 2023, Vol. 43
2023, Vol. 43文章信息
- 刘雨, 高光耀, 李宗善, 王聪, 田立德
- LIU Yu, GAO Guangyao, LI Zongshan, WANG Cong, TIAN Lide
- 内蒙古荒漠草原植物水分利用特征差异及对环境因子的响应
- Differences in plant water use characteristics and responses to environmental factors in the desert grassland of the Inner Mongolia
- 生态学报. 2023, 43(19): 7924-7935
- Acta Ecologica Sinica. 2023, 43(19): 7924-7935
- http://dx.doi.org/10.20103/j.stxb.202211033147
-
文章历史
- 收稿日期: 2022-11-03
- 网络出版日期: 2023-05-15






2. 中国科学院生态环境研究中心 城市与区域生态国家重点实验室, 北京 100085;
3. 陕西黄土高原地球关键带国家野外科学观测研究站, 西安 710061
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
3. National Observation and Research Station of Earth Critical Zone on the Loess Plateau in Shaanxi, Xi'an 710061, China
草地是干旱半干旱地区的主要生态系统类型[1], 在防风固沙、水土保持和水源涵养等方面具有重要作用[2—3]。水分是干旱半干旱地区植被生长的主要限制因子[4], 其中土壤水是干旱半干旱地区草地生态系统的主要水分来源[5]。植物对土壤水的利用策略和水分利用效率是干旱半干旱区植物适应环境的重要表征[4], 与气候条件、土壤特性和植物特性(根系形态和生理特性)等密切相关[6—10]。研究干旱半干旱地区草地生态系统植物水分利用特征的差异及其对环境因子的响应, 有助于理解植物生存策略及对环境变化的响应机制, 对揭示变化环境下草地生态系统变化特征和适应策略具有重要意义。
稳定同位素技术是研究植物水分利用特征的有效手段。通过比较各种水源与植物水的δ2H和δ18O等同位素特征可以量化植物的水分利用来源[11], 采用植物叶片δ13C可有效指示植物水分利用效率(WUEi, Intrinsic water use efficiency)[4]。目前, 大多数关于干旱半干旱区荒漠草原植物水分利用特征的研究主要集中在物种间差异以及对气候因子的响应。Eggemeyer等[12]发现半干旱沙丘草原的草本植物主要从上层土壤剖面(0.05—0.5 m)吸收土壤水分, 在干旱条件下, 会增加对0.5 m以下土壤水的吸收。Hoekstra[13]发现在种间竞争机制下, 深根系植物红车轴草对于0—10 cm的土壤水吸收减少, 而浅根系植物黑麦草和白三叶吸水深度趋向于更浅。Song等[14]研究发现荒漠草原优势物种甘草在面对水分胁迫时, 提高了水分利用效率应对环境变化。Diao等[15]分析了叶片δ13C与气候梯度的关系, 发现叶片δ13C与年均降雨量的关系仅在温带干旱区呈显著正相关, 而在其他地区呈显著负相关。整体来看, 目前研究缺少对干旱半干旱区草原不同科、属植物水分利用特征差异的系统分析, 对植物水分利用特征与环境因子的定量关系需要进一步研究。
为此, 本文以内蒙古荒漠草原为研究区, 通过样地调查和同位素示踪方法, 比较不同科、属植物水分利用来源和利用效率的差异, 分析水分利用特征对环境因子(气候、土壤因子及植物特性)的响应关系, 为阐明变化环境下荒漠草原植物的响应策略以及生态系统管理提供科学依据。
1 材料与方法 1.1 研究区与样地设置研究区位于中国内蒙古荒漠草原, 纬度范围从北纬41°21′到43°34′, 经度范围为东经111°11′到116°41′, 高程为1000—1610 m。研究区属于温带半干旱大陆性气候, 年均降雨量在130 mm以下, 年均温为8—19℃。植被类型主要以草本植物为主。在2021年8月, 沿干旱梯度选择7个样地进行野外调查。在样点布设45 m×45 m的典型样地, 并在每个样地中布设4条间隔10 m的样线, 每条样线设置5个1.5 m×1.5 m的小样方。具体样地信息见表 1。
| 样地 Plots |
地点(经度, 纬度) Site (Longitude, Latitude)/ (°E, °N) |
高程 Elevation/m |
年均温 Average annual temperature/℃ |
年均降雨量 Average annual rainfall/mm |
年均最高气温 Average annual maximum temperature/℃ |
年均最低气温 Average annual minimum temperatures/℃ |
| 1 | 希拉穆仁(116.21, 41.36) | 1610 | 8.0 | 127.80 | 29.33 | -11.00 |
| 2 | 四子王旗(111.90, 41.78) | 1440 | 13.3 | 105.07 | 29.33 | -11.00 |
| 3 | 黑沙图嘎查(111.20, 42.65) | 1000 | 19.0 | 56.63 | 29.33 | -11.00 |
| 4 | 苏尼特右旗(112.97, 42.08) | 1390 | 13.0 | 90.20 | 36.33 | -5.67 |
| 5 | 锡林郭勒1(116.67, 43.55) | 1270 | 10.0 | 87.83 | 36.33 | -5.67 |
| 6 | 锡林郭勒2(116.56, 43.54) | 1170 | 10.0 | 87.83 | 41.67 | -0.67 |
| 7 | 多伦十三里滩(116.28, 42.05) | 1320 | 9.3 | 126.00 | 41.67 | -0.67 |
| 气象数据来自中国气象数据网(中国地面气候标准值年值数据集1981—2010年) | ||||||
样地植被调查中, 记录植物的株高、盖度等。采集各植物的叶片进行叶片指标测定, 包括叶片周长、叶片长度、叶片宽度、叶片厚度及叶面积指标, 均采用Yaxin-1241便携式叶面积仪(精度0.1 mm2)测量。样地植被特性见表 2。土壤样品采样深度为100 cm, 采样间隔为10 cm, 设置三个重复, 采样后装在自封袋中, 用于测量土壤全氮、全磷、有机碳、速效磷、无机磷等养分指标, 样地土壤属性见表 3。
| 样地 Plots |
植物 Plants |
科 Family |
属 Genus |
株高 Plant height/cm |
盖度 Cover/% |
多度 Abundance |
叶面积 Leaf area/ mm2 |
叶周长 Leaf circumference/mm |
叶长 Leaf length/mm |
叶宽 Blade width/mm |
叶厚 Blade thickness/mm |
| 1 | 糙隐子草(Cleistogenes squarrosa (Trin.) Keng) | 禾本科 | 隐子草属 | 5.50±2.10 | 4.31±3.65 | 22.42±12.77 | 25.13±6.31 | 25.37±6.10 | 25.00±1.73 | 1.63±0.21 | 0.11±0.00 |
| 针茅(Stipa capillata Linn) | 禾本科 | 针茅属 | 40.11±8.25 | 9.32±6.30 | 10.74±5.18 | — | — | — | — | 0.38±0.05 | |
| 狗娃花(Aster hispidus Thunb.) | 菊科 | 狗娃花属 | 19.00±5.49 | 3.06±3.52 | 7.85±8.36 | 16.00±3.24 | 19.13±7.63 | 12.33±0.58 | 1.77±0.25 | 0.29±0.04 | |
| 2 | 冷蒿(Artemisia frigida Willd.) | 菊科 | 蒿属 | 18.27±5.00 | 6.03±4.62 | 5.87±3.98 | — | — | 8.92±0.113 | 1.08±0.11 | 0.54±0.53 |
| 针茅(Stipa capillata Linn) | 禾本科 | 针茅属 | 20.90±4.50 | 31.55±16.36 | 12.45±5.16 | 15.07±10.98 | — | 112.00±73.63 | 0.33±0.06 | 0.40±0.06 | |
| 3 | 红砂(Reaumuria songarica (Pall.) Maxim.) | 柽柳科 | 琵琶柴属 | 24.75±7.16 | 35.00±9.60 | 13.10±5.25 | — | — | 2.71±0.05 | 0.49±0.06 | 0.49±0.06 |
| 盐爪爪(Kalidium foliatum (Pall.) Moq.) | 藜科 | 盐爪爪属 | 26.25±13.02 | 6.88±4.60 | 4.88±0.35 | — | — | 21.68±1.64 | 1.21±0.24 | 1.21±0.24 | |
| 4 | 栉叶蒿(Neopallasia pectinata (Pall.) Poljak.) | 菊科 | 栉叶蒿属 | 27.75±5.50 | 18.25±11.47 | 28.05±10.63 | 7.03±3.77 | 6.67±4.65 | 13.67±5.69 | 1.00±0.30 | 0.31±0.06 |
| 羽茅(Achnatherum sibiricum (L.) Keng) | 禾本科 | 芨芨草属 | 45.00±10.00 | 29.62±12.82 | 20.38±6.91 | 126.87±30.05 | 145.83±15.06 | 88.33±14.36 | 2.13±0.42 | 0.19±0.01 | |
| 5 | 白羊草(Bothriochloa ischaemum (Linnaeus) Keng) | 禾本科 | 孔颖草属 | 60.67±7.04 | 45.33±25.39 | 51.33±21.91 | 1067.20±65.40 | 485.13±18.00 | 255.00±15.87 | 6.07±0.25 | 0.29±0.02 |
| 6 | 大针茅(Stipa grandis P. Smirn.) | 禾本科 | 针茅属 | 101.76±16.67 | 25.18±14.23 | 29.71±11.38 | 201.17±87.36 | 264.43±118.08 | 172.00±26.51 | 1.77±0.45 | 0.22±0.01 |
| 黄花蒿(Artemisia annua) | 菊科 | 蒿属 | 66.25±5.18 | 4.31±2.69 | 9.50±5.40 | — | — | — | — | — | |
| 7 | 克氏针茅(Stipa krylovii Roshev Willd.) | 禾本科 | 针茅属 | 86.88±18.52 | 14.41±11.86 | 17.50±7.96 | — | — | — | — | — |
| 冷蒿(Artemisia frigida) | 菊科 | 蒿属 | 39.47±11.24 | 20.00±15.93 | 30.79±15.00 | — | — | 13.09±2.12 | 11.36±2.64 | 0.37±0.05 | |
| 菊叶委陵菜(Potentilla tanacetifolia Willd. ex Schlecht.) |
蔷薇科 | 委陵菜属 | 45.26±5.77 | 5.16±2.57 | 10.37±3.93 | 151.10±24.58 | 64.77±2.79 | 24.67±0.58 | 8.97±1.78 | 0.30±0.06 |
| 样地 Plots |
容重 Capacitive weight/ (g/cm3) |
粘粒 Sticky particles/% |
粉粒 Powder grains/% |
砂细 Fine sand/% |
粗砂 Coarse sand/% |
全氮 Total nitrogen/ (g/kg) |
全磷 Total phosphorus/ (g/kg) |
有机碳 Organic carbon/ (g/kg) |
速效磷 Available phosphorus/ (mg/kg) |
无机磷 Inorganic phosphorus/ (g/kg) |
| 1 | 1.36±0.01 | 8.29±0.82 | 18.58±1.46 | 57.50±2.93 | 15.63±2.16 | 0.79±0.54 | 0.56±0.20 | 9.18±5.03 | 2.24±0.35 | 0.34±0.21 |
| 2 | 1.16±0.24 | 10.40±1.69 | 20.77±1.79 | 52.66±5.13 | 16.18±5.01 | 0.76±0.73 | 0.26±0.09 | 7.87±6.52 | 3.81±2.29 | 0.12±0.01 |
| 3 | 1.39±0.20 | 14.91±3.69 | 26.51±4.48 | 43.71±4.28 | 14.88±11.75 | 0.41±0.24 | 0.23±0.08 | 4.83±2.27 | 5.38±4.12 | 0.13±0.04 |
| 4 | 1.49±0.03 | 13.82±0.37 | 24.77±3.05 | 47.87±4.86 | 28.82±2.59 | 0.61±0.36 | 0.25±0.08 | 6.71±3.98 | 2.11±0.53 | 0.12±0.02 |
| 5 | 0.84±0.12 | 9.41±1.02 | 17.94±2.51 | 63.64±2.49 | 9.01±5.90 | 0.83±0.72 | 0.27±0.05 | 9.51±8.19 | 2.15±1.05 | 0.14±0.02 |
| 6 | 1.10±0.08 | 10.31±2.42 | 17.97±5.75 | 54.48±8.17 | 17.23±16.24 | 0.72±0.60 | 0.22±0.07 | 7.84±6.09 | 2.29±0.61 | 0.11±0.01 |
| 7 | 1.15±0.05 | 8.71±3.91 | 14.58±8.08 | 37.23±4.60 | 39.48±16.32 | 0.55±0.62 | 0.12±0.09 | 6.12±7.07 | 2.05±1.38 | 0.07±0.01 |
同位素样品采集包括土壤样品、植物根茎结合部(茎秆)样品和植物叶片样品三部分。土壤样品采集深度、间隔等与上述一致, 分别采集两份, 一份保存在100 mL玻璃瓶中, 聚乙烯封口膜封口, 测量土壤水δ2H和δ18O值, 一份装在铝盒中, 用于测量土壤含水量(105℃, 24 h)。对于小灌木植物, 在样地中选择3株植物, 沿着植物的四个方向选取非绿色的栓化的枝条, 去除外部韧皮部, 剪成小段, 保存在100 mL玻璃瓶中, 并用封口膜封口, 设置4个重复。对于草本植物, 在样地中选取3—4株草本植物, 采集每株植物的根茎结合部, 同样置于玻璃瓶中, 设置4个重复。同时在设置的样地中选择8—10株植物, 沿不同方向采集各种植物的叶片, 置于自封袋中, 样品经烘干(70℃, 72 h)、研磨、过筛(100目筛)及再烘干(70℃, 48 h), 用于测量δ13C。
1.3 实验分析野外采集的土壤、根茎结合部(茎秆)样品全部密封在冰箱(-20℃)中保存, 进行水分抽提。水分抽提装置为全自动真空冷凝抽提装置(LI-2100, LICA, Beijing, China), 抽提时间和效率为3 h和98%。水分抽提后将水样保存在2 mL玻璃瓶中, 密封在冰箱(-4℃), 进行后续的同位素分析工作。采用LGR水同位素分析仪(DLT-100, Los Gatos Research, Mountain View, USA)测定土壤水中的δ2H(测试精度为±1.2‰)和δ18O(±0.3‰)。采用稳定同位素质谱仪(MAT253, Thermo Fisher Scientific, Bremen, Germany)测量植物根茎结合部(茎秆)水中的δ2H(±1‰)、δ18O(±0.2‰)与叶片δ13C值。稳定同位素值为[4]:
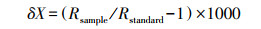
|
(1) |
式中, X可以代表2H, 18O, 13C。Rsample和Rstandard分别是样品和标准样品中的2H/1H, 18O/16O或者13C/12C。V-SMOW(Vienna Standard Mean Ocean Water)是2H和18O的标准物质, V-PDB(Vienna Pee Dee Belemnite)是13C的标准物质。
1.4 数据分析根据0—100 cm土层剖面土壤水δ2H和δ18O值及土壤含水量的变化情况及其变异程度, 将土层划分0—30 cm、30—60 cm和60—100 cm。利用贝叶斯同位素混合模型MixSIAR(R4.0.5软件中MixSIAR包)分析不同层次土壤水源对各植物根茎结合部(茎秆)水的贡献比例。模型中植物根茎结合部(茎秆)水δ2H和δ18O值作为混合源数据, 不同层次(0—30 cm、30—60 cm和60—100 cm)土壤水的δ2H和δ18O均值及标准差作为源数据, discrimination data设置为0, MCMC链设置为“long”。
利用SPSS 24.0中单因素方差分析(One-way ANOVA)对不同科、属植物土壤水来源比例及叶片δ13C的差异进行分析。由于环境变量较多, 且因子间存在多重共线性, 因此需要对其进行筛选。利用SPSS 24.0中Pearson相关分析计算环境因子之间的相关程度, 剔除共线性较强的因子。经过筛选, 气候因子包括年均降雨量和年均温, 植物因子包括株高、盖度、叶周长, 土壤因子包括容重、粗砂含量和土壤含水量。利用Origin 2018对所选环境因子与植物水分来源比例(水分利用效率)进行拟合, 寻求最优拟合模型。
2 结果与分析 2.1 土壤含水量变化特征如图 1所示, 不同样地的土壤含水量在2%—18%。希拉穆仁草地(样地1)土壤含水量随土壤深度的增加而逐渐增加, 黑沙图(样地3)、苏尼特(样地4)、锡林郭勒2(样地6)和多伦草地(样地7)土壤含水量随土壤深度的增加而逐渐降低, 四子王旗(样地2)、锡林郭勒1草地(样地5)土壤含水量随土壤深度的变化不大。方差分析表明, 不同样地之间土壤含水量具有显著性差异(P < 0.01)。

|
| 图 1 各样地土壤水含水量变化 Fig. 1 Changes of soil water content in various plots 样地1:希拉穆仁草地;样地2:四子王旗草地;样地3:黑沙图嘎查草地;样地4:苏尼特右旗草地;样地5:锡林郭勒草地1;样地6:锡林郭勒草地2;样地7:多伦草地; 不同小写字母表示不同样地间土壤含水量差异显著(P < 0.05) |
图 2所示, 希拉穆仁、四子王旗、黑沙图样地土壤水氢氧同位素随着土壤深度的增加呈减少的趋势, 苏尼特草地土壤水同位素随土壤深度的增加呈现先增加后减少的趋势, 锡林郭勒草地1和多伦草地整体呈现先减小后增加的趋势。方差分析表明, 不同样地之间土壤水同位素具有显著性差异(P < 0.01)。不同植物根茎结合部(茎秆)水氢氧同位素值具有显著差异(P < 0.05), 其中针茅同位素值在不同降雨梯度下具有显著差异(P < 0.01), 冷蒿(四子王旗草地)的δ2H、δ18O均值分别为-30.69‰和10.37‰, 显著高于多伦草地(-46.58‰和-3.85‰)(P < 0.05)。

|
| 图 2 各样地土壤水和植物根系结合部(茎秆) 水δ2H和δ18O变化 Fig. 2 Soil water and plant root junction (stem) water δ2H and δ18O changes in various plots 图中红色和蓝色分别代表各样地土壤水δ2H和δ18O值, 垂直实线和虚线分别代表各植物根茎结合部(茎秆)水δ2H和δ18O值 |
不同科植物对0—30 cm深度土壤水的吸收比例分别是55.63%(禾本科)、51.84%(菊科)和27.67%(其他科, 包括柽柳科、藜科和蔷薇科)(图 3)。禾本科和菊科植物对60—100 cm深度土壤水的利用比例分别为18.76%和11.76%, 显著低于其他科(36.83%)(P < 0.05)(图 3)。禾本科和菊科叶片δ13C均值分别为-26.61‰和-27.91‰, 低于其他科(-26.36‰)(图 4)。不同属植物水分利用特征没有显著性差异(P>0.05)。针茅属植物0—30 cm深度土壤水平均贡献比例(55.28%)、60—100 cm深度土壤水平均贡献比例(15.33%)高于蒿属(31%和14.33%), 而30—60 cm深度土壤水平均贡献比例则低于蒿属。针茅属植物叶片δ13C均值为-26.38‰, 高于蒿属(-28.14‰)。

|
| 图 3 不同科、属植物水分来源差异 Fig. 3 Differences in water sources of plants of different families and genera 不同小写字母表示不同科(属)植物间植物水分来源比例差异显著(P < 0.05) |

|
| 图 4 不同科、属植物叶片δ13C差异 Fig. 4 Differences in leaf δ13C in different families and genera |
0—30 cm深度土壤水贡献比例主要受年均降雨量、年均温、叶周长、粗砂含量等因素影响(P < 0.05)(图 5)。随着年均降雨量和粗砂含量的增加, 草本植物0—30 cm深度土壤水贡献比例整体呈现增加的趋势, 而年均温和叶周长增加时, 0—30 cm深度土壤水贡献比例呈现降低的趋势。30—60 cm深度土壤水贡献比例主要受年均降雨量和叶周长影响(R2>0.5, P < 0.05), 随着年均最高气温、叶周长的增加, 水分来源比例呈现增加的趋势, 年均降雨量与之相反(图 6)。60—100 cm深度土壤水贡献比例受年均温、年均降雨量和盖度显著影响(P < 0.05)(图 7)。随着年均温的增加, 60—100 cm深度土壤水贡献比例整体呈现增加的趋势, 年均降雨量则与之相反。叶片δ13C与气候、土壤和植物因子的相关性均不显著(P>0.05)(图 8)。

|
| 图 5 0—30 cm深度土壤水分贡献比例与气候、土壤和植物因子的关系 Fig. 5 Relationship between soil moisture contribution ratio and climate, soil and plant factors at a depth of 0—30 cm |

|
| 图 6 30—60 cm深度土壤水分贡献比例与气候、土壤和植物因子的关系 Fig. 6 Relationship between soil moisture contribution ratio and climate, soil and plant factors at a depth of 30—60 cm |

|
| 图 7 60—100 cm深度土壤水分贡献比例与气候、土壤和植物因子的关系 Fig. 7 Relationship between soil moisture contribution ratio and climate, soil and plant factors at a depth of 60—100 cm |

|
| 图 8 植物叶片δ13C与气候、土壤和植物因子的关系 Fig. 8 Relationship between plant leaf δ13C and climate, soil and plant factors |
本研究中禾本科与菊科对0—30 cm土壤水的平均吸收比例高于其他三科(柽柳科、藜科和蔷薇科), 而对60—100 cm土壤水的吸收比例显著低于其他科, 表明禾本科和菊科主要利用0—30 cm土壤水, 而其他科植物主要利用60—100 cm土壤水。不同生活型植物具有不同的水分来源[12]。禾本科中糙隐子草、针茅、大针茅、羽茅、白羊草和克氏针茅均为多年生草本植物;菊科中狗娃花为一年或二年生草本植物, 黄花蒿为一年生草本植物, 冷蒿为多年生草本植物, 栉叶蒿为一年或多年生草本植物;其他科中红砂(柽柳科)与盐爪爪(藜科)是小灌木植物, 菊叶委陵菜为多年生草本植物。在植物群落中, 灌草不同生活型植物具有不同的水分利用策略, 灌木对地下生物量投入较多, 对于中深层土壤水利用更多, 而草本植物对地上生物量投入更多, 对浅层土壤水利用更多[16—17]。针茅属主要利用0—30 cm土壤水, 蒿属主要利用30—60 cm土壤水。针茅属与蒿属均为草本植物, 其中四子王旗草地、锡林郭勒草地及多伦草地中均有针茅属和蒿属的植物, 针茅属植物高度均高于蒿属。同时四子王旗草地(锡林郭勒草地2)中针茅(大针茅)的盖度和多度均高于冷蒿(黄花蒿), 说明针茅对该区域环境的适应能力较强。
叶片上的气孔作为二氧化碳和水汽的通道, 控制植物蒸腾耗水和固碳间的平衡, 进而影响植物水分利用效率(WUEi)[18]。研究表明, 植物叶片δ13C值与WUEi之间存在正相关关系, 即δ13C值越高, WUEi越高[19]。在干旱半干旱地区, 水分是限制植物生长的主要因子, 植物水分利用效率(WUEi)决定了它们在群落中的竞争优势[20]。浅层土壤水受降雨补充, 干旱条件下植物易受水分胁迫影响[21—22], 深层土壤水受蒸发和降水影响较小, 对植物而言是相对稳定的水源。其他科植物δ13C高于禾本科和菊科植物, 同时获取较深层的水源, 保证了其在干旱环境中产生更多的干物质, 有利于自身生存[23]。针茅属叶片δ13C值高于蒿属, 其WUEi也高于蒿属, 表明针茅属的植物通过较高的水分利用效率而不是获取深层土壤水来适应环境。研究认为大针茅适应干旱环境的条件是将更多的资源分配给横向的扩展, 占据更大面积的土壤表面, 进而提高水分竞争能力和光合能力[24]。
3.2 植物水分利用特征对环境因子的响应植物水分利用特征受气候(降水和气温等)、土壤(土壤水分可用性等)以及植物自身特性的影响, 在不同环境中具有不同的水分利用状况。降水是干旱半干旱地区植物生长发育的限制因子。本研究发现, 年均降雨量是影响植物水分利用特征的主要因子。随着年均降雨量的增加, 植物对浅层(0—30 cm)土壤水的利用显著增加, 而对中深层(30—100 cm)土壤水的利用显著减少。随着降雨量的增加, 降雨对浅层土壤水的补给较易, 植物对浅层土壤水分利用更高[25]。年均温则与年降雨量呈现相反的趋势, 这可能是由于气温升高造成土壤表面蒸发增加[26], 促使植物吸收深层水分。在土壤特性方面, 降水会通过影响土壤水分[27], 间接影响植物水分利用特征。但是, 在本研究中土壤含水量对植物水分利用特征影响不大, 可能是样点较少的原因。同时土壤特性可以通过改变渗透系数和持水能力改变植物吸水模式[28—29]。随着粗砂含量的增加, 土壤孔隙度增加, 降水入渗率高, 植物对浅层土壤水的利用也显著增加。
植物水分来源除受降雨、土壤因素影响以外, 还与植物本身的特性相关, 比如植物叶片和根系。叶片是植物进行光合及蒸腾作用的重要器官[30], 叶周长越大, 其气孔可能越多, 蒸腾作用消耗的水分越多, 若浅层土壤水分无法满足植物需求, 可能会吸收更深层次的土壤水。根系是植物水分吸收的主要组织, 根系深度会对植物水分利用产生重要影响[31]。针茅根系生物量位于土层0—20 cm[32], 羊草根系分布集中层是0—40 cm[33], 红砂根系主要位于地下0—70 cm, 盐爪爪根系主要位于0—30 cm[34], 其水分利用特征有所差异。根系较深的植物能利用深层水源, 在水分胁迫环境下具有更强的竞争性适应环境变化[13]。植物叶片δ13C与各环境因子间并无显著相关性, 即水分利用效率随环境梯度的变化不大, 很可能是通过其他途径来适应环境变化, 具体原因还需进一步探究。研究结果初步反映了不同科、属植物水分利用特征的差异, 对探究干旱半干旱区植物的生态适应性及对荒漠草原的生态系统管理具有重要意义。在后续研究中, 应增加样点及采样次数对其进行分析, 有助于进一步理解干旱半干旱区荒漠草原植物的适应策略以及对环境变化的响应。
4 结论本研究利用稳定同位素技术和贝叶斯同位素混合模型(MixSIAR), 确定内蒙古荒漠草原各植物水分来源比例, 分析不同科、属植物水分利用特征的差异及其对环境因子的响应。主要结论如下:
(1) 不同科植物间水分利用特征存在差异。禾本科和菊科植物主要利用0—30 cm深度土壤水, 而其他三科(柽柳科、藜科和蔷薇科)植物主要利用60—100 cm深度土壤水。叶片δ13C表明, 其他科植物具有较高的水分利用效率, 增强了其他科植物在干旱条件下的适应性。
(2) 不同属植物水分利用特征不同。针茅属和蒿属植物主要土壤水分来源深度分别是0—30 cm和30—60 cm。针茅属植物水分利用效率高于蒿属, 表明其主要通过保持较高的水分利用效率而不是获取深层土壤水来适应干旱环境。
(3) 植物不同深度土壤水分贡献比例和叶片δ13C对不同环境因子的响应不同。其中, 年均降雨量、年均温和叶周长对植物水分来源比例有显著影响。随着年均降雨量的增加, 植物对0—30 cm土壤水的吸收比例显著增加, 对30—100 cm土壤水的利用逐渐减少。年均温和叶周长与之相反。
| [1] |
Tang X G, Ma M G, Ding Z, Xu X B, Yao L, Huang X J, Gu Q, Song L S. Remotely monitoring ecosystem water use efficiency of grassland and cropland in China's arid and semi-arid regions with MODIS data. Remote Sensing, 2017, 9(6): 616. DOI:10.3390/rs9060616 |
| [2] |
Bi X, Li B, Fu Q, Fan Y, Ma L X, Yang Z H, Nan B, Dai X H, Zhang X S. Effects of grazing exclusion on the grassland ecosystems of mountain meadows and temperate typical steppe in a mountain-basin system in Central Asia's arid regions, China. Science of the Total Environment, 2018, 630: 254-263. DOI:10.1016/j.scitotenv.2018.02.055 |
| [3] |
朱趁趁, 龚吉蕊, 杨波, 张子荷, 王彪, 矢佳昱, 岳可欣, 张魏圆. 内蒙古荒漠草原防风固沙服务变化及其驱动力. 生态学报, 2021, 41(11): 4606-4617. |
| [4] |
王剑. 基于稳定同位素技术的黄土丘陵区典型植物水分利用来源研究[D]. 北京: 中国科学院大学, 2019.
|
| [5] |
邹慧, 高光耀, 傅伯杰. 干旱半干旱草地生态系统与土壤水分关系研究进展. 生态学报, 2016, 36(11): 3127-3136. |
| [6] |
Liu Z Q, Yu X X, Jia G D. Water uptake by coniferous and broad-leaved forest in a rocky mountainous area of Northern China. Agricultural and Forest Meteorology, 2019, 265: 381-389. DOI:10.1016/j.agrformet.2018.11.036 |
| [7] |
Wang T, Xu Q, Zhang B B, Gao D Q, Zhang Y, Ren R R, Jiang J. Effects of understory removal and thinning on water uptake patterns in Pinus massoniana Lamb. plantations: evidence from stable isotope analysis. Forest Ecology and Management, 2022, 503: 119755. DOI:10.1016/j.foreco.2021.119755 |
| [8] |
Zhou H, Zhao W Z, He Z B, Yan J L, Zhang G F. Variation in depth of water uptake for Pinus sylvestris var. mongolica along a precipitation gradient in sandy regions. Journal of Hydrology, 2019, 577: 123921. DOI:10.1016/j.jhydrol.2019.123921 |
| [9] |
Meißner M, Köhler M, Schwendenmann L, Hölscher D, Dyckmans J. Soil water uptake by trees using water stable isotopes (δ2H and δ18O)-a method test regarding soil moisture, texture and carbonate. Plant and Soil, 2014, 376(1): 327-335. |
| [10] |
Dai Y, Zheng X J, Tang L S, Li Y. Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantonggut Desert. Plant and Soil, 2015, 389(1): 73-87. |
| [11] |
Wang S F, Gao X D, Yang M, Huo G P, Song X L, Siddique K H M, Wu P T, Zhao X N. The natural abundance of stable water isotopes method may overestimate deep-layer soil water use by trees. Hydrology and Earth System Sciences Discussions, 2022. DOI:10.5194/hess-2022-142 |
| [12] |
Eggemeyer K D, Awada T, Harvey F E, Wedin D A, Zhou X H, Zanner C W. Seasonal changes in depth of water uptake for encroaching trees Juniperus virginiana and Pinus ponderosa and two dominant C4 grasses in a semiarid grassland. Tree Physiology, 2009, 29(2): 157-169. |
| [13] |
Hoekstra N J, Finn J A, Hofer D, Lüscher A. The effect of drought and interspecific interactions on depth of water uptake in deep- and shallow-rooting grassland species as determined by δ18O natural abundance. Biogeosciences, 2014, 11(16): 4493-4506. DOI:10.5194/bg-11-4493-2014 |
| [14] |
Song K C, Hu H Y, Xie Y Z, Fu L. The effect of soil water deficiency on water use strategies and response mechanisms of Glycyrrhiza uralensis fisch. Plants: Basel, Switzerland, 2022, 11(11): 1464. |
| [15] |
Diao H Y, Wang A Z, Yuan F H, Guan D X, Wu J B. Changes in tree leaf δ13C along climatic and geographical gradients in China. Trees, 2022, 1-12. |
| [16] |
Dodd M B, Lauenroth W K, Welker J M. Differential water resource use by herbaceous and woody plant life-forms in a shortgrass steppe community. Oecologia, 1998, 117(4): 504-512. DOI:10.1007/s004420050686 |
| [17] |
Villagra P E, Giordano C V, Alvarez J, Bruñol J, Guevara A, Carmen S, Passera C B, Greco S A. To be a plant in the desert: water use strategies and water stress resistance in the Central Monte desert from Argentina. Ecologia Austral, 2011, 21(1): 29-42. |
| [18] |
刘宁. 川西亚高山森林生态系统水碳过程耦合与模拟[D]. 北京: 中国林业科学研究院, 2013.
|
| [19] |
Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, 1989, 40: 503-537. DOI:10.1146/annurev.pp.40.060189.002443 |
| [20] |
Diao H J, Kardol P, Dong K H, Wang C H. Effects of nitrogen addition and mowing on nitrogen- and water-use efficiency of Artemisia frigida in a grassland restored from an abandoned cropland. Journal of Plant Ecology, 2021, 14(3): 515-526. DOI:10.1093/jpe/rtab006 |
| [21] |
Asbjornsen H, Shepherd G, Helmers M, Mora G. Seasonal patterns in depth of water uptake under contrasting annual and perennial systems in the Corn Belt Region of the Midwestern U.S. Plant and Soil, 2008, 308(1/2): 69-92. |
| [22] |
Yang H, Auerswald K, Bai Y F, Han X G. Complementarity in water sources among dominant species in typical steppe ecosystems of Inner Mongolia, China. Plant and Soil, 2011, 340(1): 303-313. |
| [23] |
赵丹, 程军回, 刘耘华, 刘利利, 李瑞霞, 盛建东. 荒漠植物梭梭稳定碳同位素组成与环境因子的关系. 生态学报, 2017, 37(8): 2743-2752. |
| [24] |
鲍雅静, 曹明, 李政海, 郭鹏, 张靖, 秦洁. 羊草与大针茅根系构型对水分梯度响应的比较研究. 生态学报, 2019, 39(3): 1063-1070. |
| [25] |
Yang L, Chen L D, Wei W, Yu Y, Zhang H D. Comparison of deep soil moisture in two re-vegetation watersheds in semi-arid regions. Journal of Hydrology, 2014, 513: 314-321. |
| [26] |
Tang J W, Qi Y, Xu M, Misson L, Goldstein A H. Forest thinning and soil respiration in a ponderosa pine plantation in the Sierra Nevada. Tree Physiology, 2005, 25(1): 57-66. |
| [27] |
Wang C, Wang S, Fu B, Yang L, Li Z S. Soil moisture variations with land use along the precipitation gradient in the north-south transect of the loess plateau. Land Degradation and Development, 2017, 28: 926-935. |
| [28] |
Coners H, Leuschner C. In situ measurement of fine root water absorption in three temperate tree species—temporal variability and control by soil and atmospheric factors. Basic and Applied Ecology, 2005, 6(4): 395-405. |
| [29] |
Zhao Y, Wang L, Knighton J, Evaristo J, Wassen M. Contrasting adaptive strategies by Caragana korshinskii and Salix psammophila in a semiarid revegetated ecosystem. Agricultural and Forest Meteorology, 2021, 300: 108323. |
| [30] |
Hu D W, He S P, Sun G F, Jia Y H, Su Y H, Ma X J, Dev W, Nazir M F, Geng X L, Wang L R, Pan Z E, Chen B J, Li H G, Wang X Y, Pang B Y, Du X M. Integrating Genome-wide association and whole transcriptome analysis to reveal genetic control of leaf traits in Gossypium arboreum L. Genomics, 2022, 114(3): 110331. |
| [31] |
Schenk H J. Soil depth, plant rooting strategies and species' niches. The New Phytologist, 2008, 178(2): 223-225. |
| [32] |
柳隽瑶. 内蒙古主要针茅草地优势种植物根系的时空格局[D]. 北京: 中国科学院大学, 2019.
|
| [33] |
宋炳煜, 杨劼, 旭日, 乌江雨. 羊草群落的水分利用. 植物学报, 2003, 45(10): 1245-1250. |
| [34] |
杨昊天, 李新荣, 刘立超, 贾荣亮, 王增如, 李小军, 李刚. 荒漠草地4种灌木生物量分配特征. 中国沙漠, 2013, 33(5): 1340-1348. |