩虹明樱蛤Moerella iridescens | 3933.32 | 52.09 | 181.95 | 1489.83 | |
| 21 | 刀明樱蛤Moerella culter | 0.04 | — | — | — | ||
| 22 | 虹光亮樱蛤Nitidotellina iridella | 0.04 | 0.31 | — | — | ||
| 23 | 江户明樱蛤Moerella jedoensis | 0.09 | — | — | 0.06 | ||
| 24 | 刀蛏科Cultellidae | 小刀蛏Cultellus attenuatus | — | 0.08 | — | 3.75 | |
| 25 | 灯塔蛏科Pharellidae | 缢蛏Sinonovacula constricia | 611.97 | 46.18 | 75.50 | 721.78 | |
| 26 | 绿螂科Glauconomidae | 中国绿螂Glauconome chinensis gray | — | 0.06 | — | — | |
| 27 | 列齿目Taxodonta | 蚶科Arcidae | 橄榄蚶Estellarca olivacea | 0.93 | 2.63 | 3.73 | 10.24 |
| 28 | 笋螂目Pholadomyoida | 鸭嘴蛤科laternulidae | 渤海鸭嘴蛤Laternula marilina | 0.29 | 8.05 | — | — |
| 29 | 头楯目Cephalaspidea | 阿地螺科Atyidae | 泥螺Bullacta exarata | 279.18 | 20.59 | 675.38 | 1497.70 |
| 30 | 囊罗科Retusidae | 婆罗囊螺Retusa boenensis | 888.16 | 972.72 | 1657.36 | 1096.77 | |
| 31 | 壳蛞蝓科Philinidae | 经氏壳蛞蝓Philine kinglipini Tchang | 0.28 | 0.02 | 0.95 | — | |
| 32 | 象牙贝目Dentalioida | 角贝科Dentaliidae | 钝角贝Dentalium obtusum | 0.13 | — | — | — |
| 33 | 肋娈角贝Dentalium octangulatum Donovan | 0.31 | — | 0.04 | 0.03 | ||
| 34 | 原始腹足目Archaeogastropoda | 马蹄螺科Trochidae | 丽口螺Calliostoma aculeatum | — | — | 0.01 | — |
| 35 | 异腹足目Heterogastropoda | 玉螺科Naticidae | 扁玉螺Glossaulax didyma | 148.42 | 2612.18 | 139.50 | 442.14 |
| 36 | 微黄镰玉螺Lunatia gilva | 665.81 | 2708.59 | 5423.76 | 360.90 | ||
| 37 | 斑玉螺Natica tigrina | — | — | 0.87 | 0.05 | ||
| 38 | 光螺科Eulimidae | 马氏光螺Melanella martinii | 0.02 | — | 0.22 | 0.03 | |
| 39 | 中腹足目Mesogastropoda | 滨螺科littorinidae | 粗糙滨螺Littorina scabra | 1.23 | — | — | 0.06 |
| 40 | 拟沼螺科Assimineidae | 堇拟沼螺Assiminea violacea | 0.01 | — | 0.04 | 1.93 | |
| 41 | 绯拟沼螺Assiminea latericea | 0.02 | 0.00 | — | — | ||
| 42 | 狭口螺科Stenothyridae | 光滑狭口螺Stenothyra glabra | 12.17 | 2497.79 | 946.94 | 676.92 | |
| 43 | 汇螺科Potamodidae | 红树拟蟹守螺Cerithidea rhizophorarum | — | 0.00 | — | — | |
| 44 | 尖锥拟蟹守螺Cerithidea largillierti | 0.31 | 0.27 | 6.97 | — | ||
| 45 | 珠带拟蟹守螺Cerithidea cingulata | 395.36 | 2258.16 | 1658.11 | — | ||
| “—”表示未采集到数据 | |||||||
由表 1可知, 焦河蓝蛤(Potamocorbula ustulata)是乐清湾第一优势种, 4个季节的平均优势度为5824.6, 因其较强的环境适应能力, 优势地位极其明显。春季的优势种主要是彩虹明樱蛤(Moerella iridescens)(3933.32);夏季分别为扁玉螺(Neverita didyma)(2612.18)、微黄镰玉螺(Lunatia gilva)(2708.59)、光滑狭口螺(Stenothyra glabra)(2497.79)和珠带拟蟹守螺(Cerithidea cingulata)(2258.16);秋季为婆罗囊螺(Retusa boenensis)(1657.36)、微黄镰玉螺(5423.76)和珠带拟蟹守螺(1658.11);冬季为半褶织纹螺(Nassarius sinarus)(2059.43)、彩虹明樱蛤(1489.83)、泥螺(Bullacta exarata)(1497.7)、婆罗囊螺(1096.77)。除了焦河蓝蛤之外, 其他优势种在不同季节基本上为重要种或常见种。
2.2 资源密度资源密度2016—2017年乐清湾滩涂调查总计12个站位, 其中春季贝类平均栖息密度为41.58×103个/m2, 平均生物量密度为1434.99g/m2(图 2、图 3)。从春季的栖息密度分布图看呈现南北多, 中部少的趋势, 生物量密度图分布沿岸趋势较为均匀; 夏季贝类平均栖息密度为58.65×103个/m2, 平均生物量密度为1939.78g/m2, 夏季的栖息密度分布图与春季相反, 呈现出中间多两边少的情况, 生物量密度分布趋势与春季基本相同, 但要大于春季的生物量密度; 秋季贝类平均栖息密度为14.98×103个/m2, 平均生物量密度为557.74g/m2, 秋季的栖息密度图表现最为平均, 只有S3站位栖息密度最高, 为153×103个/m2, 生物量密度图表现为两边多, 中间少; 冬季贝类平均栖息密度为13.18×103个/m2, 平均生物量密度为472.69g/m2, 栖息密度表现较为均匀, 只有S1站位的栖息密度较大, 生物量密度图呈现出由北向南逐渐减少的趋势。从2016—2017年调查的贝类的资源密度分布图看, 夏季的栖息密度与生物量密度最大, 其次为春季, 秋季和冬季的资源密度相当。

|
| 图 2 2016—2017年乐清湾贝类资源数量密度 Fig. 2 The distribution of shellfish resource density of individual in2016 and 2017 |

|
| 图 3 2016—2017年乐清湾贝类资源重量密度 Fig. 3 The distribution of shellfish resource density of weight in2016 and 2017 |
不同季节贝类的多样性指数结果显示(表 2), 丰富度指数(D)随季节的变化逐渐减小, 春季最高, 冬季最低; 多样性指数(H′)秋季最高, 夏季和冬季次之, 春季最低; 均匀度指数(J′)以秋季最高, 夏季和冬季相差无几, 春季最低。利用多样性指数在季节序列上进行Pearson相关性分析, 得出4个季节中只有均匀度指数与多样性指数显著相关(R=0.993, P<0.05), 其余指数之间在时间尺度上均不相关。
| 季节 Season |
丰富度指数 Richness index(D) |
多样性指数 Diversity index(H′) |
均匀度指数 Evenness index(J′) |
季节 Season |
丰富度指数 Richness index(D) |
多样性指数 Diversity index(H′) |
均匀度指数 Evenness index(J′) |
|
| 春季Spring | 3.77 | 1.14 | 0.30 | 秋季Autumn | 3.18 | 2.07 | 0.59 | |
| 夏季Summer | 3.37 | 1.68 | 0.45 | 冬季Winter | 2.73 | 1.54 | 0.44 |
不同滩涂断面的多样性指数显示(表 3), 四季的丰富度指数(D)平均值随着潮位的降低呈现递增的趋势, 在春季和夏季, 不同潮位间丰富度指数(D)变化较明显, 秋季和冬季随着潮位的降低, 丰富度指数(D)越来越大; 四季多样性指数(H′)的平均值在中潮带最高, 高潮带次之, 低潮带最低。春季多样性指数(H′)随着潮位升高, 多样性指数(H′)增加, 秋冬两季多样性指数(H′)均在中潮位最高, 夏季在低潮带多样性指数(H′)最高。四季的均匀度指数(J′)平均值变化复杂, 以中潮带的均匀度指数(J′)最高, 高潮带和低潮带紧随其后。春季, 随着潮位上升, 均匀度指数(J′)增加, 在夏季, 低潮带均匀度指数(J′)最高, 秋冬两季中潮带均匀度指数(J′)最大。
| 多样性指数 Diversity index |
潮带 Intertidal zone |
春 Spring |
夏 Summer |
秋 Autumn |
冬 Winter |
四季平均值 Seasonal mean |
| 高潮带 | 1.26±0.39 | 1.26±0.38 | 1.38±0.74 | 1.43±0.45 | 1.33 | |
| 丰富度指数D | 中潮带 | 1.27±0.46 | 1.38±0.58 | 1.40±0.50 | 1.59±0.29 | 1.41 |
| 低潮带 | 1.25±0.25 | 1.35±0.43 | 1.49±0.42 | 1.63±0.53 | 1.43 | |
| 高潮带 | 1.50±0.45 | 1.29±0.68 | 1.96±0.92 | 1.78±0.74 | 1.63 | |
| 多样性指数H′ | 中潮带 | 1.29±0.66 | 1.24±0.79 | 1.98±0.73 | 2.10±0.56 | 1.65 |
| 低潮带 | 1.11±0.61 | 1.42±0.43 | 1.90±0.85 | 1.96±0.63 | 1.60 | |
| 高潮带 | 0.50±0.21 | 0.38±0.18 | 0.58±0.24 | 0.50±0.18 | 0.49 | |
| 均匀度指数J′ | 中潮带 | 0.44±0.21 | 0.35±0.21 | 0.70±0.11 | 0.60±0.17 | 0.52 |
| 低潮带 | 0.34±0.13 | 0.43±0.10 | 0.55±0.23 | 0.55±0.16 | 0.47 |
4个季节的ABC曲线表明(图 4), 乐清湾贝类的生物量优势度曲线和丰度优势度曲线均相交, 对应的W值分别为-0.041、0.026、-0.017、-0.095, W值小于0或者微大于0, 说明乐清湾贝类的生物群落处于中度干扰状态。

|
| 图 4 乐清湾不同季节贝类ABC曲线 Fig. 4 ABC curve and W value of shellfish community in Yueqing Bay from different seasons ABC curve, 丰度生物量曲线abundance-biomass curve |
对乐清湾滩涂不同季节贝类绘制生物量粒径谱图形(图 5), 发现四种谱型形状均不相同。春季出现了3个波峰与波谷, 且波峰与波谷呈现的趋势相反。起初在0—2粒级出现最低波谷, 接着曲线直线上升, 在2—4出现了最高波峰; 夏季的谱型呈“W”形, 为双波谷, 单波峰型。波峰出现在4—6粒级; 秋季的双波谷在夏季的基础上分别移向两端, 且生物量与夏季对应的波谷相比减小; 冬季生物量粒径谱表现出“V”形, 波谷出现在2—4粒级。除波谷外其余粒级上生物量分布较为均匀。

|
| 图 5 乐清湾滩涂不同季节贝类生物量粒径谱 Fig. 5 Biomass size spectra of shellfish community in different seasons in Yueqing Bay |
对不同季节的贝类生物量进行分析, 描绘标准化生物量粒径谱图形并建立回归直线(图 6), 斜率变化范围在-1.5539—-0.7373之间, 斜率最大的季节为秋季, 最小为春季。只有秋季的斜率大于-1, 表明除了秋季外, 其余季节的贝类生物量随生物个体增大反而减小; 截距范围在16.673—21.597之间, 截距最小在秋季, 春季相反。结果表明春季的生产力最高, 秋季生产力最低。

|
| 图 6 乐清湾滩涂不同季节贝类标准化生物量粒径谱 Fig. 6 Normalized biomass spectra of shellfish community in different seasons in Yueqing Bay |
与1996的调查相比, 2016年的贝类由73种减少到45种, 一些野生经济种尚未出现[29—30]; 乐清湾的优势种在织纹螺科类、玉螺科、汇螺科等基础上增加樱蛤科、狭口螺科、蓝蛤科、囊螺科等, 优势群落逐渐发生转变, 说明贝类物种群落结构具有次生演替的趋势[28]。2016年乐清湾的贝螺种类按照夏、春、秋、冬呈现出逐渐递减的趋势。在不同季节, 水温对贝类的数量分布有一定的限制作用, 且与季节存在一定的的相关性[29—30]。从采样站位分析, 贝类的种类数量分布由北向南逐渐增多, 水温随着纬度降低而逐渐升高, 这一结论在前人的一些研究中也有体现[31—34]。由此可以得出贝类的分布与温度有着密切的关系。
本次贝类资源密度调查显示:四季的平均栖息密度和平均生物量密度分别为32.09个/m2和1103g/m2。调查站位的栖息密度分布特点随着季节的变化, 由两边向中间聚集, 最后呈均匀分布。春季气温开始回升, 加之S1站位与S12站位的海水交换量相对较少, 栖息环境较为稳定, 栖息密度在这两个站位相对较大。到了夏天, 正是人工养殖的黄金时期, 中部地区有许多养殖区域, 许多种类的外逃可能造成栖息密度向中间聚集的趋势。秋天与冬天的栖息密度较小, 这可能是与环境温度存在关联; 生物量密度的分布特点是夏季最大, 春季次之, 秋季较小, 冬季最小, 这也与前面的结论相同, 再次证明贝类的分布受温度影响较大[30]。此外, 资源密度分布和种类组成在不同季节变化趋势基本一致。
3.2 生物多样性指数多样性是表征群落水平和群落稳定性的重要生态特征, 与人类的生存与发展密切相关。以时间尺度结合贝类多样性指数的研究结果分析, 春夏的物种丰富度高于秋冬两季, 由此可以说明季节对贝类的生长有着一定的影响, 春夏水温逐渐升高, 贝类的生长速度加快, 生物丰富度指数较高, 优势种资源密度的表现更加明显。从空间尺度分析, 高潮带与中潮带的生物多样性要高于低潮带, 这主要是由于高潮带被海水冲刷的较少, 生境较为稳定, 为贝类栖息创造了良好的环境。中潮带的生境分化程度较低, 植被较为稀疏, 环境也较为稳定, 因此高中潮带的生物多样性较高, 生物种类组成复杂, 但冬夏两季的多样性指数低潮带要高于高潮带, 这也说明潮带的变化不是唯一影响贝类栖息的因素[35]。结合时序与潮间带的结果显示, 乐清湾贝类的均匀度始终处于较低水平, 优势种始终由几种贝类控制, 导致均匀分布程度较低, 同时也表明乐清湾的贝类结构组成较为单一。
3.3 群落结构的稳定性分析结合ABC曲线特征及W统计值, 根据Clarke和Warwick的划分标准, 结果显示:在4个季节中, 丰度曲线与生物量曲线先分离后相交, 原来的优势种减少, 群落向随机化、小型化种类方向演替, 导致丰度曲线与生物量曲线优势度很难辨别, 说明调查海域贝类群落处于中度扰动状态。究其原因主要有两方面, 一是生境的不稳定性, 湾区贝类生境常常遭到破坏。近几十年来, 乐清湾沿岸的经济迅速发展, 海岸工程的大量兴起, 大量工厂建筑拔地而起, 在施工及运作过程中, 给乐清湾滩涂带来巨大影响, 导致生境破碎, 造成生物多样性下降; 二是水质条件的恶化。根据2016—2017年中国海洋生态环境状况公报显示乐清湾海域为劣四类海水, 主要污染物为无机氮和活性磷酸盐。水质条件差的原因主要归结为围塘养殖与污水的排放。为了满足人们对于海产品的需求, 围塘养殖模型风靡于此, 在此过程中农药的大量使用给周围滩涂生境带来了“灾难性”影响, 加之沿岸的工农污水、生活污水等排放严重影响滩涂水域环境; 抛开上述因素外, 养殖逃逸对野生种群的冲击也很大。养殖品种逃逸后, 侵占土著种栖息环境, 产生食物竞争, 逐渐导致原有种群结构出现衰退。多种因素的耦合作用才导致了乐清湾生物群落结构稳定性差、生物量与栖息密度低, 生态系统处于中风险。因此应该以生态本底和自然禀赋为基础, 坚持人与自然和谐相处, 实现生物多样性保护和经济高质量发展相匹配。
3.4 乐清湾贝类生物量粒径谱与标准化粒径谱特征分析通过分析不同季节生物量粒径谱可知, 大型底栖生物与粒径谱往往较为复杂, 表现出一定的不规则性。焦河蓝蛤等相对重要性指数较高的底栖生物通常位于不同粒级的波峰区, 这表明绝对优势种始终控制着生物量粒径谱的峰值。波谷通常由一些处于幼体状态的底栖动物组成, 这些幼体通常具有生长速度快、死亡率高的特性, 所以生物量处于较低水平[35]。因此可以推测出粒径级的改变可能与生物体生长速率存在某些关联。
标准化粒径谱的斜率、截距、最小粒级的生物量往往可以揭示不同季节潮间带生态系统的特征。斜率可以描述生态系统中生物量的变化趋势:当斜率为-1时, 作为生物量与个体大小变化之间的一个平衡点, 二者不发生变化; 当斜率大于-1时, 生物量随着生物个体的增大而增大; 当斜率小于-1时, 则相反。乐清湾不同季节的贝类标准化生物量粒径谱的斜率在春、夏、冬均小于-1, 说明生物量随着个体的增大而减小。乐清湾贝类标准化生物量粒径谱斜率小于国内大多数海域[36—39], 这可能与标准化粒径谱选取的生物种类有关。由于本次只研究贝类, 而其他学者多数是研究较为完整的底栖生物, 包括一些线虫和小型底栖生物等, 这可能是导致斜率较小的原因之一。同时, 对比斜率还可以得出乐清湾相较其他海域, 其营养循环处于较低水平。此外斜率还与ABC曲线的W值有着一定的关联, 两者通常呈正相关[40]。标准化生物量粒径谱的截距与最小粒级生物量(BMS)可以反映生态系统中生产力水平的高低。对乐清湾不同季节的标准化生物量粒径谱截距、BMS进行比较可以发现, 春季的截距、BMS高于其他季节, 说明春季的生产力水平较高。春季乐清湾出现大量焦河蓝蛤, 其为河口常见种, 有着较强的适应能力, 数量较多, 其IRI指数达到了9655.55, 导致了次级生产力升高, 这与瓯江口、鼓浪屿海域生产力水平较高的原因相似[41—42]。
4 结论本次调查发现乐清湾贝类物种数量与20年前相比减少了28种, 优势种较为单一, 如焦河蓝蛤等。生物量粒径谱峰值区域始终由优势种控制。湾区营养循环水平在我国处于较低水平, 但贝类生产力水平较高。
生物量粒径谱作为一种生态学研究方法, 可以比较不同生态系统的生产力水平, 描述生物量的变化趋势, 并且对环境的变化非常敏感, 为认识不同层次的生物规律和环境监测提供了新的途径。但是由于采样范围不足和粒级单位不统一等原因, 造成结果可能不会反应出完整的真实情况, 这也导致了生物量粒径谱发展缓慢。随着人们对海洋生态系统结构和功能认识更加深入, 相信粒径谱理论的应用将更加广泛[43—44]。
| [1] |
周航. 浙江海岛志. 北京: 高等教育出版社, 1998.
|
| [2] |
吴奋武, 李熹. 浙江省温州市滩涂贝苗增养殖情况介绍. 福建水产, 1992(1): 77-79. DOI:10.14012/j.cnki.fjsc.1992.01.020 |
| [3] |
王姮, 郭远明, 胡红美, 应忠真, 李子孟, 鲍静姣. 2016年乐清湾北部海域富营养化现状与评价. 现代农业科技, 2019(17): 176-178. DOI:10.3969/j.issn.1007-5739.2019.17.106 |
| [4] |
吕晓婷, 温艳萍. 江厦潮汐电站库区滩涂水产养殖现状及对策. 中国农学通报, 2013, 29(20): 69-72. DOI:10.11924/j.issn.1000-6850.2012-3073 |
| [5] |
杨辉. 浙江乐清湾资源优势及其开发中的矛盾处理. 海洋通报, 2003(6): 49-55. DOI:10.3969/j.issn.1001-6392.2003.06.008 |
| [6] |
陈思杨, 宋琍琍, 刘希真, 刘瑞娟, 余骏. 浙江典型海湾潮间带沉积物污染及生态风险评价. 中国环境科学, 2020, 40(4): 1771-1781. DOI:10.3969/j.issn.1000-6923.2020.04.045 |
| [7] |
Yeates S E, Einum S, Fleming I A, Holt W V, Gage M J G. Assessing risks of invasion through gamete performance: farm Atlantic salmon sperm and eggs show equivalence in function, fertility, compatibility and competitiveness to wild Atlantic salmon. Evolutionary Applications, 2014, 7(4): 493-505. DOI:10.1111/eva.12148 |
| [8] |
Owen G. Feeding and Digestion in the Bivalvia. Advances in Comparative Physiology & Biochemistry, 1974(5): 1-35. |
| [9] |
Smaal A C, Vonck W, Prins T C. The functional role of mussels in the osterschelde estuary. J Shellfish Res, 1989, 8(2): 472-478. |
| [10] |
Reeders H H, Vaate A B D. Zebra Mussels (Dreissena polymorpha): A New Perspective for Water Quality Management. Hydrobiologia, 1990, 200-201(1): 437-450. DOI:10.1007/BF02530361 |
| [11] |
赵永强, 曾江宁, 高爱根, 陈全震, 廖一波, 寿鹿. 椒江口滩涂大型底栖动物群落格局与多样性. 生物多样性, 2009, 17(3): 303-309. |
| [12] |
徐晓群, 曾江宁, 陈全震, 刘晶晶, 杜萍, 王桂忠. 浙江三门湾浮游动物优势种空间生态位. 应用生态学报, 2013, 24(3): 818-824. DOI:10.13287/j.1001-9332.2013.0238 |
| [13] |
董静瑞, 水柏年, 胡成业, 水玉跃, 杜肖, 田阔. 温州南部沿岸海域主要鱼类的生态位及种间联结性. 应用生态学报, 2017, 28(5): 1699-1706. DOI:10.13287/j.1001-9332.201705.028 |
| [14] |
刘镇盛, 王春生, 张志南, 蔡昱明, 张东声. 乐清湾浮游动物的季节变动及摄食率. 生态学报, 2005(8): 1853-1862. DOI:10.3321/j.issn:1000-0933.2005.08.004 |
| [15] |
宋琍琍, 赵聪蛟, 龙华, 陈雷, 余骏, 吕宝强, 刘亚林. 2009年夏季乐清湾网采浮游植物群落结构特征. 海洋学研究, 2010, 28(3): 34-42. DOI:10.3969/j.issn.1001-909X.2010.03.005 |
| [16] |
彭欣, 谢起浪, 陈少波, 黄晓林, 仇建标, 仲伟, 关万春. 乐清湾潮间带大型底栖动物群落分布格局及其对人类活动的响应. 生态学报, 2011, 31(4): 954-963. |
| [17] |
高爱根, 陈全震, 曾江宁, 廖一波, 杨俊毅. 西门岛红树林区大型底栖动物的群落结构. 海洋学研究, 2005(2): 33-40. DOI:10.3969/j.issn.1001-909X.2005.02.005 |
| [18] |
郑荣泉, 张永普, 李灿阳, 周芸. 乐清湾滩涂大型底栖动物群落结构的时空变化. 动物学报, 2007(2): 390-398. |
| [19] |
龙华, 余骏, 周燕. 大型底栖动物污染指数在乐清湾潮间带环境质量评价中的应用. 海洋学研究, 2008, 26(4): 97-104. |
| [20] |
彭欣, 谢起浪, 陈少波, 黄晓林, 仇建标, 仲伟, 陈万东. 南麂列岛潮间带底栖生物时空分布及其对人类活动的响应. 海洋与湖沼, 2009, 40(5): 584-589. DOI:10.3321/j.issn:0029-814X.2009.05.010 |
| [21] |
林岿璇, 张志南, 王睿照. 东、黄海典型站位底栖动物粒径谱研究. 生态学报, 2004(2): 241-245. |
| [22] |
刘瑞玉. 中国海洋生物名录. 北京: 科学出版社, 2008.
|
| [23] |
董聿茂. 浙江动物志两栖类爬行类. 杭州: 浙江科学技术出版社, 1990.
|
| [24] |
中华人民共和国国家质量监督检验检疫局. 海洋调查规范第6部分海洋生物调查. 北京: 中国标准出版社, 2008: 56-62.
|
| [25] |
Pinkas L, Oliphant M, Iverson I L K. Food habits of albacore, bluefin tuna, and bonito in California waters. Water Research, 1971, 18(6): 653-594. |
| [26] |
Margalef R. Information theory in ecology. General Systematics Advancing Frontiers Plant Science, 1957, 3: 37-71. |
| [27] |
Clarke K R, Warwick R M. Change in Marine Communities: an Approach to Statistical Analysis and Interpretation (2nd ed). Plymouth: PRIMPER-E. 2001, 1-91.
|
| [28] |
Sprules W G, Munawar M. Plankton size spectra in relation to ecosystem productivity, size, and perturbation. Canadian Journal of Fisheries and Aquatic Sciences, 1986, 43(9): 1789-1794. |
| [29] |
张永普, 郑汉淼, 叶文俊, 余存春. 乐清湾潮间带贝类的区系研究. 温州师范学院学报: 自然科学版, 1996(3): 75-79. |
| [30] |
贾胜华, 曾江宁, 廖一波, 寿鹿, 黄伟, 高爱根, 汤雁滨. 洞头列岛及邻近海域大型底栖动物群落结构的研究. 海洋学研究, 2016, 34(2): 83-92. |
| [31] |
施华宏. 滨海采捕对岸带生物资源的影响及管理对策. 海洋开发与管理, 2001(2): 32-34. |
| [32] |
李冠国, 范振刚. 海洋生态学. 北京: 高等教育出版社, 2004: 85-325.
|
| [33] |
安传光, 赵云龙, 林凌, 吕敢堂, 陈亚瞿. 长江口九段沙潮间带大型底栖动物季节分布特征的初步研究. 水产学报, 2007(S1): 52-58. |
| [34] |
卢占晖, 朱文斌, 徐开达, 周永东, 戴乾, 卢衎尔. 浙江沿岸海域春季螺类群落结构特征及其与环境因子的关系. 水生生物学报, 2018, 42(3): 606-615. |
| [35] |
Strayer D. The size structure of a lacustrine zoobenthic community. Oecologia, 1986, 69(4): 513-516. |
| [36] |
张青田, 胡桂坤. 渤海湾近岸底栖动物的生物量粒径谱研究. 天津师范大学学报: 自然科学版, 2013, 33(2): 81-88. |
| [37] |
华尔, 张志南. 黄河口邻近海域底栖动物粒径谱研究. 中国海洋大学学报: 自然科学版, 2009, 39(5): 971-978. |
| [38] |
林岿璇, 张志南, 王睿照. 东、黄海典型站位底栖动物粒径谱研究. 生态学报, 2004(2): 241-245. |
| [39] |
邓可. 南黄海两种不同生境底栖动物粒径谱研究[D]. 青岛: 中国海洋大学, 2006.
|
| [40] |
Pagola-Carte S. ABC Method and Biomass Size Spectra: What About Macrozoobenthic Biomass on Hard Substrata?. Hydrobiologia, 2004, 527(1): 163-176. |
| [41] |
周细平, 蔡立哲, 梁俊彦, 傅素晶, 林和山, 黄昆. 厦门海域大型底栖动物次级生产力的初步研究. 厦门大学学报: 自然科学版, 2008(6): 902-906. |
| [42] |
贾胜华, 廖一波, 曾江宁, 寿鹿, 黄伟, 高爱根, 汤雁滨. 近岸典型生态系统大型底栖动物的次级生产力—以瓯江口、洞头列岛为例. 生态学报, 2017, 37(21): 7140-7151. |
| [43] |
周林滨, 谭烨辉, 黄良民, 连喜平. 水生生物粒径谱/生物量谱研究进展. 生态学报, 2010, 30(12): 3319-3333. |
| [44] |
王睿照, 张志南. 海洋底栖生物粒径谱的研究. 海洋湖沼通报, 2003(4): 61-68. |