 2023, Vol. 43
2023, Vol. 43文章信息
- 李加龙, 罗纯良, 吕恒, 许佳峰, 罗潋葱, 潘珉, 何锋, 满小明, 张如枫, 龚发露, 孙婷, 陈岚, 武桂竹
- LI Jialong, LUO Chunliang, LÜ Heng, XU Jiafeng, LUO Liancong, PAN Min, HE Feng, MAN Xiaoming, ZHANG Rufeng, GONG Falu, SUN Ting, CHEN Lan, WU Guizhu
- 2002-2018年滇池外海蓝藻水华暴发时空变化特征及其驱动因子
- Spatio-temporal variation and driving factors of algal bloom at Lake Dianchi during 2002-2018
- 生态学报. 2023, 43(2): 878-891
- Acta Ecologica Sinica. 2023, 43(2): 878-891
- http://dx.doi.org/10.5846/stxb202110253002
-
文章历史
- 收稿日期: 2021-10-25
- 网络出版日期: 2022-09-26





2. 昆明市滇池高原湖泊研究院, 昆明 650500;
3. 南京师范大学虚拟地理环境教育部重点实验室, 南京 210023;
4. 普天信息技术有限公司, 北京 100085
2. Kunming Dianchi and Plateau Lake Research Institute, Kunming 650500, China;
3. Key Laboratory of Virtual Geographic Environment of Education Ministry, Nanjing Normal University, Nanjing 210023, China;
4. Potevio Information Technology Co., Ltd., Beijing 100085, China
湖泊富营养化引起的蓝藻水华是全球最严重的环境和生态问题之一[1—2], 对人类健康有着重大影响[3—5]。根据2020年中国生态环境状况公报显示, 我国的110个重要湖泊(水库)中, 中营养状态占61.8%, 富营养状态占29%[6]。国家重点整治的“三湖”[7]中, 太湖和巢湖目前处于轻度富营养状态, 滇池为中度富营养状态, 蓝藻水华频发, 湖泊环境管理和治理面临巨大压力。
自20世纪80年代以来, 因周边城镇与农业的发展, 滇池流域营养负荷显著增加, 导致富营养化程度加剧[8—10]; 至90年代, 滇池成为国家“三湖三河”重点治理对象之一。对此当地政府实施了环湖截污、外流域引水、入湖河道整治、面源治理、生态修复、底泥疏浚[11—12]“六大工程”进行污染治理, 滇池水质稍有改善, 但富营养化问题仍未解决, 每年蓝藻水华依然严重, 带来了一系列的生态环境影响[13]。
近年来, 对滇池蓝藻水华的关注逐渐增多[8, 14—17], 水华的监测手段也逐渐丰富多样。相比传统的采样分析叶绿素a(Chl-a)浓度甚至通过浮标探头进行Chl-a定点自动监测, 卫星遥感监测手段具有大面积同时观测、时效性强、动态连续性强的特点, 已被广泛运用于蓝藻水华监测与识别研究中, 成为水环境动态监测不可或缺的有效技术手段之一[18—20]。相比于太湖和巢湖, 滇池蓝藻水华的遥感监测应用较少[21—22], 多为短期监测[23—24], 且分辨率较低[25]。在卫星遥感监测中, 目前常用的方法为单波段法、比值法、归一化植被指数(Normalized Difference Vegetation Index, NDVI)和增强植被指数(Enchanted Vegetation Index, EVI)非线性识别方法[26—27]。近期研究表明, 由中分辨率成像光谱辐射计(Moderate Resolution Imaging Spectroradiometer, MODIS)测量得到的浮藻指数法(Floating Algae Index, FAI)可提供长期且实时的表层蓝藻水华历史信息, 可准确识别水华覆盖度和水华的时间动态变化[28]。FAI法在不同的环境和观测条件下, 不易受气溶胶厚度和观测角度的影响[28], 已广泛应用于蓝藻水华监测[19, 29]。通过在太湖和巢湖的应用, 已证明该方法在实时监测和历史分析中较为客观和准确[30]。
以往有关滇池蓝藻水华的研究, 多以单点Chl-a浓度作为蓝藻水华的评价因子, 但其具有时间和空间上的不连续性[14, 31—32], 且多以整个滇池(含草海和外海)或滇池外海作为一个整体区域进行研究。滇池外海, 作为滇池的主体水域, 受气象和水文条件的影响, 其水质和水华发生规律均具有较大的空间差异性[33]。因此, 有必要对外海不同分区的水华发生特征进行详细研究。本文结合最新的滇池蓝藻水华程度级别界定标准, 结合外海水质监测点位布设规律, 将外海划分成北、中和南部3个区域(图 1), 利用FAI法对上述三区域在2002—2018年间的水华信息进行提取, 探讨该时段内外海水华的时空分布特征。通过冗余分析(Redundancy Analysis, RDA)和随机森林(Random Forest, RF)方法, 分析了各区域环境因子(水质和气象因子)对响应因子(平均水华面积和Chl-a)的影响。此外, 利用三维水动力生态模型(Aquatic Ecosystem Model—3 Dimensional, AEM3D)计算了外海不同区域的水力滞留时间(Water Residence Time, WRT), 通过斯皮尔曼相关分析法分析了水华面积与水力滞留时间的关系, 从水动力学角度, 补充了滇池外海水华暴发影响因素的研究, 为进一步有效地对滇池蓝藻水华监测、预测预警和有效防控提供科学依据。

|
| 图 1 滇池地理位置、主要河道和水华分区 Fig. 1 Location, main tributaries and zones for algal bloom of Lake Dianchi |
滇池(24°28′—25°28′ N, 102°30′—103°00′ E)位于云南省昆明市南部, 是云南省最大的淡水湖, 隶属于长江流域金沙江水系[34—35]。当滇池湖面高程为1887.4 m时, 其湖面面积298.06 km2, 湖容量14.94×108 m3, 湖周长161.55 km, 最大水深11.35 m, 平均水深5.01 m; 湖泊呈南北长条形, 南北最大长度为39.4 km, 东西最大宽度为12.66 km[36]。自1996年海埂大坝建成以后, 滇池被其分为南、北两部分, 其中北部草海面积为10.8 km2, 占滇池总面积的3.6%, 平均水深为2.5 m, 南部外海面积为289.2 km2, 占滇池总面积的96.4%, 平均水深为4.4 m[37]。
滇池流域地处低纬度、高海拔地区, 属亚热带高原季风气候, 昼夜温差大, 干湿季分明[38]。根据2002—2018年气象观测统计数据, 滇池流域年均气温约为16.1℃, 多年平均日照时数约为2177 h, 年均降雨量约为972.24 mm, 其中汛期(5—10月)降水量占全年总降水量的89%, 流域风向以西南风为主, 南风为辅, 年均风速为2.59 m/s。滇池流域水系(实为河渠)众多, 其中外海区域主要24条入湖河流分别为采莲河、金家河、盘龙江、大清河、海河、六甲宝象河、小清河、五甲宝象河、虾坝河、老宝象河、新宝象河、马料河、洛龙河、捞鱼河、南冲河、淤泥河、柴河、白鱼河、茨巷河、东大河、中河、古城河、广谱大沟及姚安河, 唯一出口位于西岸的海口河, 当滇池处于高水位时, 湖水由海口流入螳螂川, 最终向北汇入金沙江(图 1)[36]。
2 数据与分析 2.1 数据来源收集了滇池流域2002—2018年详细的气象、水质、卫星遥感数据, 以及2007—2018年外海24条主要入湖河流逐日流量、海口闸逐日出湖流量、中滩站逐日水位数据(表 1), 用于分析滇池蓝藻水华暴发时空变化特征及其驱动因子。气象数据均来自中国气象数据网(昆明市气象监测站, 台站号为56778), 包括逐时气温(AT)、降水(PP)、风速(WS)、风向(WD)、日照时长(SD)、相对湿度(RH)数据, 以及逐日平均气温、降水(当日20:00—次日20:00总降水量)、日最大风速和日主导风向等数据。此外还收集到滇池外海区域八个国控水质监测站点的逐月水质和水文数据, 包括总氮(TN)、总磷(TP)、pH、溶解氧(DO)、高锰酸盐指数(CODMn)、Chl-a和水温(WT)。获得滇池877幅遥感图像, 反演得到水华发生期间(精确到小时)不同区域的蓝藻水华面积(Algal Bloom Area, ABA)。
| 类别 Category |
指标 Indicator |
缩写 Abbreviation |
单位 Unit |
数据时段 Data period |
数据频率 Data frequency |
数据来源 Data source |
| 气象Meteorology | 气温 | AT | ℃ | 2002-2018 | 逐时、逐日 | 中国气象数据网 |
| 降水 | PP | mm | ||||
| 风速 | WS | m/s | ||||
| 风向 | WD | - | ||||
| 日照时长 | SD | h | ||||
| 相对湿度 | RH | % | ||||
| 水温 | WT | ℃ | ||||
| 水质Water quality | 总氮 | TN | mg/L | 2002—2018 | 逐月 | 中国环境监测总站 |
| 总磷 | TP | mg/L | ||||
| pH | pH | - | ||||
| 溶解氧 | DO | mg/L | ||||
| 高锰酸盐指数 | CODMn | mg/L | ||||
| 叶绿素a | Chl-a | μg/L | ||||
| 水文Hydrology | 流量 | - | m3/s | 2007—2018 | 逐日 | 昆明市水文水资源局 |
| 水位 | - | m | ||||
| 卫星遥感 Satellite remote sensing |
蓝藻水华面积 | ABA | km2 | 2002—2018 | - | NASA (https://oceancolor.gsfc.nasa.gov) |
遥感数据从NASA OceanColor数据网站(https://oceancolor.gsfc.nasa.gov)获取, 2002—2018年间滇池流域天气状况良好的Aqua/MODIS卫星遥感影像共877张, 基本涵盖各月份。影像数据使用SeaDAS(version 7.5)软件处理, 得到的去瑞利散射校正后的反射率(Rayleigh—corrected reflectance, Rrc)[39—41], 并采用墨卡托投影(Universal Transverse Mercator, UTM)进行地理参考。
2.2.2 蓝藻水华提取算法本文采用Hu等[42—43]提出一种快速、简单的蓝藻识别指标算法(FAI法)对滇池水华进行提取, 其计算公式为:
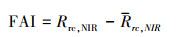
|
(1) |

|
(2) |
对于MODIS影像, RED波段为645 nm, NIR波段为859 nm, SWIR波段为1240 nm, 公式中反射率均为去瑞利散射的反射率。FAI是以645 nm和1240 nm为基线量取859 nm处的高度值。FAI的大小可以代表表层水体中藻类浓度的高低, 进而可以用于定义水华是否发生。正常水体在近红外波段呈现下降的趋势, 而水华的形成导致水色的改变, 使得水华水体在近红外波段的光谱曲线抬升, FAI的值代表光谱曲线抬起的高度。因此, 本研究根据FAI值来判断水华, 是否发生, 当FAI大于等于0时, 认为有水华发生; 当FAI的值小于0时, 认为水华未发生。以此为依据来确定滇池在像元尺度上是否发生水华。
2.2.3 AEM3D水动力生态模型使用Hydronumerics开发的三维水动力生态模型AEM3D(http://www.hydronumerics.com.au/software/aquatic-ecosystem-model-3d)对滇池外海不同区域的水力滞留时间进行计算。AEM3D模型由水动力模块和水生态模块组成[44], 水动力模块可模拟湖泊物理状况, 如波浪和不同深度的水温、水流和水下光合有效辐射; 水生态模块可模拟生态系统中的诸多因子, 例如碳—氮—磷循环、浮游植物、浮游动物、鱼类、水草、底栖生物的生命过程, 其具体配置简述如下。
(1) 模型配置
1) 网格划分和地形
整个滇池外海水面划分成35×21网格系统, X和Y方向的格距均为1000 m, 各格点水深如图 2。

|
| 图 2 AEM3D中使用的外海网格系统和各格点水深 Fig. 2 Grid system and water depth of all the grid cells at Waihai used for AEM3D AEM3D: 三维水动力生态模型Aquatic Ecosystem Model—3 Dimensional |
2) 气象与出入流边界
模型中需输入的7个气象参数分别为:水汽压(hPa)、气温(℃)、相对湿度(0—1)、降水量(m)、风速(m/s)、风向(°)和太阳长波辐射(W/m2)。入流文件由外海24条主要入湖河流的逐日流量(m3/s)、水温(℃)及主要水质数据组成。因模型中不考虑水质变化, 故所有入湖河流水质浓度均设为0。出流文件由海口闸逐日出湖水量(m3/s)和水温(℃)构成。
3) 时间步长
模拟时段为2007年1月1日—2018年12月31日, 计算时间步长为3600 s, 输出时间步长为24 h。
(2) 模型参数率定
模拟的2007—2018年逐日水位变化见图 3, 与实测水位间相关系数(r)高达0.99, 均方根误差(RMSE)为0.0072 m, 说明模型率定结果很好, 其主要物理参数如表 2所示。

|
| 图 3 2007—2018年逐日水位模拟与实测对比 Fig. 3 Comparison of simulated and measured daily water levels during 2007—2018 |
| 参数 Parameter |
单位 Unit |
参数取值 Value |
参数 Parameter |
单位 Unit |
参数取值 Value |
|
| 水体平均反射率Mean albedo of water | — | 0.08 | 临界风速Critical wind speed | m/s | 3.00 | |
| 水面辐射率Emissivity of water surface | — | 0.96 | 风力扰动效率Wind stirring efficiency | — | 0.2 |
本文通过R(Version 4.1.0;http://www.r-project.org/)软件对月时间尺度上滇池外海不同区域环境因子(气象和水质)与响应因子(ABA和Chl-a浓度)间关系进行统计分析。首先对数据进行除趋势对应分析(Detrended Correspondence Analysis, DCA), 结果表明所有排序轴梯度最大值小于3, 故选择冗余分析[45](RDA)作为分析方法。在RDA分析过程中, 所有环境因子数据全部进行标准化处理, 且采用方差膨胀因子(Variance Inflation Factor, VIF)度量环境因子贡献性程度, 结果表明所有环境因子VIF值均小于10, 无共线性问题。基于随机森林法[46](RF)评估单个环境因子的重要性, 通过“randomForest”包计算环境因子相对重要性得分, 通过得分排名从中确定环境因子对响应变量的重要程度。在RF分析过程中, 使用“A3”包对全模型显著性进行评估。通过SPSS(Version 25.0)软件对水华面积与水力滞留时间进行斯皮尔曼相关分析。
本文所有数据由Excel 2016软件处理, 所有图形绘制由Origin 2017软件完成。
3 结果与分析 3.1 滇池外海蓝藻水华年际变化特征研究时段内, 2002年滇池外海水华暴发始于4月, 以后各年均提前至每年1月(图 4)。从最大水华面积暴发月份判断, 春季(3—5月)占17.65%、夏季(6—8月)占17.65%、秋季(9—11月)占47.05%、冬季(12月和1—2月)占17.65%, 最大水华面积主要发生在秋季。2002—2018年, 年平均水华面积比例呈波动下降趋势, 多年平均水华面积比例为3.13%, 最大水华面积出现在2013年(6%), 最小出现在2005年(1.8%)。

|
| 图 4 2002—2018年外海水华统计特征 Fig. 4 Statistical characteristics of algal bloom at Waihai during 2002—2018 |
参考2021年3月1日昆明市市场监督管理局发布的《滇池蓝藻水华分级预警及应急处理》中水华程度分级标准(表 3), 统计了2002—2018年间滇池外海不同3个区域1—12月的平均水华面积比(图 5)。可以看出, 外海北部、中部和南部蓝藻水华面积在月尺度上均呈单峰型变化特征, 即夏季>秋季>冬季>春季。北部和中部区域水华面积比最大值出现在7月, 分别为33.3%(中度, 表 3)和25.2%(轻度), 南部区域最大值出现在8月, 为14.1%(轻度); 3个区域最小值均出现在2月, 分别为1.1%, 0.6%和1.7%(无明显水华); 年内平均水华面积比空间分布为外海北部(9.26%)>中部(6.17%)>南部(5.46%)。从东西方位水华面积比来看, 东北部(12.5%)>西北部(5.6%), 中西部(7.6%)>中东部(4.5%), 西南部(6.6%)>东南部(4.2%), 外海东部(东北、中东和东南)年内平均水华面积比(7.1%)略大于外海西部(西北、中西和西南部)。
| 水华程度级别Algal bloom level | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ |
| 水华面积比例P ABA proportion/% | 0 | 0 < P < 10 | 10≤P < 30 | 30≤P < 60 | 60≤P < 100 |
| 水华特征Characteristics of algal bloom | 无水华 | 无明显水华 | 轻度水华 | 中度水华 | 重度水华 |
| ABA:蓝藻水华面积Algal bloom area | |||||

|
| 图 5 外海不同区域多年月平均水华面积比例变化 Fig. 5 Monthly algal bloom area proportion at Waihai |
综上所述, 在滇池外海, 由北向南水华严重程度逐渐减轻; 在外海北部, 东部水华较西部更为严重; 而在外海中部和南部呈现与北部相反的趋势, 西部水华则较东部略为严重。
3.3 蓝藻水华影响因子分析 3.3.1 水华驱动因子为确定驱动蓝藻水华发生的影响因子, 在月尺度上对水质、气象和水华数据进行了分区(整个外海、外海北部、外海中部和外海南部)统计。通过冗余分析(RDA)法, 分别分析了上述四区域中水质因子(WT、TN、TP、DO、pH、CODMn)、气象因子(AT、PP、WS、SD、RH)与响应因子(ABA、Chl-a)间的关系, 结果见图 6。

|
| 图 6 环境因子与水华面积、叶绿素a的冗余分析 Fig. 6 Redundancy analysis of algal bloom, Chl-a and environmental factors AT:气温Air temperature; PP:降水Precipitation; WS:风速Wind speed; SD:日照时长Sunshine duration; RH:相对湿度Relative humidity; TN:总氮Total nitrogen; TP:总磷Total phosphorus; DO:溶解氧Dissolved oxygen; CODMn:高锰酸盐指数Chemical oxygen demand; Chl-a: 叶绿素a Chlorophyll a; WT:水温Water temperature; ABA:蓝藻水华面积Algal bloom area; 蓝实线为气象因子、蓝虚线为水质因子、红实线为响应因子 |
滇池外海、外海北部和外海中部区域环境因子解释量分别高达87.72%、90.32%和77.63%, 响应因子(ABA和Chl-a)之间存在正相关关系, 且环境因子对响应因子的影响体现了空间上的一致性, 即3个区域中AT、TP、WT、RH和CODMn与ABA存在显著正相关关系, WS、SD和PP与ABA存在显著负相关关系; RH、CODMn、WT、pH、DO和AT与Chl-a存在正相关关系, PP、WS、SD和TN与Chl-a存在显著负相关关系。对于外海南部区域, 环境因子解释量为62.14%, 响应因子Chl-a与ABA间无显著关系, AT、RH、TN、WT与ABA存在正相关关系, SD、WS、PP、DO和CODMn与ABA存负相关关系; Chl-a除了与RH、CODMn、WT、pH、DO和AT存在正相关关系外, 还与TP存在显著正相关关系, 而Chl-a仅与TN呈负相关关系。
为进一步分析各环境单因子对响应因子的贡献率, 确定各个分区蓝藻水华主要驱动因子, 采用随机森林法将环境因子(水质与气象)与响应因子进行分析, 并对两个响应因子进行主成分分析(PCA), 得到第一主成分解释度高达95.2%。将得到的第一主成分与环境因子进行随机森林法分析, 并按照影响程度将因子从上到下进行排序, 并剔除影响较小的因子。四个区域中显著贡献因子略有不相同, 全模型显著性P均小于0.001(图 7)。滇池外海区域对响应因子影响最大的因素是WS, 其后依次为WT、SD、RH等, 解释度为54.37%;外海北部区域对响应因子影响从大到小排列依次为WS、SD、WT、RH等, 其解释度为58.05%;相应的外海中部从大到小排列依次为WS、SD、WT、RH等, 解释度34.43%;相应的外海南部从大到小排列依次为WS、WT、SD、PP等, 解释度达69.55%。

|
| 图 7 随机森林法识别水华主要驱动因素 Fig. 7 Identification of main driving factors for algal boom by random forest at Waihai |
综合以上分析结果, 四个区域中气象因子为主要影响因素, 水质因子次之, 且在四个区域中, WS在气象因子中贡献率均为最大。
3.3.2 风速对水华面积的影响为进一步分析WS对滇池不同区域蓝藻水华的影响, 将所有水华事件发生时的WS数据进行统计, 按照0≤WS≤1 m/s、1 < WS≤2 m/s、2 < WS≤3 m/s、3 < WS≤4 m/s、4 < WS≤5 m/s和WS>5 m/s共6个区间进行划分, 统计各个区间平均水华面积比例, 结果如图 8。随风速增加, 水华发生频率呈先增加后减少的单峰型变化特征。当WS在0—1 m/s区间时, 水华发生频率最低, 仅发生17起水华事件; 当WS位于2—3 m/s区间时, 水华发生频率最高, 共发生353起水华事件; 而当WS>3 m/s时, 水华发生频率逐渐减少。滇池外海北部、中部、南部3个区域的水华面积比例随风速增加呈下降趋势, 即当WS处于0—1 m/s区间时, 3个区域水华面积比例最大, 其中外海北部处于轻度水华状态; 当WS>5 m/s时, 所有区域水华面积比例最小。

|
| 图 8 外海不同风速条件下水华发生规律特征统计 Fig. 8 Statistical characteristics of algal bloom with different wind speed at Waihai |
统计上述不同风速区间在春、夏、秋、冬四季的主导风向(图 9)。在春季, 1—2 m/s风速区间的主导风向为南南西风, 风向频率为36.4%;其他风速区间均以西南偏西为主; 在夏季, 1—2 m/s风速区间以西南风为主, 风向频率33.3%;2—3 m/s风速区间以东南偏南为主, 风向频率为100%;在秋季, 当风速处于0—1 m/s和>5 m/s区间时, 分别东南偏南和东南风为主, 风向频率分别为55.6%和100%;其余四个风速区间主导风向均为西风或南风。在冬季, 六个风速区间主导风向为西南偏西、西南或者西南偏南。

|
| 图 9 外海水华发生期间不同风速范围下的主导风向 Fig. 9 Prevailing wind direction for different wind speed ranges during algal bloom for Waihai |
综上所述, 在2002—2018年滇池流域发生的所有水华事件期间, 不同风速区间在四个季节的主导风向大体相同, 均以西南风为主。
3.3.4 WRT对水华发生的影响通过AEM3D模型, 计算了2007—2008年间每次水华发生时整个外海、外海北部、外海中部和外海南部的WRT, 将其与对应的ABA进行斯皮尔曼相关分析, 结果见表 4。整个外海和外海北部区域ABA与WRT均呈极显著正相关(P < 0.01), 外海中部和外海南部区域ABA与WRT呈显著正相关(0.01 <P < 0.05), 这表明ABA随WRT的增长而不断增加。
| 相关系数 Correlation coefficient |
外海WRT WRT of Waihai |
外海北部WRT WRT of North |
外海中部WRT WRT of Middle |
外海南部WRT WRT of South |
| 外海水华面积ABA of Waihai | 0.121** | |||
| 外海北部水华面积ABA of North | 0.197** | |||
| 外海中部水华面积ABA of Middle | 0.091* | |||
| 外海南部水华面积ABA of South | 0.08* | |||
| WRT:水力滞留时间Water Residence Time; *表示相关性达显著水平(P < 0.05);**表示相关性达极显著水平(P < 0.01);样本数n=733 | ||||
从WRT的空间分布来看(图 10), 在滇池外海水体中, WRT由北向南逐渐增长; 在外海北部水体中, 西部较东部水力滞留时间更长, 这可能是因北部区域中15条入湖河流均由东北部注入外海所致, 且包含流量最大的盘龙江; 在外海中部和南部水体中, 东部和西部WRT无明显差别。

|
| 图 10 2007—2018年滇池外海区域平均水力滞留时间 Fig. 10 Average water residence time of Waihai during 2007—2018 |
(1) 滇池外海蓝藻水华空间异质性
在滇池, 营养盐和水生生物分布的生态系统格局异质性特征明显[47], 由北向南逐渐减轻[21, 48—50], 这与本文得出滇池水华严重程度由北到南逐渐减轻变化趋势一致(图 5)。已有研究表明[51], 湖泊流域的主导风向和湖体的水动力条件(如WRT)共同驱动了蓝藻水华的空间分布。风应力可使大型浅水湖泊产生风生流, 并且驱动漂浮物沿水面进行移动和聚集[52—55], 且蓝藻漂移聚集区主要由主导风向和风速决定[56—59]。而风速除了推动蓝藻水华的堆积, 还可通过改变湖泊内部垂向扰动和物质通量从而影响湖体内的化学和生物过程[59]。对于大型浅水湖泊, 当风速大于4 m/s时, 湍流混合强烈, 可导致蓝藻的垂直上浮受到显著抑制[53], 而当风速低于3 m/s时, 蓝藻易上浮并聚集在表层[60]。
研究表明, WRT越长越有利于水体中藻类的生长和繁殖[61—62], 这与本文得到的滇池外海各区域WRT与水华面积呈显著正相关非常吻合(表 4)。在空间上, 外海WRT呈“北部 < 中部 < 南部”的分布模式, 因此北部的WRT较中南部较短, 但北部又是水华高发区且水华相对严重, 其原因是由于滇池外海常年盛行西南风所致, 使得北部区域成为藻类沿水表层漂移后的最可能聚集地。从3个湖区入湖水量分析, 北部区域中15条入湖河流占总入湖水量的84.9%, 而中部和南部区域的9条河流仅占15.1%。此外, 2013年12月牛栏江—滇池引水工程正式投入使用[63], 补水经盘龙江注入外海北部区域, 截至2019年12月31日, 已累计向滇池补水34.1亿m3。因此, 北部河流的密集及补水工程造成了北部区域WRT相对较短的局面[64]。因此, 在水华的预测预警中, 除了需关注气象条件外, 外海北部的水文条件也需重点考虑。
(2) 蓝藻水华驱动因子分析
一般情况下, 藻类水华是由于相对丰富的营养盐条件, 加上适宜的气象和水动力条件下导致藻类过度繁殖和漂移聚集而形成。铜绿微囊藻(属蓝藻门)作为滇池的优势种[65], 当湖体中TN和TP浓度分别低于2 mg/L和0.2 mg/L时, TN和TP浓度增加可以促进其生长, 但当浓度超过上述阈值时, 对其生长几乎无促进作用[65—66]。在研究时段内, 滇池外海绝大多数年份TN和TP平均浓度均超过其阈值, 这表明滇池水体营养盐浓度已经不再是主要限制因子, 因此气象和水动力条件成为关键影响因子[24], 例如水温、水表面风场条件(混合)和日照[58, 67—68]。这些结果, 均与本文所得到的结果一致。
本文在月尺度对水华暴发影响因素进行了研究, 发现WS、WT和SD是影响整个外海、外海北部、外海中部和外海南部4个区域蓝藻水华的主要驱动力(图 7), 其中本文得到WS和SD是影响蓝藻水华的最重要因素的结论, 与以往研究结果一致[8, 21, 69]。有研究认为[70], 当全年WS≤3 m/s时, 会促进滇池蓝藻水华暴发, 而目前滇池流域多年平均风速小于3 m/s的临界风速, 具备藻类上浮的水动力条件。SD与太阳辐射能密切相关, 是藻类(如蓝藻)进行光合作用, 满足自身生长需求的必要条件[32], 有学者[30]对巢湖水华驱动因素进行分析, 发现SD与ABA呈正相关关系, 与本文到结论相悖, 这可能由于滇池地处云贵高原, 海拔高于巢湖, 紫外线相对较强, 可能存在藻类的光抑制作用。因此, 有关云贵高原和长江中下游湖泊中藻类在生长机制和垂直迁移机制方面的差异, 是一个非常值得研究的科学问题。在同时具备充足的养分和有利的气候条件时, WT是直接控制滇池蓝藻水华暴发的关键因素[9]。当WT>20℃时, 利于蓝藻生长[1, 71]。目前滇池年内5—8月WT均大于20℃, 且随着全球变暖, 滇池流域气温具有每10年增加0.48℃的趋势[21], 因此可能导致水体中藻类生物量具有增加的趋势, 全球气候变化对高原富营养化湖泊水华的影响值得进一步深入研究。
目前滇池具有充足的氮磷营养盐条件, 供藻类吸收利用, 因此气象水文和水动力条件无疑成为水华暴发的主要驱动因素。本文虽然通过分析滇池外海不同区域的水华特征, 得到了上述主要结论。但滇池流域一年四季气温相对稳定导致水温长时间适宜藻类生长的特点, 与长江中下游湖泊中藻类生长机制和季节变化有何差异?以及高原较强的紫外线对藻类的生长和上浮机制是否具有抑制作用?结合滇池流域的气候特点, 滇池水体中营养盐浓度控制至何种水平才能有效防止水华的发生?尚待广大学者进一步深入研究。
5 结论基于2002—2018年MODIS卫星遥感数据, 本文对滇池外海蓝藻水华暴发的时空变化特征及不同分区驱动因子进行分析。结果表明:(1)滇池外海区域年平均水华面积比例呈逐渐下降趋势, 且由北向南水华严重程度逐渐减轻, 水华暴发面积最大主要发生在秋季。(2)在外海北部区域, 东部水华较西部水华更为严重; 在外海中部和南部区域中, 呈现西部水华较东部水华更为严重的空间分布模式。(3)气象因子为外海北部、中部和南部蓝藻水华暴发的主要影响因素, 水质因子次之。风速、水温和日照时长是上述区域中蓝藻水华暴发的决定性因素。(4)外海各区域中水力滞留时间与水华面积呈显著正相关。由北向南, 水力滞留时间逐渐增长。
| [1] |
Paerl H W, Otten T G. Harmful cyanobacterial blooms: causes, consequences, and controls. Microbial Ecology, 2013, 65(4): 995-1010. DOI:10.1007/s00248-012-0159-y |
| [2] |
Xiao M, Li M, Reynolds C S. Colony formation in the cyanobacterium Microcystis. Biological Reviews, 2018, 93(3): 1399-1420. DOI:10.1111/brv.12401 |
| [3] |
Kudela R M, Palacios S L, Austerberry D C, Accorsi E K, Guild L S, Torres-Perez J. Application of hyperspectral remote sensing to cyanobacterial blooms in inland waters. Remote Sensing of Environment, 2015, 167: 196-205. DOI:10.1016/j.rse.2015.01.025 |
| [4] |
江敏, 王婧, 许慧. 蓝藻毒素去除方法研究进展. 生态学杂志, 2014, 33(12): 3455-3462. DOI:10.13292/j.1000-4890.2014.0313 |
| [5] |
马健荣, 邓建明, 秦伯强, 龙胜兴. 湖泊蓝藻水华发生机理研究进展. 生态学报, 2013, 33(10): 3020-3030. |
| [6] |
中华人民共和国生态环境部. 2020年中国生态环境状况公报. [2021-05-26]. https://www.mee.gov.cn/hjzl/sthjzk/zghjzkgb/.
|
| [7] |
欧阳志宏, 郭怀成, 王婉晶, 高伟. 1982—2012年滇池水质变化及社会经济发展对水质的影响. 中国环境监测, 2015, 31(2): 68-73. DOI:10.3969/j.issn.1002-6002.2015.02.014 |
| [8] |
Jing Y Y, Zhang Y C, Hu M Q, Chu Q, Ma R H. MODIS-satellite-based analysis of long-term temporal-spatial dynamics and drivers of algal blooms in a plateau lake Dianchi, China. Remote Sensing, 2019, 11(21): 2582. DOI:10.3390/rs11212582 |
| [9] |
Sheng H, Liu H, Wang C Y, Guo H C, Liu Y, Yang Y H. Analysis of cyanobacteria bloom in the Waihai part of Dianchi Lake, China. Ecological Informatics, 2012, 10: 37-48. DOI:10.1016/j.ecoinf.2012.03.007 |
| [10] |
Shan K, Li L, Wang X X, Wu Y L, Hu L L, Yu G L, Song L R. Modelling ecosystem structure and trophic interactions in a typical cyanobacterial bloom-dominated shallow Lake Dianchi, China. Ecological Modelling, 2014, 291: 82-95. DOI:10.1016/j.ecolmodel.2014.07.015 |
| [11] |
Li X L, Janssen A B G, de Klein J J M, Kroeze C, Strokal M, Ma L, Zheng Y. Modeling nutrients in lake Dianchi (China) and its watershed. Agricultural Water Management, 2019, 212: 48-59. DOI:10.1016/j.agwat.2018.08.023 |
| [12] |
Zhang T, Zeng W H, Wang S R, Ni Z K. Temporal and spatial changes of water quality and management strategies of Dianchi Lake in southwest China. Hydrology and Earth System Sciences, 2014, 18(4): 1493-1502. DOI:10.5194/hess-18-1493-2014 |
| [13] |
Merel S, Walker D, Chicana R, Snyder S, Baurès E, Thomas O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environment International, 2013, 59: 303-327. DOI:10.1016/j.envint.2013.06.013 |
| [14] |
Wu Y L, Li L, Gan N Q, Zheng L L, Ma H Y, Shan K, Liu J, Xiao B D, Song L R. Seasonal dynamics of water bloom-forming Microcystis morpho species and the associated extracellular microcystin concentrations in large, shallow, eutrophic Dianchi Lake. Journal of Environmental Sciences, 2014, 26(9): 1921-1929. DOI:10.1016/j.jes.2014.06.031 |
| [15] |
Ma H Y, Wu Y L, Gan N Q, Zheng L L, Li T L, Song L R. Growth inhibitory effect of Microcystis on Aphanizomenon flos-aquae isolated from cyanobacteria bloom in Lake Dianchi, China. Harmful Algae, 2015, 42: 43-51. DOI:10.1016/j.hal.2014.12.009 |
| [16] |
Zhao C S, Shao N F, Yang S T, Ren H, Ge Y R, Zhang Z S, Feng P, Liu W L. Quantitative assessment of the effects of human activities on phytoplankton communities in lakes and reservoirs. Science of the Total Environment, 2019, 665: 213-225. DOI:10.1016/j.scitotenv.2019.02.117 |
| [17] |
Wang J H, Wang Y N, Dao G H, Du J S, Han Y P, Hu H Y. Decade-long meteorological and water quality dynamics of northern Lake Dianchi and recommendations on algal bloom mitigation via key influencing factors identification. Ecological Indicators, 2020, 115: 106425. DOI:10.1016/j.ecolind.2020.106425 |
| [18] |
胡旻琪, 张玉超, 马荣华, 张壹萱. 巢湖2016年蓝藻水华时空分布及环境驱动力分析. 环境科学, 2018, 39(11): 4925-4937. DOI:10.13227/j.hjkx.201801057 |
| [19] |
Zhang J, Chen L Q, Chen X L. Monitoring the cyanobacterial blooms based on remote sensing in Lake Erhai by FAI. Journal of Lake Sciences, 2016, 28(4): 718-725. DOI:10.18307/2016.0404 |
| [20] |
Woźniak M, Bradtke K, Darecki M, Kręel A. Empirical model for phycocyanin concentration estimation as an indicator of cyanobacterial bloom in the optically complex coastal waters of the Baltic Sea. Remote Sensing, 2016, 8(3): 212. DOI:10.3390/rs8030212 |
| [21] |
Wang J H, Yang C, He L Q S, Dao G H, Du J S, Han Y P, Wu G X, Wu Q Y, Hu H Y. Meteorological factors and water quality changes of Plateau Lake Dianchi in China (1990-2015) and their joint influences on cyanobacterial blooms. Science of the Total Environment, 2019, 665: 406-418. DOI:10.1016/j.scitotenv.2019.02.010 |
| [22] |
王菁晗, 何吕奇姝, 杨成, 刀国华, 杜劲松, 韩亚平, 吴光学, 吴乾元, 胡洪营. 太湖、巢湖、滇池水华与相关气象、水质因子及其响应的比较(1981—2015年). 湖泊科学, 2018, 30(4): 897-906. |
| [23] |
Liu Y, Wang Z, Guo H C, Yu S X, Sheng H. Modelling the effect of weather conditions on cyanobacterial bloom outbreaks in lake Dianchi: a rough decision-adjusted logistic regression model. Environmental Modeling & Assessment, 2013, 18(2): 199-207. |
| [24] |
鲁韦坤, 余凌翔, 欧晓昆, 李发荣. 滇池蓝藻水华发生频率与气象因子的关系. 湖泊科学, 2017, 29(3): 534-545. |
| [25] |
Zhao D, Li J S, Hu R M, Shen Q, Zhang F F. Landsat-satellite-based analysis of spatial-temporal dynamics and drivers of CyanoHABs in the plateau Lake Dianchi. International Journal of Remote Sensing, 2018, 39(23): 8552-8571. DOI:10.1080/01431161.2018.1488289 |
| [26] |
Hu C M. A novel ocean color index to detect floating algae in the global oceans. Remote Sensing of Environment, 2009, 113(10): 2118-2129. DOI:10.1016/j.rse.2009.05.012 |
| [27] |
Kiage L M, Walker N D. Using NDVI from MODIS to monitor duckweed bloom in lake Maracaibo, Venezuela. Water Resources Management, 2009, 23(6): 1125-1135. DOI:10.1007/s11269-008-9318-9 |
| [28] |
Zhang Y C, Ma R H, Duan H T, Loiselle S A, Xu J D, Ma M X. A novel algorithm to estimate algal bloom coverage to subpixel resolution in lake Taihu. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing, 2014, 7(7): 3060-3068. DOI:10.1109/JSTARS.2014.2327076 |
| [29] |
Shi K, Zhang Y L, Zhou Y Q, Liu X H, Zhu G W, Qin B Q, Gao G. Long-term MODIS observations of cyanobacterial dynamics in Lake Taihu: Responses to nutrient enrichment and meteorological factors. Scientific Reports, 2017, 7: 40326. DOI:10.1038/srep40326 |
| [30] |
Zhang Y C, Ma R H, Zhang M, Duan H T, Loiselle S, Xu J D. Fourteen-year record (2000-2013) of the spatial and temporal dynamics of floating algae blooms in lake Chaohu, observed from time series of MODIS images. Remote Sensing, 2015, 7(8): 10523-10542. DOI:10.3390/rs70810523 |
| [31] |
Wang S Q, Zhu L, Li Q, Li G B, Li L, Song L R, Gan N Q. Distribution and population dynamics of potential anatoxin-a-producing cyanobacteria in Lake Dianchi, China. Harmful Algae, 2015, 48: 63-68. DOI:10.1016/j.hal.2015.07.005 |
| [32] |
Zhou Q C, Zhang Y L, Lin D M, Shan K, Luo Y, Zhao L, Tan Z W, Song L R. The relationships of meteorological factors and nutrient levels with phytoplankton biomass in a shallow eutrophic lake dominated by cyanobacteria, Lake Dianchi from 1991 to 2013. Environmental Science and Pollution Research International, 2016, 23(15): 15616-15626. DOI:10.1007/s11356-016-6748-4 |
| [33] |
盛虎, 郭怀成, 刘慧, 杨永辉. 滇池外海蓝藻水华爆发反演及规律探讨. 生态学报, 2012, 32(1): 56-63. DOI:10.3969/j.issn.1673-1182.2012.01.013 |
| [34] |
祝艳, 韩林沛, 刘丽君, 邱在容, 王永肖, 刘雨雪, 何依, 王涵. 2018年滇池水环境质量评价研究. 环境科学导刊, 2020, 39(2): 75-85. DOI:10.13623/j.cnki.hkdk.2020.02.021 |
| [35] |
王明净, 杜展鹏, 段仲昭, 严长安, 高伟, 刘嫦娥. 河湖生态系统生态用水优化研究——以滇池流域为例. 生态学报, 2021, 41(4): 1341-1348. |
| [36] |
张虎才, 常凤琴, 段立曾, 李华勇, 张云鹰, 蒙红卫, 文新宇, 吴汉, 路志明, 毕荣鑫, 张扬, 赵帅营, 康文刚. 滇池水质特征及变化. 地球科学进展, 2017, 32(6): 651-659. |
| [37] |
高伟, 杜展鹏, 严长安, 陈岩. 污染湖泊生态系统服务净价值评估——以滇池为例. 生态学报, 2019, 39(5): 1748-1757. |
| [38] |
王杰, 毛建忠, 谢永红, 曹言, 段琪彩. 2008—2014年滇池水质时空变化特征分析. 人民长江, 2018, 49(5): 11-15. |
| [39] |
Chavula G, Brezonik P, Thenkabail P, Johnson T, Bauer M. Estimating chlorophyll concentration in Lake Malawi from MODIS satellite imagery. Physics and Chemistry of the Earth, Parts A/B/C, 2009, 34(13/14/15/16): 755-760. |
| [40] |
Guzmán V R, Fernando G S. Using MODIS 250 m imagery to estimate total suspended sediment in a tropical open bay. International Journal of Systesms Applications, Engineering and Development, 2009, 3(1): 36-44. |
| [41] |
Le C F, Hu C M, English D, Cannizzaro J, Kovach C. Climate-driven chlorophyll-a changes in a turbid estuary: observations from satellites and implications for management. Remote Sensing of Environment, 2013, 130: 11-24. DOI:10.1016/j.rse.2012.11.011 |
| [42] |
Hu C M, Chen Z Q, Clayton T D, Swarzenski P, Brock J C, Muller-Karger F E. Assessment of estuarine water-quality indicators using MODIS medium-resolution bands: initial results from Tampa Bay, FL. Remote Sensing of Environment, 2004, 93(3): 423-441. DOI:10.1016/j.rse.2004.08.007 |
| [43] |
Hu C M, Lee Z P, Ma R H, Yu K, Li D Q, Shang S L. Moderate Resolution Imaging Spectroradiometer (MODIS) observations of cyanobacteria blooms in Taihu Lake, China. Journal of Geophysical Research: Oceans, 2010, 115(C4). |
| [44] |
Robson B J, Hamilton D P. Three-dimensional modelling of a Microcystis bloom event in the Swan River Estuary, Western Australia. Ecological Modelling, 2004, 174(1/2): 203-222. |
| [45] |
Ter Braak C J F, Prentice I C. A theory of gradient analysis. Advances in Ecological Research. Amsterdam: Elsevier, 1988: 271-317.
|
| [46] |
Breiman L. Random forests. Machine Learning, 2001, 45(1): 5-32. |
| [47] |
李根保, 李林, 潘珉, 谢志才, 李宗逊, 肖邦定, 刘贵华, 陈静, 宋立荣. 滇池生态系统退化成因、格局特征与分区分步恢复策略. 湖泊科学, 2014, 26(4): 485-496. |
| [48] |
陈永川, 汤利, 张德刚, 周军, 李杰. 滇池叶绿素a的时空变化及水体磷对藻类生长的影响. 农业环境科学学报, 2008, 27(4): 1555-1560. |
| [49] |
凌祯, 史正涛, 徐杉, 董明华, 苏斌, 李婕. 滇池湖滨带叶绿素a与营养盐空间分布特征. 水文, 2020, 40(1): 76-80. |
| [50] |
李蒙, 谢国清, 鲁韦坤, 戴丛蕊. 气象条件对滇池水华分布的影响. 气象科学, 2011, 31(5): 639-645. |
| [51] |
Zhang Y C, Loiselle S, Shi K, Han T, Zhang M, Hu M Q, Jing Y Y, Lai L, Zhan P F. Wind effects for floating algae dynamics in eutrophic lakes. Remote Sensing, 2021, 13(4): 800. |
| [52] |
Ndong M, Bird D, Nguyen-Quang T, de Boutray M L, Zamyadi A, Vinçon-Leite B, Lemaire B J, Prévost M, Dorner S. Estimating the risk of cyanobacterial occurrence using an index integrating meteorological factors: application to drinking water production. Water Research, 2014, 56: 98-108. |
| [53] |
Cao H S, Kong F X, Luo L C, Shi X L, Yang Z, Zhang X F, Tao Y. Effects of wind and wind-induced waves on vertical phytoplankton distribution and surface blooms of Microcystis aeruginosa in lake Taihu. Journal of Freshwater Ecology, 2006, 21(2): 231-238. |
| [54] |
Moreno-Ostos E, Cruz-Pizarro L, Basanta A, George D G. The influence of wind-induced mixing on the vertical distribution of buoyant and sinking phytoplankton species. Aquatic Ecology, 2009, 43(2): 271-284. |
| [55] |
马巍, 浦承松, 罗佳翠, 陈欣. 滇池水动力特性及其对北岸蓝藻堆积驱动影响. 水利学报, 2013, 44(S1): 22-27. |
| [56] |
Wu T F, Qin B Q, Zhu G W, Luo L C, Ding Y Q, Bian G Y. Dynamics of cyanobacterial bloom formation during short-term hydrodynamic fluctuation in a large shallow, eutrophic, and wind-exposed Lake Taihu, China. Environmental Science and Pollution Research International, 2013, 20(12): 8546-8556. |
| [57] |
Webster I T. Effect of wind on the distribution of phytoplankton cells in lakes. Limnology and Oceanography, 1990, 35(5): 989-1001. |
| [58] |
Hunter P D, Tyler A N, Willby N J, Gilvear D J. The spatial dynamics of vertical migration by Microcystis aeruginosa in a eutrophic shallow lake: a case study using high spatial resolution time-series airborne remote sensing. Limnology and Oceanography, 2008, 53(6): 2391-2406. |
| [59] |
张晨, 来世玉, 高学平, 刘汉安. 气候变化对湖库水环境的潜在影响研究进展. 湖泊科学, 2016, 28(4): 691-700. |
| [60] |
Zhang M, Kong F X, Wu X D, Xing P. Different photochemical responses of phytoplankters from the large shallow Taihu Lake of subtropical China in relation to light and mixing. Hydrobiologia, 2008, 603(1): 267-278. |
| [61] |
Hu H P, Jiang R F, Wen T H, Lu Y S. Design and operation of semi-closed water flowing project with tidal influence. China Water & Wastewater, 2019, 35(16): 103-106. |
| [62] |
Sullivan B E, Prahl F G, Small L F, Covert P A. Seasonality of phytoplankton production in the Columbia River: a natural or anthropogenic pattern?. Geochimica et Cosmochimica Acta, 2001, 65(7): 1125-1139. |
| [63] |
张宇, 何苗. 2014—2018年滇池盘龙江水质变化特征. 环境科学导刊, 2020, 39(1): 1-4. |
| [64] |
王东旭, 晏欣. 牛栏江-滇池补水工程水源区水资源保护现状及对策. 水利水电快报, 2020, 41(10): 67-69, 79. |
| [65] |
Xu H, Paerl H W, Qin B, Zhu G, Hall N S, Wu Y. Determining critical nutrient thresholds needed to control harmful cyanobacterial blooms in eutrophic Lake Taihu, China. Environmental Science & Technology, 2015, 49(2): 1051-1059. |
| [66] |
Zhao C S, Shao N F, Yang S T, Ren H, Ge Y R, Feng P, Dong B E, Zhao Y. Predicting cyanobacteria bloom occurrence in lakes and reservoirs before blooms occur. Science of the Total Environment, 2019, 670: 837-848. |
| [67] |
任健, 商兆堂, 蒋名淑, 秦铭荣, 蒋薇. 2007年太湖蓝藻暴发的气象条件分析. 安徽农业科学, 2008, 36(27): 11874-11875, 11877. |
| [68] |
孔繁翔, 胡维平, 谷孝鸿, 杨桂山, 范成新, 陈开宁. 太湖梅梁湾2007年蓝藻水华形成及取水口污水团成因分析与应急措施建议. 湖泊科学, 2007, 19(4): 357-358. |
| [69] |
Wang Q, Sun L, Zhu Y, Wang S B, Duan C Y, Yang C J, Zhang Y M, Liu D J, Zhao L, Tang J L. Hysteresis effects of meteorological variation-induced algal blooms: a case study based on satellite-observed data from Dianchi Lake, China (1988-2020). Science of the Total Environment, 2022, 812: 152558. |
| [70] |
Ma X X, Wang Y A, Feng S Q, Wang S B. Vertical migration patterns of different phytoplankton species during a summer bloom in Dianchi Lake, China. Environmental Earth Sciences, 2015, 74(5): 3805-3814. |
| [71] |
Ibelings B W, Bormans M, Fastner J, Visser P M. CYANOCOST special issue on cyanobacterial blooms: synopsis—a critical review of the management options for their prevention, control and mitigation. Aquatic Ecology, 2016, 50(3): 595-605. |