 2023, Vol. 43
2023, Vol. 43文章信息
- 王梦梦, 张丽华, 当知才让, 王宏, 杨尖拉毛, 赵锐锋
- WANG Mengmeng, ZHANG Lihua, DANG-ZHI Cairang, WANG Hong, YANG-JIAN Lamao, ZHAO Ruifeng
- 高寒湿地退化对植物群落特征与土壤特性的影响
- Effects of alpine wetland degradation on plant community characteristics and soil properties
- 生态学报. 2023, 43(19): 7910-7923
- Acta Ecologica Sinica. 2023, 43(19): 7910-7923
- http://dx.doi.org/10.20103/j.stxb.202210142926
-
文章历史
- 收稿日期: 2022-10-14
- 网络出版日期: 2023-02-22






2. 甘肃省绿洲资源环境与可持续发展重点实验室, 兰州 730070;
3. 甘肃黄河首曲国家级自然保护区管护中心, 玛曲 747300;
4. 甘肃省玛曲青藏高原土著鱼类自然保护区管理局, 玛曲 747300
2. Key Laboratory of Resource Environment and Sustainable Development of Oasis, Gansu Province, Lanzhou 730070, China;
3. The first meander of the Yellow River National Nature Reserve Management Center, Gansu Province, Maqu 747300, China;
4. Maqu Qinghai-Tibet Plateau Native Fish Nature Reserve Administration, Gansu Province, Maqu 747300, China
湿地是介于陆地和水体之间的一种特殊生态系统类型, 在各类生态系统中, 生态服务价值最高。其在保护生态环境和生物多样性、维持生态平衡、调节气候等方面具有极其重要的作用, 因此, 被称为“生物基因库”和“地球之肾”[1]。近年来, 随着气候变化和人类活动的增加, 导致湿地面积急剧减少, 湿地退化问题日益严重。湿地生态系统已成为世界上最受威胁的生态系统之一[2]。
近年来, 群落特征作为定量刻化湿地退化的重要手段, 且有关退化湿地植物群落特征的研究已广泛报道。如在赫克萨尔湿地的研究中发现, 人类活动和侵占致使湿地退化, 导致植物群落组成发生改变和毒杂草过度生长[3]。对于物种多样性的报道发现, 在意大利波河三角洲[4], 南非高原东缘[5]均发现随湿地退化程度加深, 湿地生物多样性丧失。植物多样性在很大程度上反映了植物群落结构和生境多样性的特征[6]。然而, 关于湿地退化对植物多样性的研究结果尚不一致。如在三江源隆宝沼泽湿地, 发现随着湿地退化程度加剧, 地上生物量显著降低, 物种多样性指数、均匀度指数和丰富度均逐渐增大[7]。而在黄河源区沼泽湿地退化的研究发现, 随退化程度增加, 物种丰富度指数、均匀度指数和多样性指数先升后降, 优势度指数先降后升[8]。为此, 亟需进一步探究植物群落特征, 尤以反映多样性各指数是如何响应湿地退化。
湿地退化引起的土壤退化不仅会导致土壤基本理化性质的变化, 还会引起养分元素生物地球化学循环的变化, 影响湿地植物的生长和生产力[9]。大量研究表明, 湿地退化程度的增加将显著降低土壤有机碳、全氮含量, 且随土层深度增加明显降低[10]。土壤电导率及pH值因湿地退化而显著增加, 且随土层深度下移明显[11]。然而, 林春英等[12]研究发现pH值随湿地退化程度增加而增加。另有学者发现, 电导率、pH值与有机碳、全氮之间存在负向关系[13]。植物群落特征及多样性变化受到土壤性质、草食性等环境因素的显著影响[14], 而植物群落的发展在很大程度上依赖于土壤理化特性的改变。然而, 一些学者认为盐分含量的变化是直接威胁植物多样性的关键因素[4]。土壤电导率也被认为是影响植物多样性的主控因子[15]。此外, 有关湿地的识别与分类[16]、土壤有机碳、氮组分及储量变化[17]等方面的研究已占有较大比重, 而对于土壤因子如何响应湿地退化过程, 以及主导植物群落结构的关键土壤因子还需进一步查明。
高寒湿地是一种特殊的湿地类型, 具有蓄水调节、生物多样性保护和生态环境改善等重要功能[18]。作为最具代表性的内陆高寒湿地, 黄河首曲湿地素有黄河“蓄水池”、“高原水塔”之称, 其生态环境极其敏感与脆弱[19]。2013年, 甘肃黄河首曲湿地国家级自然保护区批准成立, 为黄河流域乃至高寒湿地的保护起到重要的推动与示范作用。然而, 近年来的气候变化和人为活动导致湿地植物群落组成和多样性发生变化, 湿地退化严重, 导致水源涵养能力降低[20]。同时, 有关高寒湿地相关研究尚未得出一致结论。如在青海省玛多县[21]、甘肃省尕海湿地[22]研究发现植物高度、盖度及地上生物量均随退化程度增加显著降低, 而另一项在玛多县的研究却发现地上生物量在中度退化程度下最高[23]。对于有机碳、全氮的报道多关于其组分、储量特征[24], 且土壤特性与植物群落关系研究鲜有报道。为此, 本文以黄河首曲湿地为研究对象, 立足于探讨以下几个问题:(1)不同退化程度对植物群落特征及多样性的影响;(2)不同退化程度对土壤理化特性的影响;(3)不同退化程度对植物群落特征与土壤理化性质之间的关系会产生怎样的影响?从而为揭示湿地植物群落退化机制提供科学基础, 为定向修复退化湿地提供理论依据。
1 材料与方法 1.1 研究区概况甘肃黄河首曲湿地国家级自然保护区地处甘肃省甘南藏族自治州玛曲县的东南部(101°54′12″—102°28′45″E, 33°20′01″—33°56′31″N), 保护区涉及玛曲县的曼日玛、采日玛、齐哈玛、欧拉4个镇和河曲马场、阿孜实验站(图 1);属高原大陆性气候, 一年中仅有冷暖季之别, 而无明显的四季之分。年均气温1.1℃, 7月平均气温10.7℃, 1月平均气温-9.7℃;年极端最高气温23.6℃, 年极端最低气温达—29.6℃。年平均降水量615.5mm, 全年降水约150d左右。全年没有绝对无霜期, 多年平均无霜期只有19d, 全年各月都有霜冻现象, 土壤持续冻结天数190d, 最大冻土层深度120cm[25]。研究区植物群落组成主要包括禾本科的垂穗披碱草(Elymus nutans);莎草科的华扁穗草(Blysmus sinocompressus);蔷薇科的鹅绒委陵菜(Potentilla anserina);菊科的长毛风毛菊(Saussurea hieracioides)、黄帚橐吾(Ligularia virgaurea);毛茛科的钝裂银莲花(Anemone obtusiloba)、长茎毛茛(Ranunculus nephelogenes var. longicaulis);蓼科的珠芽蓼(Polygonum viviparum)。土壤类型主要为高山草甸土, 主要分布在保护区欧拉乡西部, 海拔在4000m以上的高山陡坡, 西倾山也有少量分布。母质为残基坡积物, 土体冻融交替, 杂以冰渍沉积物, 土壤剖面厚度30—50m, 通层含砾石, 质地以轻壤、沙壤为主。

|
| 图 1 研究区位置及样方分布示意图 Fig. 1 Location of the study area and the investigated plots |
2021年7—8月对黄河首曲湿地自然保护区进行综合考察。考虑到自然保护区自然地理条件以及人为管理的影响, 为全面客观地反映自然保护区植被与土壤特征, 室内利用Google Earth沿保护区边界大致以5km为间距设置样地, 然后利用GPS在野外寻找样地, 同时根据交通可达性等适当调整样地位置, 使得样地尽可能均匀分布, 共计调查样方82个, 每个样方面积为0.5m×0.5m且均用GPS定位, 并记录样地距河道的距离、经纬度信息以及周围环境特征, 具体包括周围地形地势、放牧与啃食情况、鼠兔害情况以及采样时的天气状况。
现场鉴定样方中的植物物种, 并将个别现场无法识别的物种采集标本带回实验室进行鉴定, 记录个体数、物种数及高度, 样方总盖度及物种分盖度等指标, 地上生物量通过将样方内植株齐地刈割并装入信封袋带回实验室测定。湿地土壤样品取样时先清除覆盖在土壤上的植被地上部分, 用土钻采集0—10cm、10—20cm、20—40cm深的土样, 一部分装入铝盒用于土壤含水量的测定, 另一部分装入自封袋带回实验室进行土壤理化特性指标的测定。
将装袋的土壤样品带回实验室自然风干, 剔除较大的根系、砾石等, 研磨过筛至1mm和0.15mm粒径后进行相关理化性质测定。采用重铬酸钾氧化法、凯氏定氮法分别进行土壤有机碳(Soil organic carbon, SOC)、全氮(Total nitrogen, TN)的测定;采用pHS-3C型酸度计测定土壤pH值(水土比为2.5 ∶ 1);采用电导仪法测定土壤电导率(Electrical conductivity, EC)(水土比为5 ∶ 1)[26]。以上实验均在西北师范大学土壤地理实验室完成。
1.3 数据处理与分析为了更加准确地评价湿地退化, 参考Yang等[27]方法并选取指示湿地退化的5个因子。先将上述指标进行归一化处理, 然后进行因子分析, 求出各因子权重及得分系数, 进而根据各因子求出湿地退化指数(Wetland degradation index, WDI), 并通过K—均值聚类分析进行最优分割, 最终将所有调查样点分为无明显变化(ND)、轻度退化(LD)、中度退化(MD)和重度退化(HD)4个退化程度, 各退化程度样点数分别为10、30、28、14, 样地基本情况见表 1。
| 退化阶段 Degradation stages |
物种组成 Species composition |
基本情况 Basic condition |
| 无明显变化 Non Degradation(ND) |
早熟禾(Poa annua), 华扁穗草(Blysmus sinocompressus), 鹅绒委陵菜(Potentilla anserina) |
有季节性积水, 植物种类较丰富, 几乎无裸斑 |
| 轻度退化 Light Degradation(LD) |
垂穗披碱草(Elymus nutans), 鹅绒委陵菜(Potentilla anserina), 黄帚槖吾(Ligularia virgaurea), 钝裂银莲花(Anemone obtusiloba) | 有较少季节性积水, 植物种类丰富, 有少量干化斑块出现 |
| 中度退化 Moderate Degradation(MD) |
垂穗披碱草(Elymus nutans), 长花马先蒿(Pedicularis longiflora), 火绒草(Leontopodium hayachinense) | 无积水, 植物种类较少, 且有毒杂草出现 |
| 重度退化 Heavy Degradation(HD) |
由于严重退化, 几乎无植被 | 无积水, 植被稀疏, 属兔洞很多, 地表出现斑块性裸露, 部分土地沙化 |
数据归一化方法:
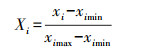
|
(1) |
式中, Xi为第i个指标归一化处理的结果;xi为第i个指标的原始值;ximin为第i个指标的最小值;ximax为第i个指标的最大值。
湿地退化指数(WDI):
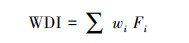
|
(2) |
式中, WDI为湿地退化评价指数;Fi表示因子;wi表示因子得分系数。
湿地退化常常是土壤和植物的退化, 随着退化过程的加剧毒杂草类逐渐取代优质牧草, 为探究退化过程中植物群落组成结构变化情况, 将植物种的功能型划分为禾本科、莎草科、豆科及毒杂草(主要包括乳浆大戟、黄帚橐吾、唐松草等), 各功能型植物的重要值计算如下[28]:
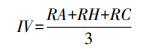
|
(3) |
式中, RA为相对密度, RH为相对高度, RC为相对盖度
群落物种多样性计算如下[28]:
Patrick物种丰富度指数(R):
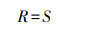
|
(4) |
Shannon-Wiener多样性指数(H):
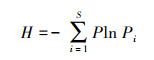
|
(5) |
Simpson多样性指数(D):
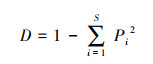
|
(6) |
Pielou均匀度指数(E):
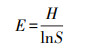
|
(7) |
式中, Pi为第i个物种在全体物种中出现的重要性比例, 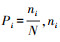
采用Origin 2017进行绘图, 采用SPSS 25.0进行单因素方差分析(One-way ANOVA), 采用最小显著性差异法(LSD)进行多重比较(α=0.05)。采用Canoco 5进行退化程度与植物因子间的主成分分析(Principal component analysis, PCA)及植物群落特征与土壤因子对应关系的冗余分析(Redundancy analysis, RDA)。
2 结果与分析 2.1 高寒湿地退化对植物群落结构的影响研究区各植物功能群重要值因退化程度和功能种类差异而表现不同(图 2)。禾本科重要值表现为ND>LD>MD>HD的变化趋势;莎草科重要值为ND>HD>MD>LD;豆科重要值的变化趋势为LD>ND>HD>MD;毒杂草重要值则呈MD>LD>HD>ND的变化, 且LD和MD程度下显著高于ND湿地(P < 0.05)。

|
| 图 2 不同退化程度下各功能型植物重要值 Fig. 2 Importance values of functional plants under different degrees of degradation 不同小写字母代表处理间存在显著差异(P < 0.05); ND: 无明显变化; LD: 轻度退化; MD: 中度退化; HD: 重度退化 |
随湿地退化, 植被高度、盖度、密度及地上生物量存在显著差异(P < 0.05)(图 3)。植被高度、地上生物量均随湿地退化呈降低趋势, LD和HD湿地植被高度显著低于ND;地上生物量在各退化程度间差异均显著, 植被盖度、密度均在HD湿地显著最低, 而在LD湿地达到高值。

|
| 图 3 不同退化程度高寒湿地植物群落特征 Fig. 3 Characteristics of alpine wetland vegetation community in different degrees of degradation 箱体代表上四分位数到下四分位数, 两端实线代表最值, 箱体内部实线代表中位数, 实心圆代表平均值 |
由图 4可知, 不同退化程度仅对Patrick物种丰富度指数产生显著影响(P < 0.05), 具体表现为在HD显著低于ND和LD湿地。湿地退化未对Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson多样性指数产生显著影响, 但Shannon-Wiener多样性指数、Pielou均匀度指数呈先增加后降低趋势, 而Simpson多样性指数随退化程度增加呈降低趋势。

|
| 图 4 不同程度退化高寒湿地植物多样性 Fig. 4 Alpine wetland plant diversity degraded to different degrees |
由图 5可知, 湿地退化仅对pH值产生显著影响(P < 0.05), 其中在0—10cm和10—20cm土层均表现为HD显著高于ND湿地;且20—40cm和0—40cm土层深度均表现为在HD下显著高于其他退化样地。各土层深度的土壤含水量与电导率因湿地退化并未发生显著变化(P>0.05), 但土壤含水量随湿地退化程度增加呈下降趋势。湿地退化使得土壤电导率在不同土层深度产生显著差异, 表现为在MD湿地, 0—10cm土层的电导率显著高于0—20cm和20—40cm(P < 0.05);土壤含水量随土层深度增加呈降低趋势, 而pH值随土层深度增加则呈增加趋势(图 6)。

|
| 图 5 不同程度退化高寒湿地土壤含水量、电导率和pH值 Fig. 5 Soil water content, electrical conductivity and pH of degraded alpine wetlands |

|
| 图 6 退化高寒湿地土壤理化性质随土层深度变化情况 Fig. 6 Variation of soil physical and chemical properties with soil depths in degraded alpine wetlands |
在0—10cm土层深度, 湿地退化对土壤有机碳、碳氮比产生显著影响(P < 0.05)(图 7)。土壤有机碳在ND下显著高于HD样地, 碳氮比在HD下显著高于ND和LD样地。在10—20cm土层深度, 土壤有机碳、全氮均表现为在ND下显著高于HD样地, 而碳氮比无显著差异。在20—40cm土层深度, 有机碳在ND下显著高于其他退化样地, 全氮在HD下显著高于ND和MD, 碳氮比无显著差异。在0—40cm土层深度有机碳和全氮均表现为ND下显著最高。此外, 土壤有机碳和全氮随土层深度增加而减少, 在ND、LD及MD样地表现为0—10cm土层深度显著高于其他土层深度(P < 0.05);而土壤碳氮比随土层深度增加未表现出显著差异(P>0.05)(图 6)。

|
| 图 7 不同程度退化高寒湿地土壤有机碳、全氮及碳氮比 Fig. 7 Soil organic carbon, total nitrogen and C/N ratio in different degraded alpine wetlands |
湿地退化与土壤理化性质的相关性表明, 退化程度与土壤有机碳、全氮及pH呈显著相关, 其中土壤有机碳、全氮与退化程度呈负相关关系, 而pH与退化程度呈正相关关系(表 2)。从图 8可以看出, 主成分1的贡献率为39.4%, 主成分2的贡献率为21.7%, 累积贡献率达到60.1%, 表明前两个主成分基本涵盖了大部分信息。Shannon-Wiener多样性在主成分1上有较高的载荷, 相关性较强, 这说明主成分1反映了Shannon-Wiener多样性的值可在一定程度上代表退化程度特征。Simpson多样性在主成分2上权重值较高, 这说明主成分2反映出Simpson多样性的值可表征湿地退化特征。
| 项目 Items |
土壤含水量 Soil water content |
电导率 Electric conductivity |
pH | 有机碳 Soil organic carbon |
全氮 Total nitrogen |
碳氮比 Carbon nitrogen ratio |
| 相关系数Correlation coefficient | -0.113 | 0.002 | 0.310** | -0.237* | -0.386** | 0.151 |
| P | 0.313 | 0.982 | 0.005 | 0.032 | 0.000 | 0.176 |
| *P < 0.05; * *P < 0.01 | ||||||

|
| 图 8 高寒湿地退化与植物因子的主成分分析 Fig. 8 Principal component analysis of alpine wetland degradation and plant factors Height:高度;Coverage:盖度;M:密度Density;AGB:地上生物量Aboveground biomass;H:Shannon-Winter多样性指数Shannon-Winter diversity index;E:Pielou均匀度指数Pielou evenness index;D:Simpson多样性指数Simpson diversity index;R:Patrick丰富度指数Patrick richness index |
本文通过对湿地不同退化程度下植物与土壤因子进行了RDA排序分析(表 3;图 9), 结果表明ND、LD、MD以及HD退化湿地轴1和轴2分别解释了植物与环境间的关系71.64%、9.3%, 80.62%、3.54%, 78.57%、10.38%以及83.1%、4.32%, 表明轴1和轴2包含了大部分信息, 因此利用前两轴的数据能够充分解释土壤因子与植物群落特征间的关系。ND湿地, RDA1与RDA2均主要反映了电导率的变化, 电导率与植物盖度、地上生物量以及多样性指数呈正向关系;LD湿地, RDA1主要反映了电导率的变化, 其与植物高度、盖度、地上生物量及均匀度指数呈正向关系, 而与Shannon-Winter多样性指数、Simpson多样性指数及Patrick丰富度指数呈负向关系, RDA2主要反映了土壤有机碳和全氮的变化, 其与植物盖度、地上生物量及多样性指数呈正向关系;MD湿地, RDA1主要反映了电导率、土壤含水量的变化, 电导率与植物盖度、地上生物量及多样性指数呈正向关系, 而土壤含水量与之相反。RDA2则主要反映了碳氮比的变化, 其与多样性指数呈正向关系;HD湿地, RDA1和RDA2均主要反映电导率的变化, 其与植物盖度、地上生物量呈正向关系, 而与多样性指数呈负向关系。
| 退化阶段 Degradation stages |
环境因子 Environmental factors |
解释量 Explains% |
伪F统计量 Pseudo-F |
P |
| 无明显变化 | SWC | 24.9 | 2.7 | 0.134 |
| Non degradation(ND) | EC | 24.9 | 3.5 | 0.04 |
| pH | 2.1 | 0.4 | 0.684 | |
| SOC | 8.4 | 1.5 | 0.302 | |
| TN | 7.9 | 1.6 | 0.322 | |
| C/N | 13.5 | 2.2 | 0.136 | |
| 轻度退化 | SWC | 6.7 | 2.3 | 0.082 |
| Light degradation(LD) | EC | 14.1 | 4.6 | 0.008 |
| pH | 1.8 | 0.7 | 0.546 | |
| SOC | 2.4 | 0.9 | 0.044 | |
| TN | 9.9 | 3.7 | 0.022 | |
| C/N | 1.7 | 0.6 | 0.594 | |
| 中度退化 | SWC | 21.2 | 9.1 | 0.002 |
| Moderate degradation(MD) | EC | 20.6 | 6.7 | 0.002 |
| pH | 2.1 | 1 | 0.436 | |
| SOC | 3.7 | 1.7 | 0.124 | |
| TN | 4.8 | 2.2 | 0.068 | |
| C/N | 1.6 | 0.7 | 0.02 | |
| 重度退化 | SWC | 2.8 | 0.5 | 0.788 |
| Heavy degradation(HD) | EC | 27.4 | 4.5 | 0.004 |
| pH | 9.7 | 1.7 | 0.156 | |
| SOC | 4 | 0.7 | 0.612 | |
| TN | 6.2 | 0.9 | 0.456 | |
| C/N | 1.4 | 0.2 | 0.946 | |
| SWC:土壤含水量Soil water content;EC:电导率Electric conductivity;SOC:土壤有机碳Soil organic carbon;TN:全氮Total nitrogen;C/N:土壤碳氮比Soil carbon nitrogen ratio;P < 0.05表示具有显著差异 | ||||

|
| 图 9 不同退化程度下植物群落特征与土壤因子RDA排序 Fig. 9 Plant community characteristics and soil factors RDA ranking under different degrees of degradation SWC:土壤含水量Soil water content;EC:电导率Electric conductivity;SOC:土壤有机碳Soil organic carbon;TN:全氮Total nitrogen;C/N:土壤碳氮比Soil carbon nitrogen ratio;RDA:冗余分析Redundancy Analysis;实线箭头表示植物因子, 虚线箭头表示土壤因子 |
湿地中物种组合和优势种地位的变化对湿地的群落特征和植物多样性有重要影响。本研究发现, 随退化程度加剧, 植物高度、盖度及地上生物量显著降低(图 2), 这与已有研究较为一致[29], 而佘延娣等[30]在三江源的研究中发现重度退化程度下地上生物量最高, 究其原因可能是退化过程中禾本科植物逐渐减少, 而毒杂草类有增加的趋势导致地上生物量部分增加。然而, 本研究中虽随退化程度增加毒杂草比例有所增加, 但多为乳浆大戟、唐松草等植株较小的物种, 地上植物部分减少。Patrick物种丰富度指数在重度退化下显著最低, 这可能是因为随退化程度增加丰富度指数呈单峰模式, 其峰值处于中度退化水平下[31]。与Wang等[32]不同的是, 其在黄河源头玛沁县研究发现在重度退化水平下具有高的物种数量, 这可能是因为虽同样存在放牧及啮齿类动物活动的影响, 但首曲湿地保护区内长期人为管理使得这种影响带来的采食偏食性及采食后排泄行为对种子传播的影响大大减弱。然而, 在退化过程中Shannon-Winter多样性指数和Pielou均匀度指数均不显著, 这可能是由于放牧活动在一定程度上抑制了植物生长, 从而利于劣势种的入侵, 导致物种多样性略有增加而在重度退化程度下明显降低[33]。综上所述, 湿地退化过程中带来的植物群落物种生长及结构的改变进而影响植物群落特征, 物种多样性的损失导致湿地功能退化对湿地生态系统带来负面影响。
3.2 高寒湿地退化对土壤理化性质的影响pH值的变化是高寒湿地土壤理化特性对退化的响应。本研究中, pH值随退化程度增加而显著增加, 随土层深度增加呈增加趋势, 且pH值在5.3—8.85, 说明退化湿地土壤呈现酸性—中性—碱性。与杨军等[34]在当雄县的研究较为一致, pH随退化程度增加呈增加趋势, 主要是因为退化导致的植被减少及强烈太阳辐射使土壤中有机酸破坏[35]。Osinuga等[36]发现不同土地利用类型湿地土壤有酸性至微碱性。土壤pH值的降低源于碱性阳离子的耗尽或是微生物氧化作用而产生有机酸[37]。
土壤有机碳、全氮是高寒湿地土壤养分对退化的响应。本研究发现, 土壤有机碳、全氮含量随退化程度加深显著降低, 且均随土层深度增加而降低, 这与已有研究较为一致[26]。这可能与植物地上部分凋落物的分解和植物根系大多分布在浅层有关。土壤有机碳含量受到有机物输入—输出关系的影响, 植被盖度与生产力以及呼吸作用的改变均会改变土壤有机碳含量[35]。一方面, 随着湿地退化程度加深, 植被盖度降低, 地表裸露面积持续增加, 加之风蚀与水流侵蚀的作用将带走大量表层积累的养分[9]。而另一方面地上生物量的减少将会间接的影响微生物数量和酶活性, 进而抑制养分供给能力, 导致养分供给能力下降[38]。然而, 因过度啃食作用的影响, 富集全氮的植物茎叶以及根系受到破坏导致养分流失[39]。杨元武等[40]研究发现退化过程中土壤温度的增加会促进养分分解, 而土壤水分的降低又会抑制分解作用, 但由于退化会造成湿地生态功能的破坏而导致其所固定的碳氮绝对含量的下降, 这也可能成为影响土壤有机碳、全氮含量降低的原因, 结果也会导致pH值的提高。综上所述, 土壤理化性质从不同角度响应高寒湿地的退化, 且二者之间相互作用的复杂关系共同影响着湿地功能。
3.3 植物群落与土壤属性之间的关系本研究结果表明, 湿地不同退化阶段影响植物群落的土壤因子略有不同, pH虽为响应湿地退化的关键土壤理化因子, 但电导率在湿地植物演替阶段扮演着重要角色。在ND和HD湿地, 电导率对植物群落的影响占据首要地位, 与植物多样性由正向关系转为负向关系。前人研究表明, 电导率与盐分含量呈正向关系, 电导率可在一定程度上表征盐分特征[41]。与ND相比, HD湿地土壤最为贫瘠, 此时电导率成为植物生长与生存的限制性因子。土壤盐分的增加将导致植物对养分物质资源的利用程度降低, 使本就有限的物种面临更大威胁, 最终改变物种多样性[42]。从LD至MD湿地, 土壤电导率与Shannon-Wiener多样性指数和Pielou均匀度指数由负向关系转为正向关系。Pielou均匀度指数可检验群落中所有物种的均匀性, 反映群落的物种多样性[43]。研究发现, 强大且具有竞争性的杂类草占据优势时, 其将在阳光、养分资源上进行掠夺, 导致种间竞争增大[44]。由LD至MD湿地土壤电导率逐渐增加, 且土壤有机碳、全氮含量相对较高。虽长花马先蒿、乳浆大戟等杂草类开始掠夺资源, 但因其植株相对矮小, 并未增加种间竞争, 这可能是导致均匀度并未降低的主要因素。加之Shannon-Wiener多样性指数最易受到人为活动干扰的影响[45]。此退化阶段牲畜所喜食的优质牧草比例下降, 导致其在采食时破坏压力减少, 为杂类草生长及生存提供空间, 使得植物物种多样性略微增加。综上所述, 土壤电导率虽不是响应湿地退化的关键因子, 但在影响湿地不同退化阶段植物群落中扮演重要角色, 充分说明研究湿地退化不可忽视土壤与植物之间的关系。
在LD湿地, 除电导率外, 有机碳和全氮成为主要影响因子, 且与植物盖度、地上生物量及多样性指数均呈正向关系(图 9)。与多数学者的研究较为一致[45—46], 即在不同类型生态系统均有发现植物盖度、生物量与有机碳、全氮之间的正向关系, 但有研究发现二者之间不存在或呈负向关系[47]。作为影响植物生产力的重要因素之一, 土壤养分供应的增加可见单峰关系[48], 当养分供应达到一定阈值或是群落结构的改变, 如毒杂草类的入侵可能会导致负向关系的出现。与Ma等[49]在甘南藏族高山湿地研究结果较为一致, Patrick物种丰富度指数与Shannon-Wiener多样性指数与全氮呈正相关, 但超过一定的阈值后就会下降, 而有机碳含量与物种多样性呈现负相关关系。一般地, 较高的有机碳含量对植物多样性具有积极影响[4]。究其原因可能是缺乏微生物生长所需要的有效氮源, 导致分解较为缓慢, 植物物种多样性较低[50]。在MD湿地, 植物多样性主要受到土壤含水量和碳氮比的影响。土壤含水量的降低将在一定程度上提高Shannon-Wiener多样性指数和Pielou均匀度指数。已有研究表明, 土壤水分的改变成为湿地植物群落演替的直接影响因素[51]。由于土壤水分含量的减少, 植物群落结构将发生改变, 更加耐旱以及牛羊所不喜食的物种将逐渐出现并被保留下来, 从而有利于多样性的提高。氮是植物生长发育的重要来源之一, 其通过直接获取和与微生物形成互惠共生体系间接获取, 而豆科植物共生固氮是主要途径[52]。然而, 随退化程度增加, 豆科植物重要值呈降低趋势, 而毒杂草类显著增加, 导致可供微生物利用的有效氮源较少不利于微生物多样性的提高。综上, 湿地退化阶段的不同, 导致植物群落与土壤因子之间的关系发生改变, 进一步充分说明湿地退化是土壤—植物的共同退化。
4 结论黄河首曲湿地不同退化程度下植物群落、土壤因子及其关系差异显著。随退化程度增加, 植被高度、盖度及地上生物量降低, 各多样性指数均随退化程度加深先增加后降低, 表明湿地退化使植物群落趋向复杂而后单一。因退化显著增加, 土壤逐渐呈碱性, 且土壤有机碳、全氮含量降低, 不利于养分积累。影响不同退化程度植物群落的土壤因子略有不同, 但土壤电导率在各湿地退化阶段扮演者重要角色。因此, 探究高寒湿地不同退化程度植物群落、土壤理化特征及其关系对于恢复管理湿地具有重要意义, 建议在高寒湿地恢复治理中注意提高有机质来源, 如进行草种补播以增加地上来源, 亦或通过施肥增加直接来源。退化程度加深时注意提高土壤有效氮源的补充。
| [1] |
Ghobadi Y, Pradhan B, Shafri H Z M, bin Ahmad N, Kabiri K. Spatio-temporal remotely sensed data for analysis of the shrinkage and shifting in the Al Hawizeh wetland. Environmental Monitoring and Assessment, 2015, 187(1): 4156. DOI:10.1007/s10661-014-4156-0 |
| [2] |
Junk W J, An S Q, Finlayson C M, Gopal B, Květ J, Mitchell S A, Mitsch W J, Robarts R D. Current state of knowledge regarding the world's wetlands and their future under global climate change: a synthesis. Aquatic Sciences, 2013, 75(1): 151-167. DOI:10.1007/s00027-012-0278-z |
| [3] |
Bano H, Lone F A, Bhat J I, Rather R A, Malik S, Bhat M A. Hokersar wet land of Kashmir: its utility and factors responsible for its degradation. Plant Archives, 2018, 18(2): 1905-1910. |
| [4] |
Gerdol R, Brancaleoni L, Lastrucci L, Nobili G, Pellizzari M, Ravaglioli M, Viciani D. Wetland plant diversity in a coastal nature reserve in Italy: relationships with salinization and eutrophication and implications for nature conservation. Estuaries and Coasts, 2018, 41(7): 2079-2091. DOI:10.1007/s12237-018-0396-5 |
| [5] |
Ladányi Z, Blanka V, Deák Á J, Rakonczai J, Mezösi G. Assessment of soil and vegetation changes due to hydrologically driven desalinization process in an alkaline wetland, Hungary. Ecological Complexity, 2016, 25: 1-10. DOI:10.1016/j.ecocom.2015.11.002 |
| [6] |
Dong S K, Sha W, Su X K, Zhang Y, Li S, Gao X X, Liu S L, Shi J B, Liu Q R, Hao Y. The impacts of geographic, soil and climatic factors on plant diversity, biomass and their relationships of the alpine dry ecosystems: Cases from the Aerjin Mountain Nature Reserve, China. Ecological Engineering, 2019, 127: 170-177. DOI:10.1016/j.ecoleng.2018.10.027 |
| [7] |
石明明, 周秉荣, 多杰卓么, 苏淑兰, 张帅旗, 马盼盼. 三江源区沼泽湿地退化过程中植被变化特征及评价指标体系. 西北植物学报, 2020, 40(10): 1751-1758. DOI:10.7606/j.issn.1000-4025.2020.10.1751 |
| [8] |
任国华, 邓斌, 后源. 黄河源区沼泽湿地退化过程中植物群落特征的变化. 草业科学, 2015, 32(8): 1222-1229. |
| [9] |
Jiménez-Ballesta R, García-Navarro F J, Martín-Consuegra S B, de los Reyes C P, Ortíz-Villajos J Á A, Miguel M F S. The impact of the storage of nutrients and other trace elements on the degradation of a wetland. International Journal of Environmental Research, 2018, 12(1): 87-100. DOI:10.1007/s41742-018-0072-4 |
| [10] |
Lin C Y, Li X L, Zhang J, Sun H F, Zhang J, Han H B, Wang Q H, Ma C B, Li C Y, Zhang Y X, Ma X Q. Effects of degradation succession of alpine wetland on soil organic carbon and total nitrogen in the Yellow River source zone, west China. Journal of Mountain Science, 2021, 18(3): 694-705. DOI:10.1007/s11629-020-6117-0 |
| [11] |
王英成, 芦光新, 赵丽蓉, 邓晔, 王军邦. 高寒草甸退化对土壤电导率变化影响的研究. 干旱区研究, 2021, 38(1): 104-113. DOI:10.13866/j.azr.2021.01.12 |
| [12] |
林春英, 李希来, 金惠瑛, 何生存, Jay Gao. 黄河源区河漫滩湿地退化过程土壤的变化特征. 中国农学通报, 2015, 31(33): 243-249. DOI:10.11924/j.issn.1000-6850.casb15080116 |
| [13] |
王婷, 张永超, 赵之重. 青藏高原退化高寒湿地植被群落结构和土壤养分变化特征. 草业学报, 2020, 29(4): 9-18. |
| [14] |
Trindade C R T, Landeiro V L, Schneck F. Macrophyte functional groups elucidate the relative role of environmental and spatial factors on species richness and assemblage structure. Hydrobiologia, 2018, 823(1): 217-230. DOI:10.1007/s10750-018-3709-6 |
| [15] |
张奇奇, 韩广轩, 路峰, 周英锋, 王晓杰, 李培广, 初小静, 贺文君, 于冬雪, 宋维民, 许景伟. 不同修复年限对黄河三角洲湿地植物多样性和群落稳定性的影响. 生态学杂志, 2022, 41(7): 1249-1257. DOI:10.13292/j.1000-4890.202207.034 |
| [16] |
Shen G, Yang X C, Jin Y X, Xu B, Zhou Q B. Remote sensing and evaluation of the wetland ecological degradation process of the Zoige Plateau Wetland in China. Ecological Indicators, 2019, 104: 48-58. DOI:10.1016/j.ecolind.2019.04.063 |
| [17] |
Kassa Y A, Anteneh B A, Teshome T T. Effect of Wetland Degradation and Conversion on Carbon Stock: The Case of Tekuma Wetland, Lake Tana Sub-Basin, Ethiopia. International Journal of Agriculture System, 2015, 3(2): 121-133. |
| [18] |
Nie Y, Li A N. Assessment of Alpine Wetland Dynamics from 1976-2006 in the Vicinity of Mount Everest. Wetlands, 2011, 31(5): 875-884. DOI:10.1007/s13157-011-0202-7 |
| [19] |
Bai J H, Lu Q Q, Wang J J, Zhao Q Q, Ouyang H, Deng W, Li A N. Landscape pattern evolution processes of alpine wetlands and their driving factors in the Zoige Plateau of China. Journal of Mountain Science, 2013, 10(1): 54-67. DOI:10.1007/s11629-013-2572-1 |
| [20] |
薛鹏飞, 李文龙, 朱高峰, 周华坤, 刘陈立, 晏和飘. 黄河首曲玛曲县高寒湿地景观格局演变. 植物生态学报, 2021, 45(5): 467-475. |
| [21] |
Li X L, Xue Z P, Gao J. Environmental Influence on Vegetation Properties of Frigid Wetlands on the Qinghai-Tibet Plateau, Western China. Wetlands, 2016, 36(5): 807-819. DOI:10.1007/s13157-016-0788-x |
| [22] |
马维伟, 李广, 石万里, 王辉, 罗永忠, 张亮, 聂志刚. 甘肃尕海湿地退化过程中植物生物量及物种多样性变化动态. 草地学报, 2016, 24(5): 960-966. |
| [23] |
刘凯, 李希来, 金立群, 孙华方, 李清德. 黄河源湖泊湿地退化过程土壤和植被的变化特征. 生态科学, 2017, 36(3): 23-30. |
| [24] |
林春英, 李希来, 李红梅, 孙海松, 韩辉邦, 王启花, 金立群, 孙华方. 不同退化高寒沼泽湿地土壤碳氮和贮量分布特征. 草地学报, 2019, 27(4): 805-816. |
| [25] |
王亚茹, 赵雪雁, 张钦, 雒丽, 薛冰. 高寒生态脆弱区农户的气候变化适应策略评价——以甘南高原为例. 生态学报, 2017, 37(7): 2392-2402. |
| [26] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000. |
| [27] |
Yang T X, Chen J D, Zhong X Y, Yang X C, Wang G, Yao Y, Sternberg M, Sun W. Divergent responses of plant biomass and its allocation to the altered precipitation regimes among different degraded grasslands in China. Plant and Soil, 2022, 473(1): 149-166. |
| [28] |
Zhang Y R, Wang R Q, Kaplan D, Liu J. Which components of plant diversity are most correlated with ecosystem properties? A case study in a restored wetland in northern China. Ecological Indicators, 2015, 49: 228-236. DOI:10.1016/j.ecolind.2014.10.001 |
| [29] |
Zhang D J, Qi Q, Tong S Z, Wang X H, An Y, Zhang M Y, Lu X G. Soil Degradation Effects on Plant Diversity and Nutrient inTussock meadow Wetlands. Journal of Soil Science and Plant Nutrition, 2019, 19(3): 535-544. |
| [30] |
佘延娣, 杨晓渊, 马丽, 张中华, 王党军, 黄小涛, 马真, 姚步青, 周华坤. 退化高寒草甸植物群落和土壤特征及其相互关系研究. 草地学报, 2021, 29(S1): 62-71. |
| [31] |
郝爱华, 薛娴, 彭飞, 尤全刚, 廖杰, 段翰晨, 黄翠华, 董斯扬. 青藏高原典型草地植被退化与土壤退化研究. 生态学报, 2020, 40(3): 964-975. |
| [32] |
Wang X X, Dong S K, Sherman R, Liu Q R, Liu S L, Li Y Y, Wu Y. A comparison of biodiversity-ecosystem function relationships in alpine grasslands across a degradation gradient on the Qinghai-Tibetan Plateau. The Rangeland Journal, 2015, 37(1): 45-55. |
| [33] |
袁建立, 江小蕾, 黄文冰, 王刚. 放牧季节及放牧强度对高寒草地植物多样性的影响. 草业学报, 2004, 13(3): 16-21. |
| [34] |
杨军, 刘秋蓉, 王向涛. 青藏高原高山嵩草高寒草甸不同退化阶段植物群落与土壤养分. 应用生态学报, 2020, 31(12): 4067-4072. |
| [35] |
Rebelo A J, Emsens W J, Meire P, Esler K J. The impact of anthropogenically induced degradation on the vegetation and biochemistry of South African palmiet wetlands. Wetlands Ecology and Management, 2018, 26(6): 1157-1171. |
| [36] |
Osinuga O A, Oyegoke C O. Degradation assessment of wetlands under different uses: implications on soil quality and productivity. African Journal of Agricultural Research, 2019, 14(1): 10-17. |
| [37] |
Norbert N F, Tening Aaron S, George B C, Kenneth M, Godswill A A, Vivian B C. Selected physicochemical properties and quality of soils around some rivers of Cameroon. Journal of Soil Science and Environmental Management, 2018, 9(5): 68-80. |
| [38] |
刘纪远, 徐新良, 邵全琴. 近30年来青海三江源地区草地退化的时空特征. 地理学报, 2008, 63(4): 364-376. |
| [39] |
白军红, 邓伟, 朱颜明, 栾兆擎, 张玉霞. 霍林河流域湿地土壤碳氮空间分布特征及生态效应. 应用生态学报, 2003, 14(9): 1494-1498. |
| [40] |
杨元武, 李希来, 周旭辉, 祁银姐, 师月英, 李成一, 周华坤. 高寒草甸植物群落退化与土壤环境特征的关系研究. 草地学报, 2016, 24(6): 1211-1217. |
| [41] |
Jiang Y, Hao W, Zhang Y G, Liang W J. Geostatistical analyses of soil electrical conductivity in a vegetable greenhouse field with different data sets. Journal of Environmental Research, 2008, 2(3): 125-130. |
| [42] |
Zhao C Z, Zhang H, Song C P, Zhu J K, Shabala S. Mechanisms of plant responses and adaptation to soil salinity. The Innovation, 2020, 1(1): 69-109. |
| [43] |
Joyce C B. Ecological consequences and restoration potential of abandoned wet grasslands. Ecological Engineering, 2014, 66: 91-102. |
| [44] |
Kołos A, Banaszuk P. Mowing as a tool for wet meadows restoration: effect of long-term management on species richness and composition of sedge-dominated wetland. Ecological Engineering, 2013, 55: 23-28. |
| [45] |
Hu Z M, Li S G, Guo Q, Niu S L, He N P, Li L H, Yu G R. A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China. Global Change Biology, 2016, 22(4): 1385-1393. |
| [46] |
Yuan Z Q, Jiang X J. Vegetation and soil covariation, not grazing exclusion, control soil organic carbon and nitrogen in density fractions of alpine meadows in a Tibetan permafrost region. Catena, 2021, 196: 104832. |
| [47] |
Bi X, Li B, Fu Q, Fan Y, Ma L X, Yang Z H, Nan B, Dai X H, Zhang X S. Effects of grazing exclusion on the grassland ecosystems of mountain meadows and temperate typical steppe in a mountain-basin system in Central Asia's arid regions, China. Science of the Total Environment, 2018, 630: 254-263. |
| [48] |
Tschirhart J. Resource competition among plants: from maximizing individuals to community structure. Ecological Modelling, 2002, 148(2): 191-212. |
| [49] |
Ma M Y, Zhu Y J, Wei Y Y, Zhao N N. Soil Nutrient and Vegetation Diversity Patterns of Alpine Wetlands on the Qinghai-Tibetan Plateau. Sustainability, 2021, 13(11): 6221. |
| [50] |
Rezgui C, Trinsoutrot-Gattin I, Benoit M, Laval K, Riah-Anglet W. Linking changes in the soil microbial community to c and n dynamics during crop residue decomposition. Journal of Integrative Agriculture, 2021, 20(11): 3039-3059. |
| [51] |
Wang C, Fu B J, Zhang L, Xu Z H. Soil moisture-plant interactions: an ecohydrological review. Journal of Soils and Sediments, 2019, 19(1): 1-9. |
| [52] |
Genre A, Lanfranco L, Perotto S, Bonfante P. Unique and common traits in mycorrhizal symbioses. Nature Reviews Microbiology, 2020, 18(11): 649-660. |